Глава III. Морфология клетки. Цитоплазма. Клеточное ядро
Благодаря разработке новых методов цитологического исследования (люминесцентной микроскопии, цитофотометрии, авторадиографии и др.), а особенно в результате использования для изучения клетки электронного микроскопа, знания о ней резко обогатились. Электронный микроскоп не только дал возможность уточнить тончайшее строение клеток и их ранее известных органелл (органоидов), но и выявил новые, до тех пор не известные структурные элементы.
Важнейшим строительным материалом клетки, в том числе ее органелл, оказались тончайшие перепонки - мембраны. На долю всех мембран в клетке приходится свыше 40% ее общей массы. Даниелли и Давсон предложили единую схему молекулярного строения элементарной мембраны (рис. 12В). Такое представление об общем принципе строения элементарных мембран как образованных одним бимолекулярным слоем липидов в середине и двумя мономолекулярными белковыми слоями с обеих поверхностей, подтвержденное Робертсоном, господствует в цитологии до последнего времени. Толщина элементарной мембраны составляет 60 - 75 Å, а каждого из ее слоев - 20 - 25 Å. Мембраны пронизаны порами, отграниченными белковыми молекулами.
Несмотря на большое сходство трехслойного электронномикроскопического строения, различные липопротеиновые мембраны клетки отличаются друг от друга по конкретному составу белковых и липидных молекул. Содержание последних сильно варьирует - от 20 до 75%. Соответственно в обратном отношении колеблется количество белков. Белки в разных мембранах различны, их макромолекулы могут быть индивидуальными или же организованными в комплексы. Все это обусловливает неодинаковую проницаемость элементарных мембран и придает им различные функциональные свойства. При общем сходстве в строении (в принципе отвечающем схеме Даниелли - Давсона - Робертсона) разные элементарные мембраны прежде всего варьируют по набору ферментов, определяющих характер метаболических процессов, в том числе синтетических, разыгрывающихся на их поверхностях. Белки - ферменты располагаются в мембранах в соответствии с порядком их включения в цепи реакций.
Было установлено, что клетка целиком одета тончайшей субмикроскопической оболочкой (рис. 13), имеющей типичную структуру трехслойной элементарной мембраны. Толщина ее обычно варьирует в указанных пределах - 60 - 75 Å. Наружную клеточную, цитоплазматическую мембрану называют еще плазматической оболочкой, цитолеммой, а чаще - плазмолеммой. Через плазмолемму непрерывно происходит обмен веществ между клеткой и ее окружением. Он осуществляется как через поры, так и непосредственно через мембрану и определяется ее способностью к проницаемости.
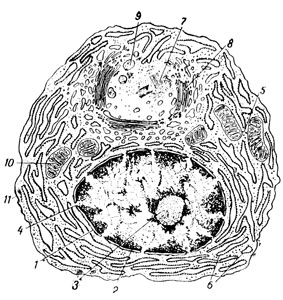
Рис. 13. Схема электронно-микроскопического строения плазматической клетки (из И. Ф. Иванова): 1 - ядерная оболочка; 2 - хроматин; 3 - ядрышко; 4 - пора в ядерной оболочке; 5 - свободные рибосомы; 6 - гранулярный эндоплазматический ретикулум; 7 - центриола; 8 - комплекс Гольджи; 9 - секреторная гранула; 10 - митохондрия; 11 - плазмолемма
Вопрос о наличии мембраны вокруг клетки долгое время был спорным, причем большая часть ученых склонялась к тому, что подобная, невидимая в обычный микроскоп перепонка отсутствует. Явления проницаемости внутрь клетки одних веществ и непроницаемости других они объясняли не характером тончайшей структуры нежного поверхностного слоя клетки, а исключительно состоянием ее живого вещества в целом. Электронный микроскоп разрешил этот давний спор в пользу сторонников мембранной теории, хотя и не умалил значения работ, показавших важнейшую роль состояния протоплазмы в процессе поступления тех или иных веществ в клетку.
Было обнаружено, что клеточная мембрана во многих случаях образует разного рода лабильные и стабильные выпячивания и впячивания. Постоянные многочисленные пальцевидные выросты мембраны на свободной поверхности некоторых клеток получили наименование микроворсинок. Они являются приспособлением к увеличению поверхности соприкосновения клетки с наружной средой. Так, например, клетки тонкого кишечника обращены микроворсинками в просвет кишки (рис. 10Б и 11В) и через них происходит всасывание переваренных продуктов пищи. Благодаря наличию микроворсинок всасывающая поверхность клеток оказывается увеличенной в тысячи раз.
Аналогичное функциональное значение имеют и впячивания плазмолеммы внутрь клетки. Подобные складки встречаются у базальной поверхности клеток (например, в мочевых канальцах почки) и служат той же цели - увеличению площади контакта клетки, но уже с внутренней средой организма. Наконец, часто встречающиеся выпячивания и впячивания мембраны на боковых сторонах клеток, объединенных в пласт, играют роль скреплений между клетками. Они имеют характер "замков" - выпячивание мембраны одной клетки входит в соответствующее впячивание соседней (рис. 14), причем благодаря конечному расширению такого выпячивания происходит его фиксация в теле рядом лежащей клетки. Наличие многочисленных "замков" подобного рода, расположенных на прилежащих друг к другу стенках соседних клеток, обеспечивает их прочное скрепление.
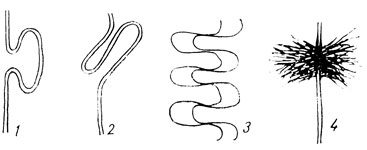
Рис. 14. Способы взаимного скрепления клеток. Схема: 1 и 2 - 'замков', 3 - интердигитаций, 4 - десмосомы
Встречается еще один вид взаимоскрепления клеток, обозначаемый как интердигитация. Расположенные рядом клетки образуют навстречу друг другу пальцевидные выросты плазмолеммы, входящие в промежутки между противоположными (рис. 14-3).
Прочные скрепления между клетками образуются при помощи десмосом (соединяющих телец). Эти расположенные друг против друга уплотненные участки плазмолеммы имеют овальную форму диаметром около 0,5 мк (рис. 14-4).
До сих пор еще не выяснено, имеется ли цементирующее вещество между соседними клетками. Большая часть исследователей считает, что между клетками находится щелевидное пространство, по которому циркулирует тканевая жидкость, тканевая лимфа.
Благодаря электронному микроскопу в клетке между ее органоидами в гиалоплазме (матриксе цитоплазмы) были открыты новые структуры, состоящие из мембран, а точнее целая система канальцев с местными расширениями (цистернами), пронизывающая все клеточное тело наподобие трехмерной сети. На срезе эти канальцы обычно поперечно или косо перерезаны (рис. 10Б-3 и рис. 13-6). За сходство с сетью данную циркуляторную систему клетки назвали цитоплазматическою сетью, или эндоплазматическим ретикулумом. В отдельных местах стенки канальцев эндоплазматического ретикулума непосредственно переедят в плазмолемму, и таким образом циркуляторная система оказывается связанной со средой, окружающей клетку, откуда в нее непосредственно могут поступать различные вещества.
Некоторые ученые сравнивают эндоплазматический ретикулум с сосудистой системой нашего тела. Согласно их мнению, по эндоплазматическому ретикулуму непрерывно циркулирует жидкость с различными растворенными в ней веществами, участвующими в метаболизме клетки, подобно тому как по кровеносным сосудам циркулирует кровь, несущая каждой клетке нашего тела необходимые для ее жизнедеятельности вещества.
Мембраны, образующие стенки эндоплазматического ретикулума, имеют толщину 40 Å, хотя могут достигать и 75 Å. Их толщина, как и общая конструкция эндоплазматического ретикулума, в разных клетках различна. Просвет канальцев варьирует в очень больших пределах от 700 до 5000 Å.
Одни из мембран эндоплазматического ретикулума гладкие (рис. 10Б), другие - шероховатые (рис. 13). Последние имеют на своей наружной поверхности гранулы диаметром 150 - 200 Å. Такого же размера и характера гранулы лежат свободно в матриксе цитоплазмы. Они называются рибосомами.
Рибосомы, или гранулы Палада, построены из двух субъединиц, отличающихся размерами. Они состоят наполовину из белка и наполовину из рибонуклеиновой кислоты, за что и получили наименование рибосом. Каждая из субъединиц рибосом представляет собой одну нитевидную молекулу рибонуклеопротеида (РНП), свернувшуюся в шарик (глобулу). Из всех разновидностей РНК клетки рибосомальная обладает наибольшим молекулярным весом и на ее долю приходится почти 9/10 всей клеточной РНК.
Густой гранулярный эндоплазматический ретикулум, придающий цитоплазме в месте его расположения базофильный характер (из-за обилия рибонуклеиновой кислоты), называют эргастоплазмой (рабочей плазмой), так как здесь происходит интенсивный синтез белка. Такой ретикулум свойствен клеткам, усиленно вырабатывающим, секретирующим ферменты - вещества белковой природы, например, клеточным элементам поджелудочной железы. Канальцы эргастоплазмы располагаются в них концентрически вокруг ядра. Такой же характер присущ гранулярному эндоплазматическому ретикулуму и в плазматических клетках (рис. 13) соединительной ткани, вырабатывающих антитела - белковые вещества, обеспечивающие организму иммунитет.
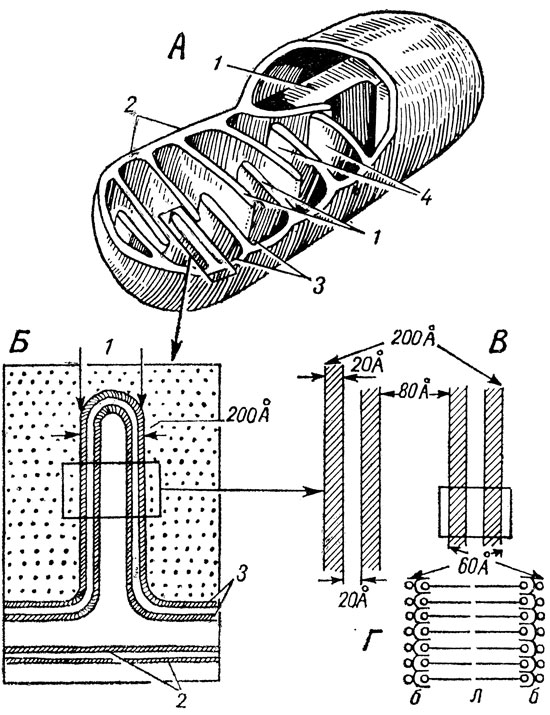
Рис. 15. Схема (де Робертиса) электронномикроскопического строения митохондрии. А - объемная схема расположения наружной (2) и внутренней (1) мембран, крист (3) и матрикса (4). Б - митохондриальная криста при большем увеличении. В - часть кристы при еще большем увеличении. Г - молекулярная структура одной элементарной мембраны
В гладком ретикулуме также протекают синтетические процессы, но они связаны с продукцией не белка, а углеводов и липидов.
Митохондрии обнаруживают отчетливое мембранное строение - их стенка сформирована из двух мембран. Наружная мембрана гладкая, а внутренняя образует многочисленные складки или гребешки, обозначаемые еще кристами (рис. 13 и 15 А). Кристы делят полость митохондрии на отсеки. Наружная и внутренняя мембраны митохондрии являются элементарными мембранами. Каждый из трех слоев этих элементарных мембран имеет толщину 20 Å (рис. 15В). Промежуток между наружной и внутренней мембранами составляет 80 Å, так что общая толщина стенки митохондрии равняется 200 Å. В матриксе митохондрии обнаруживаются гранулы размером около 500 Å. Такое своеобразное строение митохондриев способствует выполнению ими сложной биохимической работы - окислению поступающих продуктов и синтезу молекул АТФ. Эти химические процессы протекают поэтапно и весь цикл осуществляется путем последовательного включения целой цепи ферментов. В каждой из камер митохондриев, отделенной кристами, происходят характерные для нее реакции. Это как бы отдельные "цеха фабрики" и процесс завершается, пройдя поэтапно через все отсеки. Течению реакций способствует увеличенное кристами количество плоскостей, ибо, как известно из физической химии, различного рода реакции протекают на поверхностях гораздо быстрее.
В настоящее время установлено наличие скрученной нити ДНК, лежащей в центральной светлой зоне матрикса вдоль митохондриев. Митохондрии в процессе активной деятельности снашиваются, срок их существования измеряется несколькими неделями. Новые митохондрии возникают путем отпочкования от материнских, что связывают с наличием в этих органоидах собственной ДНК. Однако имеются описания развития митохондрий из выпячиваний плазмолеммы или канальцев эндоплазматического ретикулума, отрывающихся от них и постепенно приобретающих структуру данных органоидов.
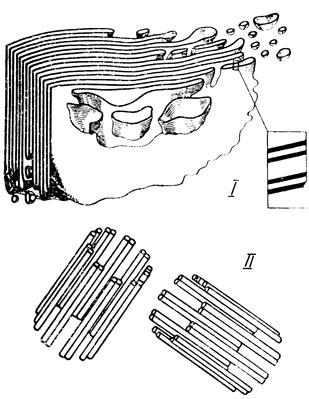
Рис. 16. Схемы пространственной организации пластинчатого комплекса (I - по И. Б. Токину) и центриолей (II - по Карассо и Фавру)
Аппарат Гольджи, обозначаемый теперь как пластинчатый комплекс, под электронным микроскопом представляет совокупность следующих элементов: 1) 5 - 8 уплощенных мешочков-цистерн, расположенных параллельно друг другу, наподобие штабеля (рис. 16-1). Стенки цистерн - элементарные мембраны толщиной 60 - 75 Å; 2) мелкие пузырьки, локализованные у боков штабеля и отрывающиеся от краев мешочков; 3) крупные пузырьки, лежащие по периферии. Стенка пузырьков также представляет собой трехслойную элементарную мембрану. Данные новейших исследований подтвердили, что аппарат Гольджи может иметь и сетчатую структуру. Особенно четко такая структура аппарата Гольджи была обнаружена в некоторых растительных клетках. Уложенные в виде штабеля уплощенные цистерны занимают центральную часть пластинчатого комплекса. Они дают начало многочисленным трубочкам диаметром 200 - 400 Å, которые, анастомозируя друг с другом, образуют сеть, оплетающую уплощенные цистерны.
Электронный микроскоп подтвердил теорию Д. Н. Насонова об основном функциональном значении аппарата Гольджи как органоида клетки, в котором происходит аккумуляция синтезируемых и резорбируемых продуктов. По современным представлениям, синтезированный на рибосомах гранулярного ретикулума белок поступает в итоге в пластинчатый комплекс, где постепенно накапливается и оформляется в виде плотных гранул секрета, по-видимому, в результате потери им воды осмотическим путем. Исходя из этого, пластинчатый комплекс называют "упаковочным цехом" клеточной фабрики, вырабатывающей продукт по описанному "конвейеру".
Благодаря электронномикроскопическим исследованиям выявлена определенная закономерность в структуре клеточного центра; цилиндрические центриолы, имеющие диаметр около 1500 Å, во всех случаях состоят из 9 одинарных, чаще двойных (рис. 16-II), а иногда тройных трубочек. Стенки трубочек имеют толщину 50 - 60А, диаметр просвета между ними 50 Å. Центриолы диплосомы располагаются взаимно перпендикулярно (рис. 13 и 16-II). Это объясняется тем обстоятельством, что основной способ образования центриол - отпочковывание от материнской - происходит именно в таком положении. Было установлено, что почкование центриол начинается на стадии анафазы митоза. Новая центриола строится у одного из концов "старой", располагаясь перпендикулярно к последней. В конце профазы следующего митоза она достигает размеров материнской. В настоящее время имеются основания считать, что к тому моменту, когда дочерняя центриола полностью созревает и отделяется от материнской, в них обеих уже начинается процесс образования новых центриол. Таким образом, в телофазе уже в каждой из дочерних клеток имеется по 4 центриолы - 2 полностью сформированных и 2 еще не созревших.
Центросфера переходит непрерывно в окружающую цитоплазму, в ней не встречаются ни митохондрии, ни рибосомы.
Как уже отмечалось в предыдущей главе, клеточный центр, представленный расходящимися к полюсам делящейся клетки центриолами, является той осью, вдоль которой строится митотический аппарат. На основании трубчатого строения нитей митотического аппарата и сходства их диаметра с диаметром трубочек центриол была высказана мысль о непосредственном образовании нитей веретена центриолярными трубочками. Было установлено, что митотический аппарат на 9/10 состоит из белков (а это составляет приблизительно 1/10 всего клеточного белка), причем к моменту его построения все необходимые белковые молекулы уже имеются в цитоплазме. Их сборка в нити веретена осуществляется центриолами.
Такие метаплазматические структуры, как мио-, невро- и тонофибриллы под электронным микроскопом оказались пучками соответствующих тончайших протофибриллей, или филаментов, диаметр которых составляет 40 - 100 Å.
Сравнительно недавно в клетках разных тканей был обнаружен еще один структурный элемент клетки - микротрубочки. По диаметру они близки нитям митотического аппарата (порядка 200 - 250 Å). Микротрубочки являются в основном опорными структурами клетки, о чем можно судить по характеру их расположения. Они залегают как вблизи плазмолеммы, так и около разных клеточных органоидов. В вытянутых клетках микротрубочки лежат по их длине, а в шарообразных - радиально.
Реснички, встречающиеся на свободной поверхности клетки, одеты плазмолеммой, а в их матриксе залегают 2 центральные непарные фибриллы-трубочки и 9 пар периферических. В жгутиках сперматозоидов имеется еще 9 непарных мелких добавочных фибрилл, связанных тончайшими нитями с основными.
Базальные тельца (кинетосомы), лежащие в основании ресничек, по своему строению сходны с центриолами. Они также обладают цилиндрической формой, имеют длину около 0,5 мк и ширину около 1500 Å. Сходство это обусловлено тем, что базальные тельца являются продуктами многократного деления центриол. Реснички вырастают из базальных телец, а от трубочек последних начинаются периферические фибриллы ресничек. Центральные фибриллы не связаны с базальным тельцем. В их основании располагается аксосома - аксиальная (осевая) гранула, лежащая над базальным тельцем. С центральными фибриллами связывают опорную функцию, с периферическими - двигательную. Основанием этому служит обнаружение в периферических фибриллах сократимого белка, аналогичного мышечному.
Корневые нити реснитчатого конуса, основанием которому служат базальные тельца (рис. 10А), под электронным микроскопом имеют поперечную исчерченность, что присуще структурам белковой природы. Базальные тельца, как показал химический анализ, состоят наполовину из белка. В них обнаружены РНК (2%) и ДНК (3%), чем объясняется их способность к самовоспроизведению.
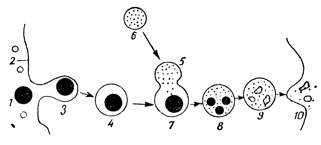
Рис. 17. Схема функционирования лизосом (по Де-Дюву, изменено): 1 - 3 - 4 - образование фагосомы; 2 - плазмолемма; 6 - лизосома; 5 и 7 - слияние лизосомы с фагосомой, в результате чего образуется пищеварительная вакуоль (8); 9 - непереваренные остатки в пищеварительной вакуоли, выбрасываемые из клетки (10)
В 1955 году Де-Дюв открыл еще один компонент клеток - лизосомы. В их названии отражена специфическая функция данных телец - растворение (по-гречески "лизис" - переваривание). Вещества, поступающие в клетку, перевариваются с помощью лизосом. Лизосомы чаще всего имеют эллипсоидную форму и сильно варьируют по своим размерам, достигая в среднем 0,5 мк в диаметре. Одеты они трехслойной элементарной мембраной, что определяется их происхождением. По-видимому, лизосомы формируются, подобно гранулам секрета, в комплексе Гольджи. В образовании их содержимого участвует и эндоплазматический ретикулум. Лизосомы представляют собой как бы замкнутые округлые мешочки, заполненные переваривающими гидролитическими ферментами (рис. 17), с помощью которых могут быть расщеплены разные органические соединения (белки, нуклеиновые кислоты, углеводы), фагоцитированные клеткой.
Как показали электронномикроскопические исследования, захват клеткой фагоцитируемых частичек обычно происходит путем впячивания под ними плазмолеммы и последующего смыкания ее краев над частичками, погрузившимися в цитоплазму. После отрыва от плазмолеммы ее участка, одевающего фагоцитированную частичку, возникает фагосома, свободно лежащая в цитоплазме. Фагосомы могут возникать также в результате образования клеткой псевдоподий (ложноножек) и постепенного активного захвата ими фагоцитируемой частички. В итоге образуется такая же фагосома, окруженная мембраной (участком плазмолеммы) и погруженная в цитоплазму.
Лизосомы, сливаясь с фагосомами, образуют пищеварительные вакуоли, в которых фагоцитированные частички подвергаются действию протеолитических ферментов, внесенных в пищеварительную вакуоль лизосомой.
Аналогично захвату плотных частичек путем инвагинации под ними плазмолеммы может происходить захват клетками капелек жидкости - явление, которое получило название пиноцитоза (по-гречески "пино" означает питье). Пиноцитированные пузырьки могут, подобно фагосомам, сливаться с лизосомами в пищеварительные вакуоли.
Что же касается гиалоплазмы, матрикса цитоплазмы, в которой залегают все органоиды клетки и включения, то и под электронным микроскопом она оказывается в общем бесструктурной. Уровень ее организации уже не субмикроскопический, а молекулярный - она представлена главным образом глобулярными белковыми молекулами.
Изучение интерфазного ядра под электронным микроскопом показывает, что оболочка его состоит из двух элементарных мембран, разделенных перинуклеарным пространством, и пронизана многочисленными порами, через которые облегчается двусторонний обмен между кариоплазмой и цитоплазмой (рис. 18А). Мембраны, образующие ядерную оболочку, толще, чем в эндоплазматическом ретикулуме, хотя наружная из них местами непосредственно связана с последним. Толщина мембран достигает 100 Å, а пространство между ними колеблется от 100 до 300 Å. В результате общая толщина ядерной оболочки в различных клетках составляет 300 - 400 Å. Поры ядерной оболочки имеют диаметр около 500 Å и лежат на поверхности ядра более или менее густо (примерно 50 на 1 мк2 ядерной оболочки). В тех местах, где наружная мембрана ядерной оболочки переходит непосредственно в канальцы эндоплазматического ретикулума, могут образовываться местные расширения перинуклеарного пространства. Полость этих цистерн достигает в диаметре 0,1 мк.
Поверхности контакта между ядром и цитоплазмой могут увеличиваться благодаря впячиваниям или выпячиваниям ядерной оболочки (рис. 18Б). С ядерной оболочкой, точнее с наружным ее листком, связывают новообразование канальцев и цистерн эндоплазматического ретикулума. В то же время возникновение заново ядерной оболочки, резорбирующейся в поздней профазе митоза и восстанавливающейся в телофазе, происходит, как считают, за счет везикулярного материала, берущего начало от эндоплазматического ретикулума.
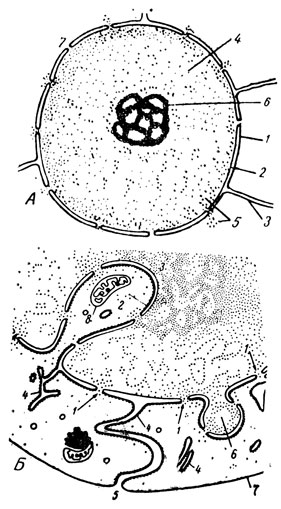
Рис. 18. А - схема субмикроскопического строения ядра клетки (из А. Н. Студитского): 1 - наружная мембрана ядерной оболочки; 2 - внутренняя мембрана ядерной оболочки; 3 - каналец, связывающий оболочку ядра с эндоплазматическим ретикулумом; 4 - кариоплазма; 5 - выход рибосом из ядра в цитоплазму через поры в ядерной оболочке (7); 6 - ядрышко. Б - схема (по В. Я. Бродскому), показывающая взаимоотношения между ядром клетки и цитоплазмой; 1 - поры ядерной оболочки; 2 - впячивание ядерной оболочки до соприкосновения ее с ядрышком (3), 4 - участки гранулярного ретикулума; 5 - место перехода канальца эндоплазматического ретикулума в плазмолемму; 6 - выпячивание ядерной оболочки; 7 - плазмолемма
В кариоплазме неделящейся клетки под электронным микроскопом между ядерной оболочкой и ядрышком видна бесструктурная жидкость, представляющая собой раствор белков. Ее называют клеточным соком, или кариолимфой. В кариолимфе заметны лишь гранулы - рибосомы и мелкая зернистость, которую рассматривают как выступающие на тончайшем срезе поперечные разрезы хроматиновых нитей - хромонем, деспирализующихся в период интерфазы. Однако иногда на срезах выявляются через ядра небольшие отрезки нитей, а в некоторых случаях и образуемые этими нитями хромосомы (рис. 19). Эти данные свидетельствуют о том, что хромосомы в интерфазном ядре могут не распадаться на отдельные нити или молекулы нуклеогистона и сохраняются как целые структуры.
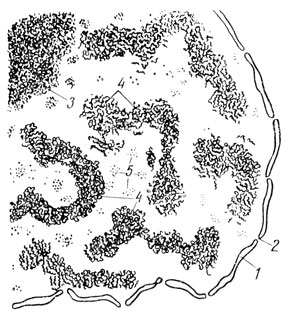
Рис. 19. Схема электронно-микроскопического строения ядра (из Л. Н. Жинкина и Ю. Б. Бахтина): 1 - ядерная оболочка; 2 - пора; 3 - ядрышко; 4 - хромосомы; 5 - рибосомы (возникают в ядрышках)
Ядрышко является постоянной структурой интерфазного ядра (рис. 18А). Оно состоит из двух компонентов гранулярных и фибриллярных образований, содержащих РНК. Гранулы соответствуют (по размерам (диаметр 150 Å) и химическому составу рибосомам цитоплазмы. Нитчатые элементарные структуры ядрышка имеют толщину около 100 Å. В совокупности они образуют нуклеолонему - нить, переплетающуюся в клубок.
Методом авторадиографии установлено, что обмен РНК в ядрышках протекает наиболее интенсивно по сравнению со всеми остальными компонентами клетки. Электронномикроскопические исследования, проведенные в недавнее время, показали, что в клетках, активно синтезирующих РНК, ядрышки имеют типичное строение - они состоят из гранулярной и фибриллярной частей, организованных в виде нуклеолонемы. В тех клетках, где синтез РНК ингибирован (заторможен), ядрышки теряют гранулы и превращаются в мелкие плотные фибриллярные тельца. В среднем на РНК в ядрышках приходится приблизительно одна пятая часть их массы.
Ядрышки возникают в телофазе митоза на хромосомах, обладающих "спутником", непосредственно в месте вторичной перетяжки на нити, соединяющей хромосому со спутником (рис. 20). Потому число ядрышек в клетке оказывается равным количеству хромосом спутниками. На политенных гигантских хромосомах двукрылых было установлено, что ядрышки продуцируются определенными дисками хромосом.
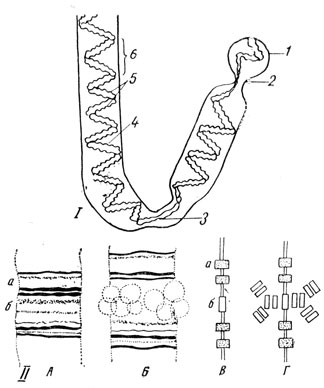
Рис. 20. I - схема строения хромосомы (из И. А. Алова, А. И. Брауде, М. Е. Аспиз). В хромосоме видны 2 хроматиды (5) и их спирали, большая (5) и малая (4); 1 - спутник; 2 - вторичная перетяжка; 3 - центромер (первичная перетяжка). II - схема образования ядрышка (по Штиху). А - участок гигантской хромосомы; а - с поперечными полосами, содержащими ДНК; б - с дисками, содержащими РНК. Б - образование вещества ядрышка на поперечной полосе, содержащей РНК. В - схема строения хроматиды: а - с генными участками, содержащими ДНК; б - с участком, содержащим РНК, который представляет собой 'организатор ядрышка'. Г - редупликация нуклеопротеида, содержащего РНК, на месте 'организатора ядрышка' - процесс, ведущий к возникновению видимого под микроскопом ядрышка
Примерно 20 лет тому назад была высказана идея, что нитчатые структуры ядрышка, когда оно исчезает в профазе митоза, концентрируются на хромосомах, располагаясь о поверхности хроматид. В анафазе, когда хроматиды расходятся по дочерним клеткам, фибриллярные элементы ядрышка оказываются распределенными поровну между ними. На стадии телофазы эти нитчатые структуры ядрышка объединяются с хромосомой в клубок. Затем восстанавливается нормальная структура ядрышка путем новообразования его второго нефибриллярного компонента при участии ядрышкового организатора, недавнее время этот цикл изменений ядрышка в процессе митоза был подтвержден и схематизирован на основании новых данных. Согласно указанной схеме, фибриллярное предъядрышко формируется в области ядрышкового организатора и при его участии начинает синтезировать р-РНК, строить рибосомы и накапливать гранулярный компонент. В итоге восстанавливается типичная структура нуклеолонемы.
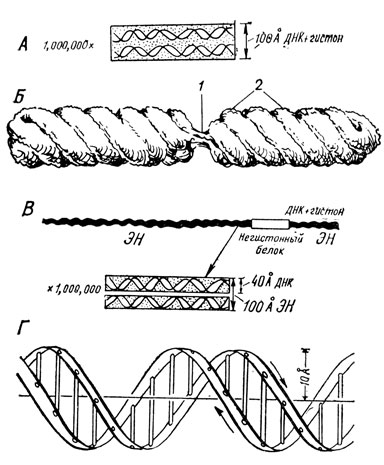
Рис. 21. Схемы молекулярной (А - по Рису) и субмикроскопической (Б - по Ю. С. Ченцову) организации хромосомы; 1 - центромерный участок; 2 - хромонемы (четыре). В - схема строения элементарной хромосомной нити (ЭН). Г - модель молекулы ДНК по Уотсону и Крику
Хромосомы, выявляющиеся в процессе перестройки ядра при митозе, согласно господствующим представлениям, построены из спирализованных элементарных нитей (рис. 21), имеющих диаметр 100 Å и объединенных в пучки - хромонемы. Число элементарных хромосомных нитей в хромонемах организмов разного вида различно.
Соединяясь в пары (или четверки), хромонемы образуют хроматиды. В хромосоме большие спирали хроматид располагаются параллельно друг другу. На рис. 21Б представлена одна из новых предложенных моделей хромосомы, образованной 4 хромонемами, то есть 2 хроматидами. Как выяснилось с помощью электронного микроскопа, хромосомы лишены оболочки.
Была высказана идея, получившая широкое распространение, что хромосомы, все более раскручиваясь в телофазе, переходят в интерфазу в деспирализованном виде и потому становятся неразличимыми. В профазе нитевидные хромосомы вновь начинают спирализоваться, и в метафазе плотность витков большой спирали достигает своего максимума. Из-за тесного прилегания друг к другу витков большой спирали закрученность хромосом на данной стадии митоза маскируется. Усиление спирализации и увеличение плотности залегания витков приводит к укорочению хромосомы до 1/12 и даже в некоторых случаях до 1/25 по сравнению с их длиной в профазе. Витки спирали начинают расходиться в анафазе, и тогда на некоторых объектах опять удается увидеть спирализацию хромосом. Обнаруживаемые на хромосомах хромомеры считают особенно резко спирализованными участками хромосом.
При деспирализации хромосом в интерфазе некоторые их участки остаются сильно спирализованными с плотным расположением витков. Эти участки назвали гетерохроматиновыми в отличие от подвергающихся деспирализации - эухроматиновых. На обычных гистологических препаратах в интерфазных ядрах гетерохроматиновые участки хромосом окрашиваются очень интенсивно. Они и дают картину хроматиновых глыбок. Комплексные генетические и цитологические исследования показали, что гены сосредоточены в эухроматиновых участках хромосом.
Большинство цитологов разделяет мнение, что каждая элементарная хромосомная нить (рис. 21В) является двойной - образована двумя еще более тонкими, перевивающимися друг с другом нитями диаметром 40 Å . Каждая такая нить представляет собой молекулу ДНК, имеющую вид двойной спирали, окутанную основным белком, у большей части животных - гистоном. Молекулы ДНК и гистона, взаимно спирализуясь, образуют дезоксирибонуклеопротеид (ДНП) нуклеогистон. Длина молекулы ДНП составляет 0,4 мк. Соединяясь друг с другом конец в конец и цементируясь в этих местах белком пока еще не уточненного состава (но не гистоном), молекулы ДНП образуют длинные нити, идущие на всем протяжении хромосом.
Молекулы ДНК являются носителями наследственных зачатков - генов. В них как бы записана генетическая информация, передаваемая от клетки к клетке, от поколения к поколению. Молекулы ДНК как носители наследственной информации не одинаковы по своим зачаткам - генам. У каждого вида животных и растений различаются между собой молекулы ДНК, входящие в состав разных хромосом хромосомного набора. В совокупности все разновидности молекул ДНК содержат полный геном организма, то есть весь состав его генов.
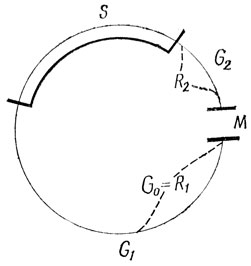
Рис. 22. Схема митотического цикла: G1 - постмитотический, или пресинтетический период; S - период синтеза ДНК; G2 - премитотический, или постсинтетический период; M - митоз
Сущность митоза состоит в тождественном распределении между дочерними клетками ДНК материнской клетки. Хромосомные нити, строящие разные хромосомы, несут различную генетическую информацию. Поскольку между дочерними клетками поровну распределяются хроматиды всех хромосом, каждая новая клетка получает весь набор генов, имевшихся в материнской клетке, но только в половинном количестве. Последующее удвоение количества наследственного материала, осуществляющееся путем удвоения - редупликации молекул ДНК, происходит в интерфазе в ее среднем периоде, названном синтетическим (S-период, или фаза синтеза). В интерфазе происходит подготовка к митотическому делению клетки, и весь этот процесс вместе с митозом обозначают как митотический цикл. Период интерфазы, предшествующий синтетическому, называют постмитотическим, или пресинтетическим (период g1, или фаза первого интервала), а последний период интерфазы - премитотическим, или постсинтетическим (период g2, или фаза второго интервала).
Среднее соотношение во времени между периодами интерфазы, а также между ними и митозом представлено на рис. 22. У организмов разных видов и разных тканей эти соотношения сильно варьируют. Например, у мыши время митотического цикла клеток эпителия кишечника составляет 19 часов, а эпителия кожи ее уха - 576 часов; в клетках надкостницы крысы оно равно 114 часам. Наиболее коротким периодом митотического цикла всегда является сам митоз. В клетках взрослых животных он продолжается большей частью 0,5 - 1 час. Очень быстро, в течение нескольких минут, протекают мета- и анафаза. Более длительны про- и телофаза, что обусловлено перестройкой в это время ядра клетки. Период g2 является наиболее коротким в интерфазе. Время редупликации ДНК чаще всего равно 6 - 8 часам. Наиболее продолжителен пресинтетический период g1.
Однако не все клетки даже активно пролиферирующих тканей постоянно находятся в состоянии митотического цикла, то есть в процессе митоза или подготовки к нему. Закончив митоз, они могут сразу же выйти из митотического цикла и приступить к дифференцировке и функционированию. Это состояние пролиферативного покоя обозначают как период G0 или стадия R1. Клетки могут выйти из митотического цикла и после синтетического периода, в этом случае они обладают удвоенным количеством ДНК. Клетки, еще не очень далеко зашедшие по пути дифференцировки, не потерявшие способности к дедифференцировке и пролиферации, могут под влиянием цитогенетического воздействия возвратиться к митотическому циклу. Клетки, находившиеся на стадии R2, то есть вышедшие из митотического цикла в премитотическом периоде, уже после возвращения к нему вступают в митоз. Интерфазу клеток, находящихся в митотическом цикле, называют аутосинтетической. Гетеросинтетической интерфазой обозначают состояние клеток в пролиферативном покое. Соматические клетки, о которых была речь, содержат в интерфазе, по сравнению со зрелыми половыми (сперматозоидами и яйцеклетками), двойное диплоидное количество ДНК.
Теория строения молекулы ДНК, предложенная Уотсоном и Криком и удостоенная в 1962 году Нобелевской премии как одно из величайших открытий современной биологии, позволяет легко расшифровать процесс редупликации ДНК, совершающийся в синтетическом периоде митотического цикла при подготовке клетки к делению. Согласно этой теории, молекула ДНК представляет собой как бы веревочную лестницу, спирально свернутую по своей длине (рис. 21Г). Продольные шнуры "лестницы" - это цепочки чередующихся молекул фосфорной кислоты (точнее - фосфатных остатков) и сахара - дезоксирибозы (рис. 23), причем против фосфатного остатка одного бокового шнура "лестницы" располагается фосфатный остаток другого, а против сахара одного шнура - сахар другого. Боковые части "лестницы" соединены перекладинами, лежащими через правильные интервалы около 3 - 4 Å. Перекладины присоединены непосредственно к молекулам сахара боковых шнуров особыми своими частями - продуктами азотистого обмена в организме, так называемыми пуриновыми и пиримидиновыми основаниями. Мы не будем касаться химической формулы их, весьма сложной, а дадим наименованиям буквенные обозначения. Это аденин (А), гуанин (Г), тимин (Т) и цитозин (Ц). Данные азотистые основания, обозначаемые еще нуклеотидами, располагаются вдоль спиральной молекулы ДНК всегда так, что против А залегает Т, против Г - Ц (рис. 23).
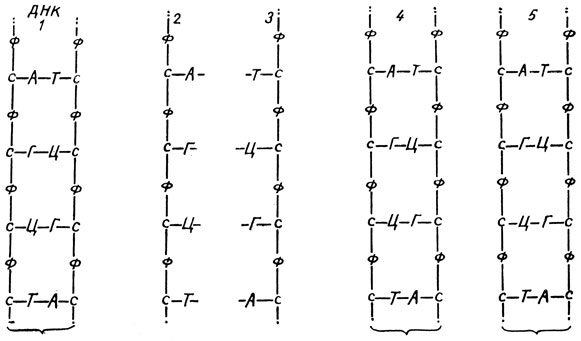
Рис. 23. Схема строения (1) и редупликации молекулы ДНК; 2 и 3 - разошедшиеся цепи двуцепочечной молекулы ДНК; 4 и 5 - новообразованные молекулы ДНК
В лестничных перекладинах эти основания - нуклеотиды соединяются друг с другом с помощью водородных связей.
Молекулы ДНК имеют огромную длину. Достаточно сказать, что в их состав входят десятки тысяч нуклеотидов, а молекулярный вес исчисляется десятками миллионов. Поэтому возможные варианты и комбинации упомянутых азотистых оснований - нуклеотидов по длине молекулы практически беспредельны. Общее же количество нуклеотидов в молекулах ДНК клетки человека составляет 10 миллиардов, что практически повышает до бесконечности количество возможных комбинаций азотистых оснований.
В процессе редупликации молекул ДНК водородные связи между нуклеотидами разрываются и двойная спираль разворачивается, давая две одинарные цепочки. Затем каждая из этих цепочек дает начало новой молекуле ДНК, строя вдоль себя вторую, дополняющую ее комплементарную цепочку. Так как расположение нуклеотидов в цепочке, формирующей молекулу ДНК, строго предопределяет их порядок в создаваемой цепочке по принципу комплементарности, дополнительности, то цепочка, обозначенная на схеме (рис. 23) цифрой 2, обязательно создаст, присоединяя необходимые компоненты (нуклеотиды, фосфорные остатки, молекулы дезоксирибозы) из окружающей среды, цепочку 3 и наоборот. В результате на основе цепочки 2 возникнет молекула 4, а на базе цепочки 3 - молекула 5. Обе новообразованные молекулы окажутся тождественными друг другу и материнской молекуле I. Из одной материнской молекулы ДНК образуются таким образом две ее точные копии. С помощью меченых атомов было выяснено, что в S-периоде редупликация молекул ДНК как в разных хромосомах, так и в разных участках одной хромосомы протекает неодновременно. Удвоение, редупликация молекул ДНК происходит с помощью особого фермента полимеразы.
В результате редупликации молекул ДНК удваивается количество элементарных хромосомных нитей. При наличии 2 хромонем в анафазных хромосомах, а следовательно, и в телофазе митоза и в постмитотическом периоде митотического цикла, в постсинтетическом периоде, а затем в про- и метафазе следующего митоза их будет 4.
На основании данных электронной микроскопии новообразование ядерных оболочек в дочерних клетках чаще всего связывают с эндоплазматическим ретикулумом. Считают, что оболочка ядра возникает из уплощающихся и сливающихся вместе пузырьков эндоплазматического ретикулума.
В 1953 году Гейтлер открыл особую форму митоза, которая получила название эндомитоза (по-гречески "эндо" означает внутри), или скрытого митоза. Его особенность состоит в том, что все превращения ядра протекают под сохраняющейся ядерной оболочкой. Естественно, что при этом в цитоплазме не формируется митотический аппарат. Проходя в типичном случае эндомитоза все фазы митоза и удваивая как количество ДНК, так и число хромосом, сама клетка не делится. Таким образом, эндомитоз приводит к полиплоидии (по-гречески "поли" означает много, "плоос" - складывать). В полиплоидных клетках не диплоидный набор хромосом и диплоидное количество ДНК, а больше. Довольно часто встречаются тетраплоидные клетки, содержащие 4 гаплоидных набора хромосом, то есть 2 диплоидных набора и удвоенное против нормы количество ДНК. При двух последовательных эндомитозах возникают октоплоидные и т. д., вплоть до тысячи "n" и более.
Одним из результатов эндомитоза является политения, о которой была речь во второй главе, когда говорилось о гигантских хромосомах в ядрах слюнных желез двукрылых.
Известно, что существует постоянство отношений между объемом ядра и цитоплазмы. Так называемое ядерно-цитоплазменное отношение различно у организмов разного вида и в разных тканях. Полиплоидия соматических клеток приводит к соответствующему увеличению размеров клеточного тела. В результате повторных эндомитозов возникают гигантские полиплоидные клетки. Полиплоидные клетки являются физиологически более работоспособными и в этом отношении представляют преимущества для тканей и органов, в которых они встречаются. Возникновение полиплоидных клеток в тканях можно вызвать специальными воздействиями, например, радиационным излучением. В настоящее время метод полиплоидизации широко используют в сельскохозяйственной практике с целью повышения продуктивности разных культур (сахарной свеклы, винограда и др.).
Полиплоидия может выражаться и в двуядерности либо многоядерности клеток. Исследования, проведенные после открытия эндомитоза, показали, что картины амитоза обычно связаны с разделением ядра в полиплоидных клетках, которое может сопровождаться и перетяжкой клеточного тела. Суммарное количество ДНК в подобных двуядерных клетках или возникающих из них двух одноядерных, соответственно кратно увеличено, причем дезоксирибонуклеиновая кислота может быть распределена между ядрами с разной степенью неравномерности. По-видимому, при наличии в ядре нормального количества ДНК клетка, образовавшаяся таким способом, может далее размножаться митозом.
Образование дву- или многоядерности, осуществляющейся путем перешнуровки материнского ядра, академик А. А. Заварзин рассматривал как процесс приспособительного характера, ведущий к увеличению поверхности взаимодействия между ядром и цитоплазмой и способствующий функциональной активности клетки. Что же касается тех видов амитоза, которые получили наименование фрагментации и почкования, то они оказались свойственными лишь отмирающим клеткам.
Таким образом, на основании всей совокупности известных данных амитоз не может рассматриваться как полноценная форма клеточного деления.
Сопоставление по целому ряду признаков митоза, эндомитоза и амитоза приводится в таблице 1.
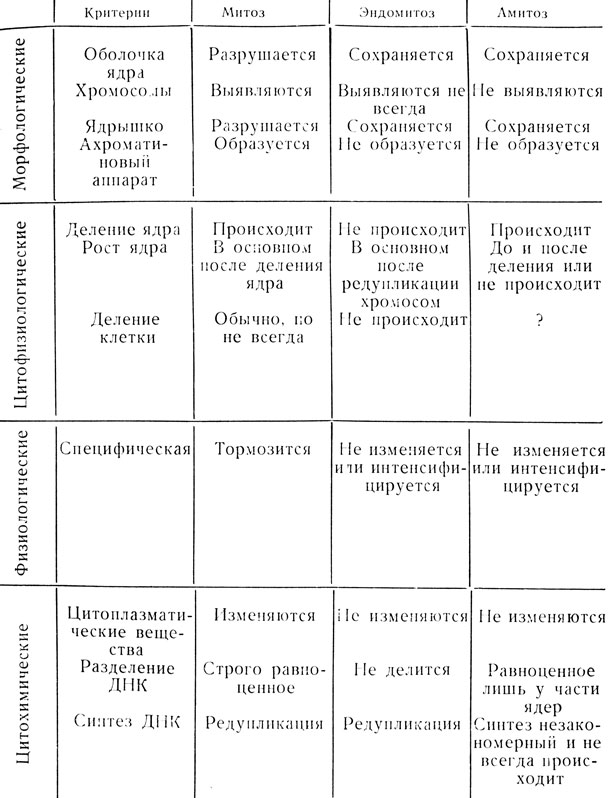
Таблица 1. Сравнительная характеристика митоза, эндомитоза и амитоза в соматических тканях позвоночных (по В. Я. Бродскому, 1966)
При использовании материалов сайта активная ссылка обязательна:
http://anfiz.ru/ 'Анатомия и физиология человека'