Глава II. История учения о клетке
Первое представление о клетке возникло в науке более трех веков назад. В 1665 году английский физик Роберт Гук опубликовал в книге "Микрографии" результаты своих наблюдений, выполненных на сконструированном им примитивном микроскопе. Среди прочих предметов он описал и зарисовал тонкий срез через пробку - кору пробкового дуба. Согласно этим описаниям, вещество пробки наполнено воздухом, который заключен в небольших коробках-ячейках, или клетках (cells, cellula), плотно прилегающих друг к другу и напоминающих пчелиные соты. Аналогичное ячеисто-клеточное строение было обнаружено Гуком и при изучении под микроскопом срезов стеблей различных растений. Он открыл наличие в клетках сока, имеющего, по его млению, питательное значение.
Почти через два столетия после открытия Гука в 1830 году крупнейший чешский ученый прошлого века Ян Пуркине установил наличие в клетке пузырьковидного ядра, а в 1831 году англичанин Роберт Броун описал ядро в растительных клетках и признал его постоянной структурой клетки. Ему же принадлежит термин Nuckleus (ядро). Однако ни Броун, ни многие ученые после него не придавали ядру существенного значения и рассматривали его как уплотненную слизь или как питательное вещество.
Изучение тонкого строения растительных и животных организмов стало бурно развиваться после изобретения более совершенных ахроматических микроскопов. В отличие от старых микроскопов, страдавших сферической и хроматической аберрацией и дававших поэтому расплывчатое изображение, окрашенное во все цвета радуги, новые оптические приборы давали четкое и ясное изображение. Введение их в практику исследовательских лабораторий вызвало появление многочисленных работ с описанием тонкого строения организмов, в частности различных частей тела растений: листьев, лепестков, цветков, стеблей, корней.
Были выявлены клетки во многих органах тела животных. Главная заслуга в этом принадлежит Пуркине и его школе. Пуркине обозначал клетки шариками или зернами. В 1836 году один из учеников Пуркине - Валентин впервые описал в ядре ядрышко. Суммируя накопленный материал, Пуркине выступил в 1837 году с "теорией зернышек", согласно которой зернышки, подобные клеткам растений, являются структурными элементами тела животных. Но он еще не был создателем клеточной теории. Честь ее формулирования принадлежит немецкому зоологу Теодору Шванну, опубликовавшему в 1839 году свою знаменитую книгу "Микроскопическое исследование о соответствии в структуре и росте животных и растений". В нейлон на большом фактическом материале обосновал идею о принципиальном сходстве клеток животных и растительных организмов: все ткани животных, как и растений, построены из клеток и им присущ единый принцип развития всех структур на основе клеток.
С тех пор клеточная теория стала одной из незыблемых основ современного естествознания. Фридрих Энгельс, придавая клеточной теории громадное значение в науке, ставил ее наряду с законом превращения энергии и дарвиновской теорией эволюции органического мира. Клеточная теория, то есть теория клеточного строения и развития тела животных и растений, подтверждала единство происхождения всего живого на Земле. Множество мельчайших, видимых только под микроскопом животных и растений тоже оказалось построенным по клеточному принципу. Эти простейшие типы растений и животных представляют собой свободно живущие клетки-организмы.
Говоря о значении клеточной теории и выявившейся роли клеток, как основных структурных элементов тела растительных и животных организмов, Ф. Энгельс подчеркивал чрезвычайную важность для развития биологии открытия клетки - "той единицы, из размножения и дифференцирования которой возникают и вырастают все организмы, за исключением низших. Только со времени этого открытия стало на твердую почву исследование органических, живых продуктов природы - как сравнительная анатомия и физиология, так и эмбриология. Покров тайны, окутывавший процесс возникновения и роста структуры организмов, был сорван. Непостижимое до того времени чудо предстало в виде процесса, происходящего согласно тождественному по существу для всех многоклеточных организмов закону"1.
1 (Ф. Энгельс. Диалектика природы. М., 1955, стр. 155.)
На современном этапе развития науки установлено, что в теле человека лишь одна пятая - одна шестая часть его массы приходится на клетки. Большая часть человеческого тела состоит из надклеточных (вся скелетная мускулатура - мышцы) или межклеточных (кости и хрящи скелета, связки, сухожилия и тому подобное) структурных элементов. Однако все эти надклеточные и межклеточные структуры развиваются из клеток или за их счет. Таким образом, если понимать клеточную теорию не буквально, как теорию клеточного строения, а с учетом высказанной еще Шванном идеи об общем принципе развития всех структур растительного и животного тела на основе клеток, то она остается бесспорной и в настоящее время.
Современная клеточная теория рассматривает клетку как главнейшую форму существования жизни на Земле, учитывая при этом, что, во-первых, клетки многоклеточного организма отнюдь не являются индивидуумами, способными существовать самостоятельно, а представляют собой лишь части целого, строго подчиненные его разностороннему регулирующему влиянию; во-вторых, клетки - это не единственная форма жизни, так как наряду с клетками имеются доклеточные (вирусы, бактериофаги, сине-зеленые водоросли, лишенные обособленного ядра) и надклеточные (разного рода симпласты, содержащие много ядер в одном протоплазматическом теле) формы живого.
Интересно отметить, что основоположник клеточной теории Теодор Шванн, как и его предшественники, считал важнейшей частью клетки оболочку, а не содержимое. И только в 1861 году Макс Шультце вывел оболочку из состава обязательных частей животной клетки. Однако и в его время еще не было выяснено значение клеточного ядра. Это следует из развернутого определения клетки, принадлежащего Шультце: "Клетка является комочком протоплазмы, внутри которого лежит ядро. Клетка характеризуется полноценностью жизненных отправлений, субстратом которых является преимущественно протоплазма. Однако также и ядро играет значительную, хотя все же до сих пор неизвестную роль".
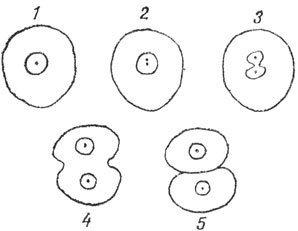
Рис. 3. Прямое деление клетки по Ремаку
Большое значение для утверждения клеточной теории имело открытие клеточного деления. Еще до появления клеточной теории Шванна были известны данные о делении растительных клеток в результате образования перегородки в материнской клетке. Однако состояние клеточного ядра при этом не учитывалось. В 1841 - 1852 годах Ре-мак дал подробное описание прямого деления животных клеток. Согласно Ремаку, оно совершается в три этапа (рис. 3): 1) происходит разделение ядрышка, в котором появляется посредине перетяжка, а потом оно в этом узком месте разрывается; 2) вслед за этим подобная перетяжка появляется в ядре, и оно постепенно принимает вид песочных часов; в дальнейшем благодаря полной перешнуровке ядро превращается в два пузырька равных размеров; 3) происходит аналогичная перетяжка клеточного тела, и в результате из одной материнской клетки возникают две дочерние, общий объем которых равен объему клетки, давшей им начало.
Позже были обнаружены способы прямого деления ядер путем образования перегородки (ядерной пластинки) или односторонней перетяжки (при подковообразном ядре). При этом прямое деление ядра часто связано не с клеточным делением, а с образованием дву- или многоядерных клеток. Были открыты еще две другие разновидности прямого деления клеток. В одном случае в результате разделения материнской возникают дочерние клетки равных размеров, но число их велико. То есть ядрышко разделяется не на две, а на много частей, и соответственно этому на столько же фрагментов делятся ядро и клеточное тело. Этот способ прямого деления получил наименование фрагментации. В другом случае появляются две дочерние клетки, но очень разные по своим размерам. Создается впечатление, что материнская клетка как бы отпочковывает от себя небольшую дочернюю клетку. Такой способ деления называется почкованием. В настоящее время установлено, что и почкование, и фрагментация не представляют собой нормального способа клеточного размножения, а встречаются лишь в дегенерирующих и гибнущих клетках.
Рудольф Вирхов (1859) возвел факт клеточного деления в общий принцип, провозгласив "всякая клетка от клетки". Он имел в виду возникновение клеток в онтогенезе - индивидуальном развитии организмов - и вовсе не касался вопроса об историческом происхождении клетки. Сформулированный Вирховым закон "всякая клетка от клетки" сохраняет свою силу и поныне.
Постепенно шло накопление данных, касающихся другой формы клеточного деления, получившего название непрямого. Первоначально было замечено, что во время деления клетки ее ядро часто исчезает, как бы растворяется, а затем опять появляется в дочерних клетках. В 1871 году русский ботаник И. Д. Чистяков зарисовал отдельные стадии непрямого деления растительных клеток, а в 1873 году зоолог Шнейдер, изучавший дробящиеся клетки червя Mesostomum Ehrenbergeri, дал уже в динамике описание последовательных стадий непрямого деления.
Наиболее полное и детальное исследование непрямого деления было выполнено Флеммингом (1879). Он установил, что в ядре делящейся клетки возникают сильно окрашивающиеся нити - хромосомы (по-гречески "хрома" - краска и "сома" - тело), которые затем расщепляются, а образовавшиеся половинки хромосом расходятся к полюсам клетки и из них формируются ядра дочерних клеток (рис. 4). Исходя из перестройки ядра в процессе непрямого деления в нитевидные образования, Флемминг назвал всю динамику ядерного метаморфоза митозом (по-гречески "митос" - нить), а непрямое деление клетки в целом кариокинезом ("карион" - ядро, "кинезис" - движение). В настоящее время оба эти термина употребляются как синонимы.
В 1885 году Рабль установил, что каждый вид растительных и животных организмов имеет постоянное число хромосом во всех клетках его тканей, а в 1888 году Бовери обнаружил факт постоянного повторения морфологических особенностей каждой хромосомы (индивидуальность хромосом) в хромосомном наборе клеток тела. В целом количество хромосом в наборе и их индивидуальные особенности создают для каждого вида растительных и животных организмов особый тип ядра - кариотип.
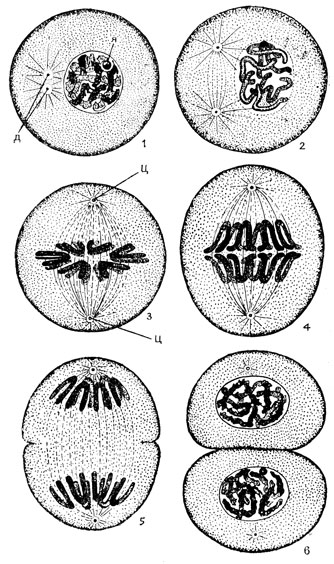
Рис. 4. Митотическое деление клетки: 1 - ранняя профаза; 2 - поздняя профаза; 3 - метафаза; 4 - ранняя анафаза; 5 - поздняя анафаза (стадия дочерних звезд); 6 - телофаза. Я - ядрышко, Ц - центриола, Д - диплосома
Наряду с уточнением картины митоза в последней четверти прошлого столетия были открыты различные структуры - органоиды или органеллы в цитоплазме: клеточный центр, хондриосомы, внутриклеточный сетчатый аппарат. Первым из органоидов был обнаружен клеточный центр, названный центросомой. Его описали Ван Бенден и Гертвиг (1875). Просто организованный клеточный центр состоит из двух коротеньких палочек - центриолей, имеющих длину около 0,5 мк. В световом микроскопе они имеют вид гранул, расположенных взаимно перпендикулярно в светлом шарообразном пространстве - центросфере. Иногда центросома имеет более сложное строение: ее периферия образована лучистой сферой, или астросферой - системой тонких радиальных нитей, исходящих из центросферы.
В интерфазе, или интеркинезе, как теперь обозначают период между митозами, клетка обладает ядром, оптически пустым в прижизненном состоянии. В нем видны лишь оболочка и одно-два ядрышка. Кариоплазма (нуклеоплазма) находится в жидком состоянии и если механически разрушить ядерную оболочку, то она вытекает в окружающую цитоплазму, но не смешивается с ней. При применении обычных фиксаторов, убивающих клетку, происходит коагуляция ее живого вещества, в том числе кариоплазмы, и в ядре появляются глыбки разного размера. Вследствие способности сильно окрашиваться (они проявляют базофилию - окрашиваются основными, ядерными красителями) глыбки получили название хроматиновых. Лишь при фиксации осмиевой кислотой картина напоминает прижизненную, так как протоплазма образует в этих условиях очень нежную зернистость.
Вступив в митоз, клетка перестает специфически функционировать и округляется. В первой фазе митоза - профазе начинается расхождение центриолей клеточного центра и образование вокруг них фигуры звездчатого сияния, а между ними ахроматинового, то есть плохо красящегося веретена (рис. 4). Одновременно происходит перестройка ядра клетки. В ранней профазе в ядре выявляются хромосомы, которые вначале лежат плотным клубком. В это время еще сохраняются ядерная оболочка и ядрышко. В поздней профазе хромосомы уже хорошо различимы и часто удается видеть, что каждая из хромосом "расщеплена" по длине на две одинаковые половинки - хроматиды. Оболочка и ядрышко постепенно исчезают.
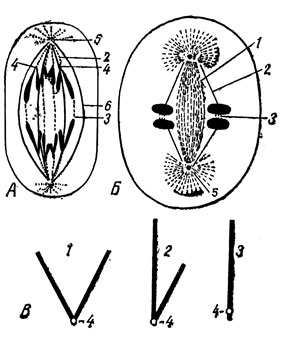
Рис. 5. А - митотический аппарат животной клетки на стадии анафазы и Б - схема строения веретена: 1 - опорные волокна; 2 - хромосомная нить; 3 - межзональные волокна; 4 - центромер; 5 - центриола; 6 - мантийное волокно. В - различные типы хромосом (анафазные хромосомы): 1 - метацентрическая; 2 - субметацентрическая; 3 - акроцентрическая; 4 - центромеры
Во второй фазе митоза - метафазе - центриоли устанавливаются по полюсам клетки и веретено достигает своего максимального развития. К концу метафазы отчетливо расщепленные хромосомы оказываются расположенными в области экватора клетки и образуют вместе экваториальную пластинку. Хромосомы изогнуты в форме шпилек или подковок, концы которых направлены к периферии. На хромосомах обычно более или менее отчетливо заметны перетяжки, называемые центромерами, или кинетохорами (рис. 5А). К этим участкам хромосом прикрепляются нити веретена, называемые хромосомными нитями (рис. 5А и 5Б). В зависимости от того, где лежат центромеры, хромосомы принято разделять на метацентрические (рис. 5В), субметацентрические и акроцентрические ("акрос" - край).
В центре веретена, в пространстве, отграниченном хромосомами, залегают опорные нити. Непрерывные нити, идущие от одного полюса веретена к другому, могут обнаруживаться и по его периферии. Если посмотреть на клетку со стороны одного из ее полюсов, то весь набор хромосом будет похож на звезду. Поэтому данную стадию непрямого деления клетки называют стадией материнской звезды. Она наиболее удобна для изучения кариотипа клетки. На стадии материнской звезды особенно отчетливо видно, что каждая из хромосом состоит из двух тождественных друг другу хроматид. Каждая хроматида в свою очередь состоит из двух более тонких нитей - полухроматид, субхроматид, или хромонем, которые располагаются в хроматиде так, как хроматиды в хромосоме.
В третьей фазе митоза - анафазе происходит разделение надвое центромеров во всех хромосомах, то есть отделение хроматид друг от друга и расхождение их от экватора клетки в сторону центриолей. Хроматиды становятся при этом сестринскими хромосомами, а их хромонемы соответственно хроматидами. Таким образом, профазные и метафаэные хромосомы состоят из 4, а анафазные - из 2 хромонем.
Движение хроматид к полюсам клетки происходит центромерами вперед. Поэтому его обычно объясняют растягиванием хроматид друг от друга укорачивающимися хромосомными нитями. На этом основании хромосомные нити веретена получили еще одно название - тянущих нитей. Передвигаясь в направлении полюсов клетки, хромосомы скользят вдоль опорных нитей веретена. Всю совокупность этих ахроматиновых, желеобразно уплотненных нитей обозначают как митотический аппарат клетки. В результате его деятельности каждая из дочерних клеток получает тождественный набор хромосом.
Поздняя анафаза характеризуется значительным расхождением дочерних наборов хромосом от экватора материнской клетки, где в это время может уже начаться перетяжка клеточного тела (процесс, обозначаемый как плазмотомия, или цитокинез, и ведущий в итоге к разделению материнской клетки на две дочерних). Эту стадию называют стадией дочерних звезд.
Последняя, четвертая фаза митоза - телофаза - по существу обратна профазе. Хромосомы постепенно становятся все менее заметными и в конце концов их уже не удается обнаружить. Ядро окружается оболочкой, и в нем появляется ядрышко. В телофазе заканчивается плазмотомия и на месте материнской клетки теперь имеются две дочерних, суммарный объем которых равен объему клетки, давшей им начало. Некоторое время еще можно видеть остаточное веретено - продукт обратного развития митотического аппарата.
В интерфазе клетка вырастает до объема материнской и готовится к новому митотическому делению либо проделывает настолько высокую дифференцировку, что уже становится неспособной к митозу.
Еще Флемминг описал многочисленные утолщения на протяжении хромосом. Эти узелки на хромосомах называются хромомерами. Хромомеры по своему расположению, величине и форме весьма специфичны. Они строго постоянны во всех однозначных хромосомах данного кариотипа и являются морфологическим выражением индивидуальности хромосом в их наборе.
Вскоре после уточнения картины митоза многие авторы (Ру, 1883; Гертвиг, 1884; Страсбурге; 1884; Геккель, 1886) высказали мысль о том, что ядро клетки является носителем наследственности. С этих позиций становилось понятным расщепление хромосом на две тождественные половинки и точное распределение этих половинок в анафазе между дочерними клетками. В 1902 году Сеттон выдвинул хромосомную теорию наследственности, использовав цитологические данные для расшифровки механизма менделевского расщепления наследственных зачатков. Так было положено основание новой ветви цитологии - цитогенетике.
Большую роль в дальнейших цитогенетических исследованиях сыграли гигантские хромосомы двукрылых насекомых. Эти многонитчатые хромосомы (рис. 6) образуются путем многократного удвоения хромонем (иногда число их увеличивается в 512 раз), не сопровождающегося их расхождением и увеличением количества хромосом. При этом соответственно возрастает и объем цитоплазмы. Поскольку все хромонемы располагаются параллельно друг другу и однозначные хромомеры оказываются лежащими на одном уровне, то в гигантской хромосоме они образуют в совокупности диски. Количество дисков равно числу хромомер в обычных хромосомах. Толщина дисков строго отвечает размерам соответствующих хромомер, а расстояние между дисками - промежуткам между хромомерами. Использование гигантских хромосом в качестве объекта изучения представляло для исследователей большие удобства и оказалось плодотворным. Сочетая генетические исследования с тонким изучением хромосом, проводившимся на плодовой мушке - дрозофиле и гигантских хромосомах ее слюнных желез, Морган создал в 1911 году карты линейного расположения в хромосомах данных организмов наследственных зачатков - генов.
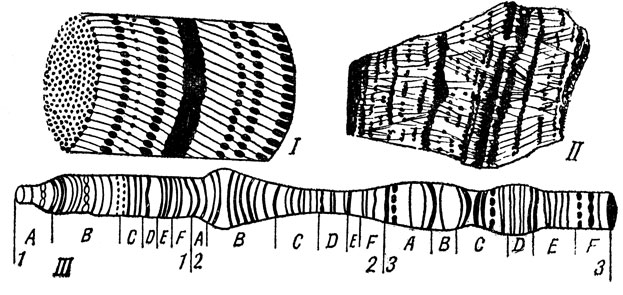
Рис. 6. Строение политенных гигантских хромосом: 1 - параллельное расположение хромонем и залегание однозначных хромомеров на одном уровне (схема). Возникновение в результате этого картины поперечных дисков (II), III - линейное расположение дисков (генов и групп генов) вдоль гигантской хромосомы дрозофилы
Вторые по времени открытия органоиды клетки - хондриосомы (по-гречески "хондрос" - зерно) были фактически впервые описаны Альтманом (1890). Однако он не понял их истинного значения. Обнаруженные в цитоплазме клеток гранулы Альтман гомологизировал с микроорганизмами - кокками и назвал биобластами (он рассматривал биобласты как мельчайшие структурные компоненты клеточного тела). Бенда подробно изучил хондриосомы в развивающихся мужских половых клетках мыши (1894). В зависимости от формы хондриосом им давали различные названия, а всю совокупность в клетке обозначали как хондриом. В настоящее время стало общепринятым наименование хондриосом как митохондрий, наиболее отвечающее их морфологии (рис. 7).
В одной и той же клетке форма и размеры митохондриев могут очень сильно варьировать, тем более в клетках разных тканей различных животных. Изменяются они и в зависимости от функционального состояния клетки. Чаще всего они имеют толщину около 1 мк, а длину до 7 мк. При использовании специальных методов исследования митохондрии могут быть обнаружены и прижизненно. Установлено, что они способны к передвижению по клеточному телу.
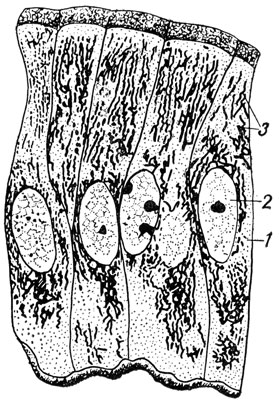
Рис. 7. Митохондрии в клетках кишечного эпителия крысы: 1 - цитоплазма; 2 - ядро; 3 - митохондрии
Теперь хорошо известно значение митохондриев как клеточных органоидов. Митохондрии - органоиды дыхания. В них происходит окисление органических веществ и за счет освобождающейся энергии строятся макроэргические соединения, в первую очередь аденозинтрифосфорная кислота - АТФ. Поэтому митохондрии образно называют силовыми станциями клетки. Они всегда концентрируются в тех местах клетки, где имеется наибольшая потребность в поступлении энергии. Во время митоза митохондрии приблизительно поровну распределяются между дочерними клетками.
Последний органоид общего характера - внутриклеточный сетчатый аппарат был открыт Гольджи в 1898 году в нервных клетках спинальных ганглиев (рис. 8) и получил впоследствии наименование аппарата Гольджи. Размеры его и расположение зависят от видовых особенностей животных, характера ткани, функционального состояния клетки. Чаще всего он лежит над ядром, около клеточного центра, или вокруг ядра.
Долгие годы ученые не могли расшифровать значения этого сетчатого образования. И только в 1923 году советский ученый Д. Н. Насонов впервые высказал идею о функциональном значении аппарата Гольджи. На основании собственных экспериментов и литературных данных Д. Н. Насонов пришел к заключению, что внутриклеточный сетчатый аппарат является органоидом аккумуляции, то есть накопления веществ, синтезируемых клеткой.
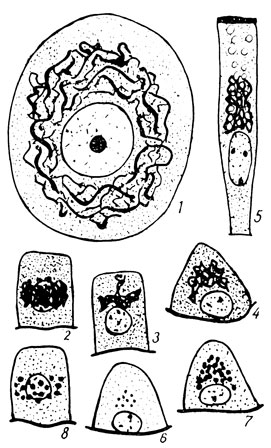
Рис. 8. Аппарат Гольджи в клетках: 1 - нервной; 2 - 5 - железистых; 6 - начало отложения; 7 - 8 - накопление гранул витального красителя в железистых клетках
Особенно наглядно выступала роль аппарата Гольджи в железистых клетках. Здесь было отчетливо видно, как первые мельчайшие капельки секрета появляются и постепенно накапливаются в петлях аппарата Гольджи. Затем они выходят из сети аппарата в цитоплазму клетки и далее выводятся из нее. Было установлено, что в аппарате Гольджи аккумулируются вещества, резорбируемые клеткой, например, жир, в клетках кишечного эпителия. Процесс появления и конденсации веществ в сети аппарата Гольджи удалось проследить путем введения в организм животного прижизненного красителя, например, трипанового синего. Краска эта мало ядовита и спустя некоторое время после ее инъекции удается обнаружить, особенно в железистых клетках, гранулы краски, по своему распределению в клетке повторяющие расположение в ней внутриклеточного сетчатого аппарата. Теория Д. Н. Насонова получила вскоре многочисленные подтверждения в Советском Союзе и за рубежом и широко вошла в цитологию.
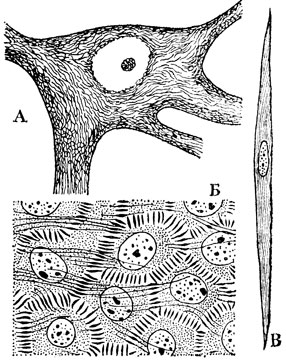
Рис. 9. Метаплазматические структуры, или специальные органоиды клеток: А - неврофибриллы в нервной клетке; Б - тонофибриллы в многослойном плоском эпителии вагины женщины; В - миофибриллы в клетке - волокне гладкой мышечной ткани
Помимо органоидов общего характера оптический микроскоп дал возможность обнаружить ряд специальных органоидов, присущих лишь клеткам определенных тканей и выполняющих частные функции. Их называют также метаплазматическими структурами (рис. 9). Так, в мышечных волокнах были найдены сократительные волоконца - иофибриллы, в нервных клетках - нейрофибриллы и в клетках других тканей опорные волоконца - тонофибриллы, располагающиеся вдоль силовых линий. На свободной поверхности некоторых эпителиальных клеток были обнаружены реснички, осуществляющие волнообразные мерцательные движения (рис. 10А и 11А). У основания мерцательных ресничек располагаются в ряд особые базальные тельца, по одному на каждую ресничку, а от них в глубь клетки тянется опорный конус из корневых нитей. Если разрезом нарушить связь ресничек с базальными тельцами, мерцание прекращается.
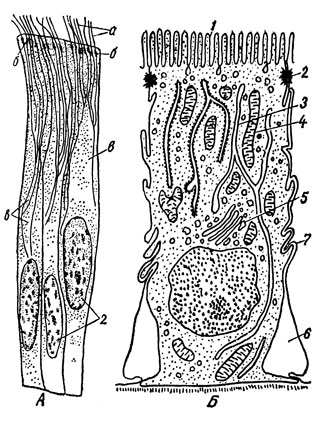
Рис. 10. А - мерцательные цилиндрические эпителиальные клетки из кишечника беззубки: а - реснички; б - базальные тельца; в - опорный конус из корневых нитей; г - ядра клеток. Б - схема электронномикроскопического строения каемчатой клетки эпителия тонкой кишки: 1 - микроворсинки; 2 - десмосома; 3 - гладкий эндоплазматический ретикулум; 4 - митохондрия; 5 - комплекс Гольджи; 6 - межклеточная щель; 7 - 'замки', скрепляющие соседние клетки пласта
Наряду с органоидами общего и специального характера в клетке могут встречаться еще параплазматические образования, то есть включения - продукты клеточной жизнедеятельности (рис. 12А и Б). Плотные включения носят название гранул, жидкие - вакуолей. Они могут быть разной природы: белковые (гранулы различного секрета, подлежащего выведению из клетки), углеводные (глыбки животного крахмала - гликогена, резервного вещества), липидные (жировые капли в жировых клетках соединительной ткани). Некоторые включения подобны специальным органоидам в том отношении, что являются постоянными структурами, обеспечивающими способность данных клеток выполнять свойственные им функции - например, зерна пигмента в хроматофорах соединительной ткани.
И органоиды клетки, и включения лежат в гомогенной, бесструктурной под световым микроскопом части цитоплазмы, носящей название гиалоплазмы (стекловидной плазмы).
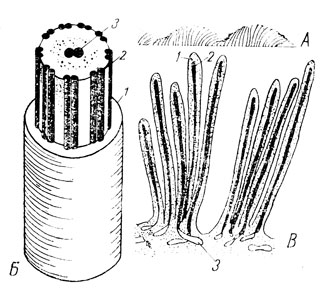
Рис. 11. А - волны мерцания ресничек. Б - схема электронномикроскопического строения реснички: 1 - плазмолемма; 2 - периферические фибриллы (9 пар); 3 - центральные фибриллы (1 пара). В - схема электронномикроскопического строения каемки эпителия тонкой кишки обезьяны. Кустиковое расположение микроворсинок; 1 - микроворсинка; 2 - микроканалец; 3 - каналец эндоплазматической сети
Клетки находятся в теснейшем и непрерывном взаимодействии с окружающей их средой, из которой в них проникают различные вещества, необходимые для поддержания их жизнедеятельности и функции, и в которую поступают из клетки продукты ее обмена. Морфологическим выражением активного захвата клеткой микроскопически видимых плотных частиц является фагоцитоз (по-гречески "фагос" означает пожирать), впервые описанный крупнейшим отечественным ученым И. И. Мечниковым в 1883 году. По И. И. Мечникову, в процессе фагоцитоза следует различать 4 этапа: активное движение фагоцита к пожираемому объекту, тесное сближение с ним (аттракция), поглощение и переваривание.
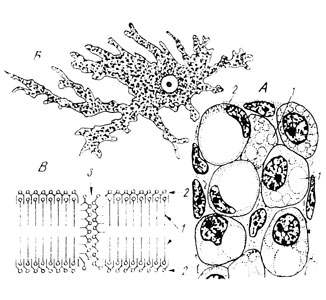
Рис. 12. А - жировые включения: 1 - в развивающихся жировых клетках; 2 - сформировавшаяся жировая клетка. Б - пигментные меланиновые зерна в пигментной клетке брыжейки тритона. В - схема молекулярной организации цитоплазматической мембраны: 1 - бимолекулярный слой липидов; 2 - мономолекулярные слои белка; 3 - пора (по Даниелли)
При использовании материалов сайта активная ссылка обязательна:
http://anfiz.ru/ 'Анатомия и физиология человека'