Мозжечок
Подобно тому как кора мозговых полушарий управляет реакциями, возникшими из внешних впечатлений (экстероцептивные реакции), так до известной степени мозжечок управляет реакциями, возникшими из внутренних впечатлений (проприоцептивные реакции). По существу мозжечок является большим проприоцептивным центром координации. Он связан приносящими волокнами с первичным проприоцептивным аппаратом спинного и стволовой части головного мозга. Он также тесно связан с мозговой корой при помощи корково-мостового пути.
"Центры корреляции мозгового ствола, - говорит Геррик*, - и по преимуществу центры корреляции коры мозга анализируют входящие в мозг импульсы и определяют те частные реакции, которые являются подходящими для каждого данного положения. После того как этим путем был определен характер движения, проприоцептивные системы содействуют его выполнению. Центральной координирующей станцией для проприоцептивных реакций является мозжечок. Ни одна из его деятельностей не проникает в сферу сознания".
* (An introduction to neurology, Philadelphia and London, 1922.)
Главнейшей функцией мозжечка в настоящее время считается его синергическая деятельность. Под словом "синергия" понимают способность организма комбинировать сравнительно простые, физиологически обособленные движения в более или менее сложные специальные акты*. Всякая синергическая мышечная деятельность зависит от двух факторов: во-первых, от установления и поддержания во всем теле синергических мышечных единиц и, во-вторых, от установления и поддержания координации между синергическими единицами при выполнении сложных мышечных актов.
* (Подробный анализ этого понятия имеется в работе Тильней, "The form and functions of the central nervous system", London, 1923.)
Каждая синергическая единица составляется из двух мышц: действующей (агониста) и противодействующей (антагониста). По существу обе эти мышцы являются синергистами, ибо постоянно действуют одновременно, выполняя одну, строго координированную работу. В этой работе агонист является доминирующим элементом, определяющим направление движения, а антагонист, который всегда работает вместе с агонистом, - элементом задерживающим, стремящимся к тому, чтобы результирующее движение ни на один момент не оказалось чрезмерным, а всегда точно приспособленным к определенной цели. Когда надлежащее соотношение между этими двумя элементами потеряно, движение становится неправильным, например, разгибание может сделаться чрезмерным, если не будет надлежащего противодействия со стороны сгибателя. Вся мышечная система туловища, шеи, конечностей, языка, жевательного аппарата и т. д. состоит из подобных синергических единиц.
При выполнении таких простых актов, как, например, сгибание или разгибание кисти, требуется участие лишь одной синергической единицы, но если мы имеем дело с более сложными актами, то в действие вводится целый ряд синергических единиц, между которыми должны быть установлены соответствующие строго координированные соотношения. Рассмотрим пример, заимствованный мной у Тильней. Мы берем со стола стакан с водой, подносим его к губам, выпиваем воду и ставим стакан обратно на стол. Здесь принимает участие целый ряд синергических единиц кисти, предплечья, плеча, плечевого пояса и т. д. Все эти единицы должны быть строго координированы между собой в смысле одновременности или последовательности их действия, в смысле надлежащего соотношения в силе и объеме производимых движений. Каждая из синергических единиц, введенная в выполнение вышеуказанных актов, может во время своего действия находиться то в кинетической, то в акинетической фазе; так, в определенный момент та или другая мышца будет сокращаться, чтобы вызвать соответствующее движение, - например, разгибание пальцев руки и их последовательное сгибание для того, чтобы захватить стакан; в другой момент, раз стакан захвачен, пальцы для удержания стакана остаются фиксированными и неподвижными, хотя мышцы пальцев находятся в активном сокращении. Первая фаза та, которая производит движение, есть кинетическая фаза; вторая, - которая обусловливает неподвижное положение в силу активного сокращения мышц, есть акинетическая фаза. Эти две фазы каждой синергической единицы должны быть приведены в соответствие во времени, например, пальцы не должны быть сжаты раньше того момента, как стакан будет захвачен. Сила мышечных сокращений в синергических единицах должна быть строго согласована с целью действия как в кинетических, так и в акинетических фазах. Объем движения должен быть равным образом регулирован так, чтобы выполнить цель акта; иначе расстояние, которое пройдет движущаяся часть, будет неправильным.
Синергическая деятельность может касаться или одного только отдела конечности, или всей конечности, или нескольких конечностей, или туловища и конечностей. Трудно даже себе представить, как сложна должна быть синергическая деятельность в таких сложных движениях, как, например, игра в теннис, когда приходится пустить в дело чуть ли не все синергические единицы тела и поддерживать между ними надлежащие соотношения (интегративная или сложная синергия).
Приносящие волокна от спинного мозга и от стволовой части головного входят в мозжечок через его верхнюю и нижнюю ножки. Средние ножки мозжечка представляют собой систему волокон, идущих от базальных ядер моста (nuclei pontis) к противоположному полушарию мозжечка. Базальными ядрами моста называют скопления серого вещества, разбросанные в вентральном отделе моста среди поперечно идущих его волокон. Волокна от ассоциационных центров мозговой коры (лобного, теменного и затылочно-височного) приходят в ядра моста корково-мостовыми путями (tracti cortico-pontini), а от двигательной области мозговой коры при помощи коллатеральных ветвей корково-спинального тракта (tractus cortico-spmalis), отходящих от него в момент прохождения через мост. Таким образом волокна средней ножки мозжечка (brachium pontis), возникающие из базальных ядер моста передают нервные импульсы, пришедшие от коры мозга, коре противоположного полушария мозжечка.
Выносящие волокна оставляют мозжечок через все три ножки, направляясь к двигательным центрам стволового отдела головного мозга - tracti cerebello-tegmentales. Наибольшее число волокон через верхнюю ножку мозжечка направляется к красному ядру (nucleus ruber). От красного ядра волокна идут вниз в спинной мозг (tractus rubro-spinalis) и вверх к коре мозга.
Одни из мозжечковых связей перекрещены раньше вхождения в мозжечок, другие входят в мозжечок неперекрещенными, но могут, конечно, перейти на противоположную сторону уже в самом мозжечке. К последним принадлежат tractus spino-cerebellaris dorsalis и tractus vestibulo-cerebellaris.
Раньше вхождения в мозжечок совершает перекрест в передней (вентральной) комиссуре спинного мозга большая часть волокон tractus spino-cerebellaris ventralis.
Корково-мостовый путь идет от коры мозга к базальным ядрам моста по своей собственной стороне, а уже от этих ядер начинается новое звено, которое по средней мозговой ножке направляется в противоположное полушарие мозжечка
Оливо-мозжечковый тракт (tractus olivo-cerebellaris) начинается от нижней оливы, переходит на противоположную сторону в межоливном пространстве, пронизывает противоположную оливу и затем через нижнюю ножку входит в мозжечок. По словам Тильней, этот тракт координирует движения головы с движениями глаз. Задние поверхностные дуговые волокна (fibrae arcuatae superf lciales posteriors), начинающиеся от nucleus fasciculi gracilis и nucleus fasciculi cuneati, входят в нижнюю ножку мозжечка той же самой стороны.
Передние поверхностные дуговые волокна происходят из нижних отделов nucleus fasciculi gracilis и nucleus fasciculi cuneati, перекрещивают срединную плоскость и вступают в нижнюю ножку мозжечка противоположной стороны. Большинство передних поверхностных волокон оканчивается в дуговом ядре (nucleus arcuatus), расположенном на вентральной поверхности пирамид, тотчас же выше перекреста. Из этого ядра начинается новое звено, которое идет далее вместе с непрерывающимися передними дуговыми поверхностными волокнами в соответствующую нижнюю ножку. У верхнего конца продолговатого мозга nucleus arcuatus увеличивается в объеме и продолжается в серое вещество вентрального отдела моста, Если правильно допущение, что nuclei arcuati получают коллатерали от пирамидных трактов коры мозговых полушарий, то исходящие из этих ядер волокна нужно считать гомологичными поперечным волокнам варолиева моста, В состав передних дуговых поверхностных волокон входят также волокна, начинающиеся в nucleus reticularis lateralis.
Импульсы, приносимые в мозжечок различными путями, могут не только координировать, но, по-видимому, также и усиливать соматические двигательные механизмы. В мозжечковых нейронах содержится большое количество потенциальной нервной энергии, за счет которой мозжечок и производит стимулирующий или тонический эффект па мышцы тела.
Мозжечковая кора отличается от мозговой коры формой и расположением своих нейронов и тем, что по своей структуре мозжечковая кора одинакова на всем ее протяжении. Мозговая же кора по форме и расположению своих нейронов оказывается различной в различных областях, что связано с областными локализациями различных функций.
Физиологическое влияние мозжечка на движение совершенно отлично по характеру от влияния мозговой коры. Мозговая кора, в ее отношении к произвольным движениям имеет как главный объект определенные цели движения, определенные двигательные задачи, тогда как синергический контроль мозжечка имеет своей основной функцией регуляцию мышечных механизмов, необходимых для выполнения данных двигательных задач.
Каждое простое синергическое движение имеет в коре мозжечка свой специальный центр, каждое сложное синергическое достижение - свою специальную зону. Ввиду сложности действий мозжечка его двигательные зоны можно наметить лишь в общих чертах, а не с такой точностью, как произвольные двигательные центры мозговой коры.
У позвоночных животных мозжечок является постоянной составной частью головного мозга, хотя и подверженной резким изменениям в степени своего развития. Животные, способные лишь к простым, ограниченным движениям, обладают небольшим, несложным мозжечком; животные же, способные к обширным и сложным движениям, обладают большим, высоко развитым мозжечком. В своей простейшей форме мозжечок является в виде двух небольших дорзально-латеральных выступов на metencephalon, связанных позади IV желудочка центральной дуговой частью.
Наиболее простую форму мозжечка мы отмечаем у круглоротых, отличающихся очень малой подвижностью. У селахий (акулы, скаты), обладающих значительной быстротой и силой движения, мозжечок является уже на более высокой ступени развития. У ганоидных и костистых рыб картина приблизительно та же, что и у селахий, так как двигательная способность указанных рыб имеет много общего с двигательной способностью селахий.
У амфибий, благодаря их малой подвижности, мозжечок мал и, подобно мозжечку круглоротых, очень простой конструкции.
У рептилий, за исключением крокодила, Аллигатора и черепахи, мозжечок очень невелик. У перечисленных, более активных форм он относительно большего размера.
У птиц в связи со способностью летания центральная часть мозжечка (центральная дуга) сильно развивается, образуя настоящий червячок.
У млекопитающих центральная дуга становится еще более выраженной, чем у птиц, и появляются полушария мозжечка, которые в сущности представляют собой разросшиеся передние отделы дорзально-латеральных выступов элементарного metencephalon. Из заднего отдела этих выступов развиваются flocculus и paraflocculus.
Червячок, полушария, flocculus и paraflocculus являются основными частями мозжечка млекопитающих. Эти анатомические подразделения органа вариируют у различных групп млекопитающих соответственно двигательным нуждам животного. У человека paraflocculus имеется в рудиментарном состоянии и представляет собой небольшие скопления серого вещества позади ножки flocculus. У многих млекопитающих paraflocculus выражен резче, чем flocculus.
Наибольшего своего развития мозжечок достигает у человека в силу чрезвычайной подвижности и сложности движений его руки и в силу чрезвычайной сложности синергического контроля мышц туловища и нижних конечностей при стоянии и хождении на двух ногах.
Объем варолиева моста и его ножек у различных млекопитающих пропорционален объему мозжечковых полушарий, которые в свою очередь вариируют в соответствии с величиной коры мозга. Особенно тесную связь с развитием мозговой коры имеют те части мозжечка, которые филогенетически появляются позднее других; это - lobus ansiformis и lobus paramedianus. По номенклатуре, принятой у Зернова, lobus ansiformis соответствует: lobus posterior superior, lobus posterior inferior и lobus cuneiformis; lobus paramedianus соответствует tonsilla.
Эта зависимость в развитии указанных частей мозжечка от развития коры мозга связана, по-видимому, с синергической координацией произвольно вызванных движений. Весьма возможно, что синергический контроль мозжечка совершенно не зависит от способа возбуждения движений, т. е. он остается тем же, будут ли движения произвольными или рефлекторными. Словом, на мозжечок возлагается детальное выполнение актов, начатых другими центрами. Такими центрами могут быть: 1) или рефлекторные центры стволовой части мозга, когда дело идет о врожденных, типичных, однообразно повторяющихся действиях, или 2) кора мозговых полушарий, когда дело идет о личном опыте, прошедшем или настоящем, и когда даже может быть поставлено veto для реакций, возникающих в низших центрах (Геррик).
В коре мозжечка млекопитающих находится известное число координирующих центров, из которых одни парные, другие непарные. Парные центры помещаются в полушариях, непарные - в червячке. Каждый непарный центр предназначен для двусторонних координированных движений парных мышечных групп. Непарные центры управляют движениями головы, глаз, рта, челюстей, языка, гортани, глотки, туловища. К непарным центрам червячка, по-видимому, относятся также и центры конечностей, заведующие одновременным согласованным действием мышечных групп конечностей, например, в актах ходьбы или стояния. Для каждой конечности той или другой стороны имеется, кроме того, соответствующий центр в одноименном полушарии мозжечка, предназначенный для самостоятельных, независимых движений данной конечности.
Верхние конечности способны производить акты, в которых их взаимные движения координированы одни по отношению к другим, но в известных актах принимает участие только одна верхняя конечность, независимо от другой верхней конечности, которая остается бездеятельной или даже может производить движения, совершенно отличные от первой. Те же самые соображения, правда, в меньшей степени, приложимы и к нижним конечностям. Можно допустить поэтому, что мышечные группы конечностей имеют в коре мозжечка два центра, - один непарный срединный для двусторонних синергических движений, другой парный (двусторонний) для самостоятельных движений какой-либо конечности одной стороны.
Чтобы легче разобраться в функциональных особенностях отдельных участков коры мозжечка, я привожу здесь в виде таблицы анатомические соотношения отдельных долек червячка с отдельными долями мозжечковых полушарий. Номенклатура базельская (рис. 205 и 206).
| Верхний червячок | Полушарие мозжечка |
|---|---|
| 1) Lingula (язычок) | 1) Vincula (связи язычка) |
| 2) Lobulus centralis (центральная долька) | 2) Ala lobuli centralis (крыло центральной дольки) |
| 3) Culmen monticuli (верхушка горки) | 3) Pars anterior lobuli quadrangularis (передняя часть четырехугольной дольки) |
| 4) Declive monticuli (скат горки) | 4) Pars posterior lobuli quadrangularis (задняя часть четырехугольной дольки) |
| 5) Folium vermis (лист червячка) | 5) Lobulus semilunaris superior (верхняя полу лунная долька). |
| Нижний червячок | Полушарие мозжечка |
| 6) Tuber vermis (бугор червячка) | 6) Lobulus semilunaris inferior (нижняя полулунная долька) |
| 7) Pyramis (пирамида) | 7) Lobulus biventer (paraflocculus dorsalis) (двубрюшная долька) |
| 8) Uvula (язычок) | 8) Tonsilla (paraflocculus ventralis) (миндалина) |
| 9) Nodulus (узелок) | 9) Flocculus (velum medullare post.) (клочок) |
На сагиттальном разрезе, проведенном через середину мозжечка (рис. 207), хорошо видно, двигательным контролем каких мышечных групп заведуют отдельные дольки червячка. На рис. 205 и 206, изображающих верхнюю и нижнюю поверхности мозжечка, указано, контролем каких мышечных групп заведуют отдельные участки мозжечковых полушарий.
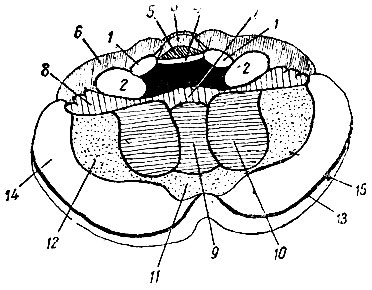
Рис. 205. Схема вентральной и нижней поверхности мозжечка. 1 - braehium conjunctivum eerebelli (ножка мозжечка к четверохолмию); 2 - braehium pontis; 3 - lobulus centralis; 4 - velum medullare anticum; 5 - lingula; 6 - pars anterior lobuli quadrangularis; 7 - nodulus; 8 - flocculus; 9 - uvula; 10 - tonsilla (миндалина); 11 - pyramis; 12 - lobulus biventer; 13 - fissura horizontalis; 14 - lobulus semilunaris inferior; 15 - lobulus semilunaris superior
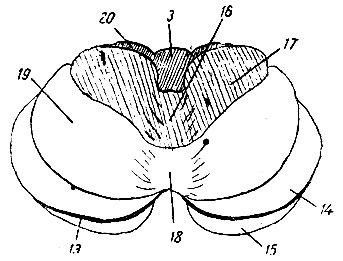
Рис. 206. Схема верхней поверхности мозжечка. 16 - culmen monticuli; 17 - pars anterior lobuli quadrangularis; 18 - deciive monticuli; 19 - pars posterior lobuli quadrangularis; 20 - ala lobuli centralis
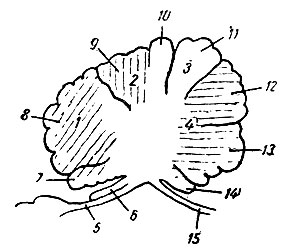
Рис. 207. Сагиттальный разрез через червячок человеческого мозжечка (из Геррика). 1 - участок, контролирующий движения в области головы (косые штрихи); 2 - участок, контролирующий движения шеи (вертикальные штрихи); 3 - участок, заведующий контролем одновременных координированных движений парных конечностей (не заштрихован); 4 - участок, контролирующий движения туловища (горизонтальные штрихи); 5 - velum medullare anterius; 6 - lingula; 7 - lobulus centralis; 8 - culmen monticuli; 9 - declive monticuli; 10 - folium vermis; 11 - tuber vermis; 12 - pyramis; 13 - uvula; 14 - nodulus; 15 - tela chorioidea ventriculi quarti
Lobulus centralis и culmen monticuli вместе с соответствующими им долями полушарий (ala lobuli centralis и pars anterior lobuli quadrangularis) заведуют движениями глаз, лицевой мускулатуры, челюстей, языка, глотки и гортани.
Сильное развитие указанных центров как в продольном, так и поперечном направлениях, особенно резко выраженное у человека, стоит, по-видимому, в связи с развитием мимики и речи. Deciive monticuli вместе cpars posterior lobuli quadrangularis заведуют контролем движений шейных мышц. Эта область особенно сильно развита у животных, где мышцы шеи играют очень важную роль в статике головы, например, у жирафы, или где, кроме статических заданий, имеется еще большое разнообразие произвольных движений головы, например, у человека; у животных, где шейные мышцы играют слишком малую роль в статике и динамике головы, указанная область или развита слишком слабо (например, у тюленя), или почти отсутствует совершенно (например, у китообразных).
Кзади от описанной выше области на верхне-задней поверхности червячка в folium и tuber vermis помещается непарный центр для синергической деятельности конечностей правой и левой стороны. Указанный отдел червячка особенно развит у животных, конечности которых действуют почти исключительно синергическим образом на обеих сторонах, например, у лошади, барана, оленя, антилопы, тапира. Центры для контроля самостоятельных односторонних движений конечностей помещаются в мозжечковых полушариях: в lobulus semilunaris superior, а частью, по-видимому, и в lobulus semilunaris inferior - для верхней конечности, в lobulus semilunaris inferior и в lobulus biventer - для нижней.
Помещающиеся в полушариях парные центры конечностей будут преобладать в своем развитии над непарным центром конечностей, помещающимся в червячке, у тех животных, у которых отдельные конечности приобрели значительную независимость в своих движениях, например, у лазящих, роющих, водных. Наибольшего развития парные центры конечностей достигают у человека, где имеется наибольшее разнообразие и наибольшая самостоятельность движений каждой отдельной конечности. Непарный центр конечностей (folium vermis и tuber vermis) у человека выражен довольно слабо, так как синергическое движение конечностей, особенно верхних, у человека довольно ограничено.
На нижней поверхности червячка, в pyramis, uvula и nodulus, располагается срединный центр для движений туловища, тесно связанный с сохранением равновесия, по-видимому, через посредство ядер крыши. Указанные части червячка, заведующие движениями туловища, дыхательными движениями, мускулатурой промежности, очень мало вариируют у различных групп млекопитающих ввиду сравнительного однообразия их деятельности у различных групп.
Paraflocculus у человека рудиментарен, и ножка его не может быть прослежена до червячка. У животных развитие flocculus и parafloccuius, по-видимому, стоит в тесной зависимости от развития мускулатуры хвоста. Какое значение они имеют у человека - неизвестно.
Кора мозжечка представляет собой массу серого вещества, равномерно расположенную как на поверхности полушарий, так и на поверхности червячка. На разрезе кора мозжечка представляется состоящей из двух слоев: 1) наружного - молекулярного и 2) внутреннего - зерновидного (рис. 208). На границе между этими слоями помещается ряд клеток Пуркинье. Каждая клетка Пуркинье, имеющая грушевидную форму, дает от своей верхушки толстый протоплазматический отросток, который почти у самого своего основания начинает древовидно разветвляться, направляясь через всю толщу молекулярного слоя к поверхности. Область, в которой происходит разветвление древовидных отростков клеток Пуркинье, располагается перпендикулярно к длинной оси листочков (folium) мозжечка. Распространение этой области по длинной оси листочков очень ограничено. Тонкие аксоны клеток Пуркинье направляются внутрь, в массу белого вещества; они покрыты миэлином и представляют главный выводящий путь из коры мозжечка. Аксоны клеток Пуркинье оканчиваются в глубоких мозжечковых ядрах (главным образом в nucleus dentatus), откуда начинается новое выносящее звено.
Зерновидный слой, расположенный тотчас же кнаружи от центрального белого вещества, состоит из маленьких зерновидных клеток, обладающих более или менее значительным ядром, окруженным небольшим количеством цитоплазмы. От каждой такой клетки отходят 3-5 коротких протоплазматических отростков (дендритов) с когтеобразными окончаниями, при помощи которых зерновидные клетки устанавливают контакты с мшистыми волокнами (о которых будет сказано ниже); своим внешним видом эти контакты напоминают glomeruli обонятельного нерва. Безмиэлиновый аксон зерновидной клетки, направляясь к поверхности, проникает в молекулярный слой. Здесь он делится на две ветви, идущие параллельно длинной оси листочков и последовательно проникающие через области распространения древовидных отростков клеток Пуркинье, с которыми аксоны зерновидных клеток, по-видимому, устанавливают синоптические связи.
Молекулярный слой мозжечка, особенно в своем более глубоком отделе, содержит так называемые корзиночные клетки. Помимо нескольких протоплазматических отростков, более или менее выраженных, каждая такая клетка обладает типичным аксоном, идущим в молекулярном слое в плоскости, перпендикулярной к длинной оси листочков. Каждый аксон на своем пути дает многочисленные коллатерали, отходящие под прямыми углами и направляющиеся от поверхности к центру. Концевые разветвления аксонов и коллатералей имеют форму корзинок, в которых помещены тела клеток Пуркинье.
Приносящие волокна могут оканчиваться в мозжечке двояким образом: 1) или как ползучие волокна, приходящие в непосредственную связь с дендритами клеток Пуркинье, или 2) как мшистые волокна, устанавливающие связь с зерновидными клетками, которые уже в свою очередь передают импульсы клеткам Пуркинье.
Рамоп-Кахаль полагает, что мшистые волокна являются окончанием приносящих волокон нижней ножки мозжечка, а ползучие - окончанием приносящих волокон средней ножки.
Так как каждое приносящее волокно нижней мозговой ножки сильно ветвится и достигает своими разветвлениями большего или меньшего числа зерновидных клеток, разбросанных на более или менее значительных расстояниях друг от друга, и так как аксоны каждой зерновидной клетки достигают дендритов очень большого числа клеток Пуркинье, то единичный импульс, входящий через отдельное нервное волокно, может возбуждать очень большое число клеток Пуркинье, и физиологический эффект, таким образом, будет значительно усилен. Такое же усиление эффекта может быть получено при помощи корзиночных клеток, из которых каждая может разряжать импульс сразу на несколько клеток Пуркинье.
Весьма возможно, что это усиление стимулов во время передачи их через кору мозжечка имеет значение как для поддержания двигательных реакций вообще, так и для поддержания мышечного тонуса в частности. Этой прибавочной энергией от мозжечка пользуются не только рефлекторные движения, но и произвольные.
Что касается анатомических основ синергической деятельности, надо думать, что те ползучие волокна, которые ограничиваются в своем распространении лишь областью дендритов одной клетки Пуркинье, ограничиваются регуляцией одной синергической единицы. Волокна корзиночных клеток, соединяющие вместе несколько клеток Пуркинье, обусловливают синергический контроль для целой группы синергических единиц. Зерновидные клетки, связующие значительное количество клеток Пуркинье, производят контроль значительного количества синергических единиц, может быть, даже всего тела (Тильней).
Весьма вероятно, что все волокна, начинающиеся в коре мозжечка, прерываются в его ядрах. Исключение, по-видимому, составляют волокна, направляющиеся непосредственно от коры мозжечка к ядру Дейтерса в продолговатом мозгу.
Итак, почти все выносящие волокна мозжечка начинаются в его ядрах. Так как каждое зубчатое ядро мозжечка (nucleus dentatus) получает волокна от соответствующего полушария мозжечка, то становится очевидным, что соответствующая ножка мозжечка к четверохолмию (brachium conjunctivum) является главным выносящим путем для данного мозжечкового полушария (рис. 209). Равным образом волокна, начинающиеся в коре червячка, оканчиваются в ядрах покрышки (nuclei fastigii), откуда берет начало главный выносящий пучок червячка (tractus fastigio-bulbaris). Некоторые из выносящих волокон ядер покрышки выходят через среднюю ножку мозжечка (tractus cerebello-tegmentalis pontis).
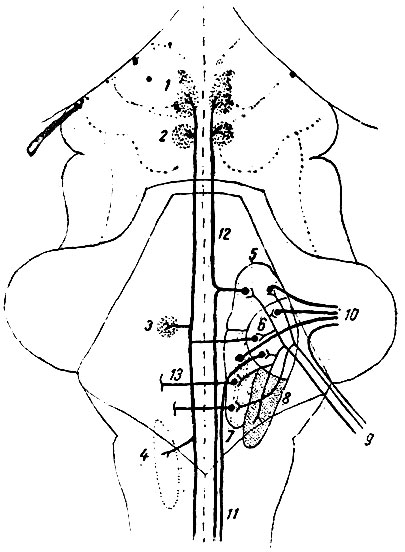
Рис. 209. Схема ядер вестибулярного нерва и некоторых из связанных с ними путей (из Геррика. Видоизменено). 1 - ядро глазодвигательного нерва; 2 - ядро блоковидного нерва; 3 - ядро отводящего нерва; 4 - двойное ядро (nucleus ambiguus); 5 - верхнее ядро вестибулярного нерва (ядро Бехтерева); 6 - боковое ядро вестибулярного нерва (ядро Дейтерса); 7 - срединное ядро вестибулярного нерва (ядро Швальбе); 8 - спинальное ядро вестибулярного нерва; 9 - вестибулярный нерв (нерв преддверия); 10 - tractus vestibulo-cerebellaris; 11 - tractus Vestibulo-spinalis; 12 - fasciculus longitudinalis medialis; 13 - одно из волокон, связующих ядра вестибулярного нерва с двигательными ядрами и ретикулярным образованием продолговатого мозга
При использовании материалов сайта активная ссылка обязательна:
http://anfiz.ru/ 'Анатомия и физиология человека'