Полосатое тело и зрительный бугор
Кора мозга связана выносящими проекционными волокнами с зрительным бугром и с красным ядром и с corpus striatum (рис. 202).
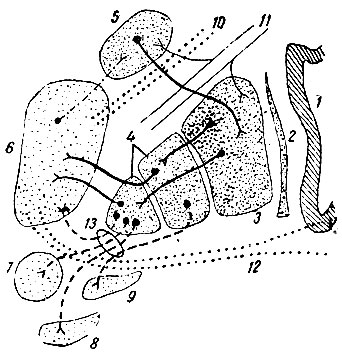
Рис. 202. Схема связей nucleus caudatus и nucleus lenticularis (по Рансону). 1 - insula; 2 - claustrum; 3 - putamen; 4 - globus pallidus; 5 - nucleus caudatus; 6 - thalamus; 7 - nucleus ruber; 8 - nucleus hypothalamicus (corpus Luysi); 9 - substantia nigra; 10 - radiatio thalamica anterior; 11 - tractus cortico-spinalis; 12 - pedunculus thalami inferior (вентральный пучок); 13 - ansa lenticularis
Corpus striatum, как известно, состоит из nucleus caudatus и nucleus lenticularis (lentiformis) вместе с разделяющими их пучками волокон внутренней капсулы.
Nucleus lentiformis в свою очередь при помощи наружной и внутренней пластинок белого вещества делится на три зоны. Наиболее боковая и в то же время наиболее обширная зона называется putamen,a две медиальных зоны, вместе взятые, образуют globus pallidus. Заложенный в globus pallidus важный двигательный центр связан с низшими двигательными нейронами передних рогов спинного мозга при помощи tractus strio-rubralis (до красного ядра) и tractus rubro-spinalis (от красного ядра до передних рогов). Возможно также, что выносящие импульсы от globus pallidus доходят до передних рогов спинного мозга через substantia nigra. Аксоны клеток, находящихся в substantia nigra, направляются в ретикулярное образование среднего мозга, а оттуда, по-видимому, начинается новый пучок, который и доходит до передних рогов. Таким образом, первая двигательная цепь от corpus striatum до передних рогов будет состоять из двух звеньев: 1) tractus strio-rubralisn 2) tractus rubro-spinalis; вторая же цепь - из трех звеньев: 1) tractus strio-nigralis, 2) tractus nigro-reticularis и 3) tractus reticulo-spinalis. Рассматриваемые нами двигательные тракты - rubro-spinalis и vestibulo-spinalis, по-видимому, являются первоначальными трактами, проводящими двигательные импульсы от более высоких центров к низшим, и лишь сравнительно поздно возник у млекопитающих двигательный путь, идущий непосредственно от коры головного мозга к передним рогам спинного мозга, - tractus cortico-spinalis s. pyramidalis.
Globus pallidus представляет собою примитивное corpus striatum и поэтому называется также paleostriatum. Putamen и nucleus caudatus представляют собой более позднее приобретение, которое, по-видимому, оказывает влияние на деятельность более примитивного отдела corpus striatum, т. е. на globus pallidus.
Corpus striatum млекопитающих рассматривают как гомолог базального узла переднего мозга (ganglion basale) низших позвоночных, у которых этот узел образует собой значительный отдел переднего мозга; он дает начало объемистому пучку волокон, так называемому базальному пучку переднего мозга (fasciculus basilaris). Базальный пучок у рыб, земноводных, пресмыкающихся и птиц представляет собой систему волокон, которая, очевидно, является гомологом двигательного пути человека. Базальный двигательный пучок, занимающий у низших форм базальное положение, направляясь кзади через; промежуточный, средний и задний мозг, вступает, в конце концов, в спинной мозг. По всей вероятности, этот базальный пучок, помимо выносящих двигательных волокон, содержит также и приносящие чувствующие. У животных, не обладающих мозговой корой, как, например, у рыб, fasciculus basilaris является единственным двигательным путем, связующим высшие отделы центральной нервной системы с низшими сегментальными отделами. Но по мере того, как начинает развиваться кора мозга, над старым двигательным центром в базальном узле начинает развиваться новый двигательный корковый центр. Новый двигательный центр создает и новый двигательный путь (tractus pyramidalis). В результате мышцы туловища (соматические) оказываются под коптролем двух путей: 1) старого, идущего от corpus striatum, и 2) нового, идущего от коры мозга.
На базальный узел переднего мозга рыб смотрят как на предшественника paleostriatum, т. е. globus pallidus. Nucleus caudatus и putamen, образующие вместе neostriatum, посылают свои нервные импульсы в globus pallidus, который в свою очередь посылает импульсы в вентральный отдел thalamus, в субталамическую область и в tegmentum среднего мозга. Nucleus caudatus nputamen производят регулирующее, задерживающее влияние на двигательные функции globus pallidus. Повреждение nucleus caudatus и putamen характеризуется дрожанием я судорожными движениями, тогда как повреждение globus pallidus характеризуется дрожанием и ригидностью мышц. В общем система corpus striatum действует тонизирующим, укрепляющим образом на произвольные движения.
Neostriatum впервые появляется у рептилии, т. е. тогда, когда переход от водной жизни к наземной уже окончательно установился. По-видимому, новые условия окружающей среды вызвали необходимость новых, специально наземных двигательных приспособлений. У птиц и млекопитающих neostriatum усиливается в своем развитии, причем у млекопитающих волокнами внутренней капсулы оно подразделяется на два отдела: 1) nucleus caudatus и 2) nucleus lentiformis.
Globus pallidus состоит почти исключительно из больших пирамидальных клеток (двигательные клетки), тогда как nucleus caudatus и nucleus lentiformis состоят преимущественно из мелких клеток, дающих начало волокнам, соединяющим различные части corpus striatum между собой. Большинство этих волокон идет от nucleus caudatus к putamen, а начавшиеся в putamen идут к globus pallidus. Среди мелких клеток neostriatum лишь изредка попадаются крупные клетки, по-видимому, однородные с клетками globus pallidus. Центробежные волокна начинаются главным образом в globus pallidus; они собраны в поперечно идущий пучок (ansa lenticularis), распределяющийся в thalamus, в красном ядре, гипоталамическом ядре (ядро Льюиса) и в substantia nigra. Другая часть волокон, принадлежащая к той же самой системе, проникает в зрительный бугор через вентральный отдел внутренней капсулы. Приносящие волокна достигают nucleus caudatus от переднего и срединного ядер зрительного бугра.
Зрительный бугор на своей верхней поверхности покрыт тонким слоем белого вещества (stratum zonale), от которого вниз, в массу серого вещества, спускается вертикальная пластинка (lamina medullaris interna), расщепленная у своего верхне-переднего конца на 2 ножки. Эта пластинка белого вещества делит серое вещество зрительного бугра на 3 отдельных участка: 1) срединное ядро, 2) боковое ядро и 3) переднее ядро (рис. 96). Срединное медиальное ядро помещается между центральным серым веществом III желудочка и lamina medullaris interna. Кзади срединное ядро доходит до области habenula, занимая задний отдел боковой стенки III желудочка. Боковое ядро является самым большим из 3 ядер зрительного бугра. Оно помещается между внутренней и наружной медуллярными пластинками.
Наружная медуллярная пластинка расположен на наружной поверхности зрительного бугра в непосредственном соседстве с волокнами внутренней капсулы. Кзади боковое ядро распространяется значительно далее срединного, включая в свой состав всю pulvinar. Боковое ядро в свою очередь подразделяется на ряд вторичных ядер: собственно боковое ядро, вентральное ядро, заднее ядро и т. д.
Переднее ядро, ограниченное расщеплением передне-верхнего отдела laminae medullaris internae, является наименьшим из 3 первичные ядер зрительного бугра. Оно образует передний бугорок thalami и в виде клина проникает на короткое расстояние книзу и кзади между передними отделами бокового и срединного ядер. Субталамическая область представляет собой продолжение tegmentum под задний отдел зрительного бугра. При помощи subthalamus мозжечок приходит в связь со зрительным бугром через промежуточную станцию в красном ядре. Таким образом устанавливается tractus cerebello-rubro-thalamicus, который, весьма возможно, продолжается и далее от зрительного бугра в кору мозга.
Тильней полагает, что клинически такое допущение должно считаться весьма вероятным, особенно для той части мозжечкового аппарата, которая имеет отношение к ориентировке в пространстве. Subthalamus является также промежуточной станцией для выносящих путей от corpus striatum.
Зрительный бугор с точки зрения его эволюции делят на 2 отдела: 1) древний зрительный бугор (paleothalamus)и 2) новый зрительный бугор (neothalamus). В состав paleothalamus входит следующие ядра, расположенные вокруг полости III желудочка: 1) nucleus anterior, 2) nucleus medialis, 3) centrummedianum (Льюис) и 4) ganglion habenulae. Centrum medianum, принадлежащее к нижне-медиальному отделу thalamus представляет собой массу серого вещества округлой формы, расположенную между nucleus medialis, nucleus lateralis и pulvinar. Своим наружным отделом centrum medianum прилежит к lamina medullaris interna, а верхне-внутренним сливается cnucleus medialis, часть которого, в сущности, оно и представляет. Эти ядра дифференцировались для первичных таламических корреляций, каковые мы наблюдаем у рыб и у других низших позвоночных. По-видимому, некоторые из функций древнего thalamus сохранились и у человека. Весьма вероятно, что paleothalamus у низших позвоночных является органом, предназначенным для очень высоких степеней чувствующей корреляции. Здесь собираются импульсы слуховой, зрительной, обонятельной, вкусовой, общей сомэстезической и висцеральной чувствительности. При известных благоприятных условиях соответствующая корреляция этих чувствующих импульсов создает ощущение благополучия. По-видимому, из этого состояния благополучия или из нарушений его и возникают первоначальные импульсы для поведения данного животного. Расстройства чувства благополучия могут обусловливаться или недостаточной, или чрезмерной, или несоответственной стимуляцией, получаемой через какой-либо из путей специальной или общей чувствительности. Так, например, чрезмерная стимуляция какого-либо участка кожи расстраивает чувство благополучия и вызывает двигательные реакции с целью уйти от раздражителя или удалить раздражитель. Чувство голода, например, также расстраивает чувство благополучия. Чувство голода обусловливается известной комбинацией соматических и внутренностных чувствующих импульсов, которая и вызывает двигательные реакции, необходимые для добывания пищи. Без изменений в ощущении благополучия и обусловленных этими изменениями соответствующих двигательных реакций животное умерло бы с голода.
Клинический опыт, говорит Геррик, показывает, что многие из примитивных функций paleothalamus сохранились и у человека. Так, аффективная сторона наших ощущений, по-видимому, не зависит от мозговой коры, а связана с деятельностью thalamus. Весьма вероятно, что и висцеральными приспособлениями также управляет не мозговая кора, а thalamus.
Neothalamus, наиболее выраженный у млекопитающих, представляет собой боковой отдел зрительного бугра. Ядра, образующие neothalamus, служат промежуточной станцией для волокон спинального, бульбарного и тройничного лемнисков и передают приносимые этими волокнами импульсы конечным чувствующим областям в коре мозга.
Pulvinar и corpus geniculatum laterale служат промежуточной станцией для волокон зрительного тракта, который, отсюда направляется к затылочной области коры. Corpus geniculatum laterale впервые появляется у селяхий и затем, постепенно увеличиваясь, достигает наибольшего развития у млекопитающих благодаря установлению зрительных связей с корой мозга.
Corpus geniculatum mediale служит промежуточной станцией для слуховых импульсов, направляющихся отсюда к коре височной области.
Corpus geniculatum med. лучше выражено у животных с более острым слухом; у летучей мыши, например, оно выражено сильнее, чем у человека. Corpus geniculatum lat. и corpus geniculatum med., развивающиеся из стенок второго мозгового пузыря (diencephalon), рассматриваются или как части зрительного бугра, или как особое образование - metathalamus.
На рис. 203, взятом мною из Геррика и несколько видоизмененном, доказаны ядра зрительных бугров и главнейшие тракты, связанные с этими ядрами.
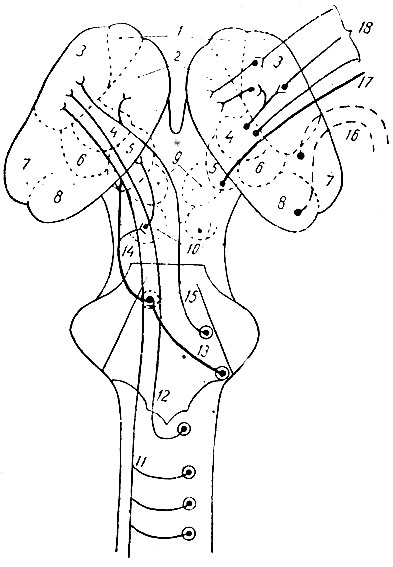
Рис. 203. Схема ядер зрительных бугров и важнейших трактов, связанных сними. На рисунке изображена верхняя поверхность стволовой части мозга после удаления мозговых полушарий и мозжечка (из Геррика; видоизменено). 1 - nucleus thalami anterior; 2 - nucleus thalami medialis; 3 - nucleus thalami lateralis; 4 - nucleus thalami ventralis; 5 - corpus geniculate mediale; 6 - nucleus thalami posterior; 7 - pulvinar; 8 - corpus geniculatum laterale; 9 - colliculus superior; 10 - colliculus inferior; 11 - lemniscus spinalis; 12 - lemniscus medialis; 13 - слуховое волокно, связующее nucleus cochlearis с oliva superior; 14 - lemniscus lateralis; 15 - lemniscus nervi trigemini; 16 - зрительная радиация; 17 - слуховая радиация; 18 - радиация общей чувствительности
Зрительный бугор связан с корой мозга волокнами, направляющимися или от бугра к коре (fibrae thalamo-corticales), или от коры к бугру (fibrae cortico-thalamicae). Все эти волокна, как центробежные, так и центростремительные, взятые в совокупности, носят название таламической радиации (radiatio thalamica). Их подразделяют на 4 группы: 1. Вентральный пучок, или нижняя ножка зрительного бугра, поперечно направляется под nucleus lenticularis от переднего вентрального отдела thalamus к коре височной доли и к insula Reilii (рис. 202). 2. Лобный пучок, или лобная ножка, состоит из волокон, идущих через переднее бедро внутренней капсулы от бокового ядра зрительного бугра к коре лобной области, а также и к nucleus caudatus (рис. 202 и 204). 3. Теменная ножка выходит из боковой поверхности thalamus и проходит вверх через заднее бедро внутренней капсулы. Она связывает боковое ядро thalamus с корой теменной доли и с задним отделом лобной области. Волокна этого пучка, оканчивающиеся в gyrus centralis posterior, несут импульсы: осязательные, тепловые, мышечные, суставные, сухожильные, а может быть также и болевые. Все эти волокна помещаются во внутренней капсуле позади кортико-спинального тракта (рис. 204). 4. Волокна затылочной ножки (зрительная радиация) начинаются от pulyinar и corpus geniculatum lat., проходят через самый задний отдел внутренней капсулы (pars retrolenticularis) и направляются по наружной стороне заднего рога бокового желудочка к коре затылочной доли, преимущественно в область fissurae calcarinae. (В этом же пучке содержится некоторое количество волокон, идущих от коры затылочной доли к передним буграм четверохолмия.)
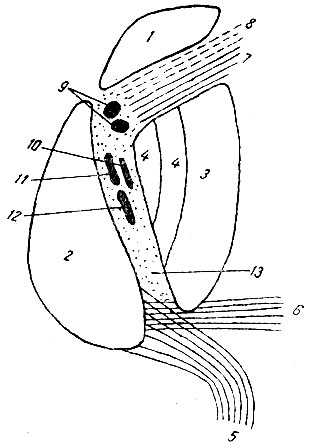
Рис. 204. Схема внутренней капсулы. 1 - nucleus caudatus; 2 - thalamus; 3 - putamen; 4 - globus pallidus; 5 - зрительная радиация; 6 - слуховая радиация; 7 - передняя таламическая радиация; 8 - лобно-мостовый тракт; 9 - tractus cortico-bulbaris; 10 - tractus cortico-rubralis; 11 - tractus cortico-spinalis (рука); 12 - tractus cortico-spinalis (нога); 13 - таламическая радиация (чувствующие волокна) (по Рансону)
В сублентикулярной части внутренней капсулы, расположенной вентрально к заднему концу nucleus lenticularis, находятся волокна слуховой радиации, идущие от corpus geniculatum mediale к височной поперечной дольке, расположенной в глубине сильвиевой борозды.
При использовании материалов сайта активная ссылка обязательна:
http://anfiz.ru/ 'Анатомия и физиология человека'