Глава XXVIII - 1932
Учение Павлова о системности в работе коры больших полушарий представляло новый важный этап в развитии физиологии и патологии высшей нервной деятельности. С первых шагов изучения физиологии больших полушарий Павлов шел путем анализа, путем дробления сложной и целостной корковой деятельности на составные, простые элементы. Синтетическая же, интегративная деятельность коры, согласно собственному признанию Ивана Петровича, оставалась как бы в тени, на втором плане. Это не значит, что она игнорировалась или была забыта: таков был избранный методологический путь. Но и до возникновения учения о системности, как о высшем синтезе - корковой интеграции, синтетический момент имел место в виде представления Павлова о коре как о "динамической функциональной системе". Это было ясно изложено в "Лекциях о работе больших полушарий головного мозга" 1927 г. Принцип системности - это принцип целостности в работе коры. Он являлся объективным выражением преодоления прежде господствовавшего аналитизма.
Учение о системности имеет свою внутреннюю историю. Оно начинается с наблюдений Кржышковского (1908 г.) над тем, что изменение обычного порядка раздражителей меняло ход опыта. Некоторый материал на эту тему мы находим в работе Сирятского с тоновой мозаикой (1923-1924 г.), когда правильное чередование положительного и тормозного тонов воспроизводилось на вновь пробуемых тонах вверх и вниз по тон-скале. Особенно рельефно самый факт системности выступил в опытах Купалова с кожной мозаикой (1924-1925 г.); им был введен стереотип с одинаковыми интервалами; эти исследования не только подтвердили на кожном анализаторе факты Сирятского, но и вскрыли взаимосвязь элементов мозаичной системы из положительных и тормозных очагов. Взаимная индукция выступала здесь, как частный случай взаимосвязи элементов системы вообще. Далее, эксперименты со стереотипной системой раздражителей (при одинаковых интервалах) были проведены д-ром Соловейчиком (1926 г.). Он обратил внимание на нарушения нормальной корковой деятельности под влиянием изменений обычного порядка условных раздражителей.
Мы уже говорили о том, что почти все сотрудники лаборатории И. П. Павлова в 1929 г. перешли на работу со стереотипом. Это способствовало быстрому накоплению нового фактического материала о системности.
Развернутое учение И. П. Павлова о системности стало возможным только после ряда специальных работ 1930-1932 гг. (Асратян, Скипин, Рикман, Выржиковский, Зевальд, Линдберг и др.).
Первое официальное выступление Павлова на эту тему было в 1930 г. (лекция врачам Института усовершенствования врачей "Физиология и патология высшей нервной деятельности").
Твердая, установившаяся система представлений по этому вопросу была изложена Иваном Петровичем в его докладе на Международном психологическом конгрессе в Копенгагене в 1932 г., где он говорил о "динамической стереотипии высшего отдела головного мозга".
Отнюдь не является случайным то обстоятельство, что Иван Петрович выступил с новым учением о системности именно на психологическом конгрессе. Он хотел показать что учение о высшей нервной деятельности преодолевает односторонности ассоциационизма и гештальтизма, как аналитического и синтетического направлений в психологии.
Почти на всех павловских "средах" этого времени (1930-1932 гг.) происходило обсуждение противоречий между ассоциативной и гештальтной психологией. Для Павлова были неприемлемы методологические установки гештальтистов, отвергавших, по существу, анализ. Мы можем с правом утверждать, что рождение понятия "системности" не только было обусловлено внутренним ходом развития учения Павлова, но и стояло в связи с методологическими спорами в области психологии. По этому поводу сам Иван Петрович говорил, что "учение о высшей нервной деятельности соединяет в себе ассоциационизм и гештальтизм"*.
* ("Среда" 30 сентября 1931 г. и др.)
"Динамическая стереотипия" обозначала в докладе Павлова (1) принцип системности. Этими словами он хотел подчеркнуть два взаимодействующих момента корковой деятельности: стереотипность и динамичность. Как животным, так и людям постоянно приходится устанавливать тот или иной жизненный стереотип временных связей и вместе с тем его менять и переделывать в зависимости от обстановки. "Система эта, - говорил Иван Петрович, - состоит из ряда положительных раздражителей разных рецепторов и различной интенсивности, а также и из отрицательных. Так как все эти раздражения оставляют после себя большие или меньшие следы, то точные постоянные эффекты раздражителей в системе могут получиться всего легче и скорее только при одних и тех же промежутках между раздражителями, притом же применяемых в строго определенном порядке, т. е. при внешнем стереотипе. В окончательном результате получается динамический стереотип, т. е. слаженная, уравновешенная система внутренних процессов. Образование, установка динамического стереотипа есть нервный труд чрезвычайно различной напряженности, смотря, конечно, по сложности системы раздражителей, с одной стороны, и по индивидуальности и состоянию животного, с другой" (стр. 623-624) (1). В процессе индивидуальной жизни стереотипы "наслаиваются" друг на друга. В основе системности функций коры лежит цепь нервных следов от прежних раздражений. Эта система следов обнаруживается и при отсутствии самих реальных раздражителей, ее образовавших, что доказывается опытами Асратяна, Скипина и др. Нарушения зафиксированного стереотипа нервных связей могут повести к хроническим нервным заболеваниям. Иван Петрович считал, что физиологические процессы, совершающиеся в коре больших полушарий при установке, переделке и прочих изменениях стереотипа, соответствуют тому, что субъективно мы называем чувствами положительного и отрицательного характера. Трудности ломки старого динамического стереотипа могут сопровождаться меланхолическими переживаниями. С этой точки зрения Иван Петрович провел физиологический анализ одного случая меланхолии, что несомненно представляет интерес для психиатрии.
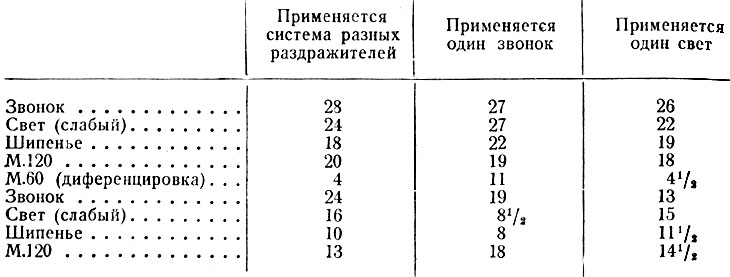
Опыты Э. А. Асратяна (2, 3) заключались в следующем: в течение более или менее длительного времени практиковалась одна и та же система условных пищевых раздражителей. Когда система разных раздражителей заменялась применением столько же раз одного раздражителя, то при действии последнего воспроизводилась старая система т. е. на месте сильных раздражителей получался большой эффект, на месте слабых маленький, а на месте тормоза - близкий к нулю. При. водим данные из работы Асратяна:
Опыты на 'АРЧ'

Опыты на 'МАСИС'
Из приведенных протоколов видно, что при замене системы разных раздражителей каким-либо одним сильным (звонок, метроном) или слабым (свет) воспроизводилась старая система отношений. Эффект зависел не только от наличного, повторяемого через те же интервалы раздражителя, но и от следов прежних раздражителей (сильных, слабых и тормозных). Старый стереотип сказывался и на секреторной и на двигательной реакциях. Так, последняя на месте прежней диферендировки была отрицательная и даже наступал отказ от еды.
С повторением нового стереотипа из одного раздражителя эффект постепенно становился одинаковым на всех местах, т. е. начинал соответствовать наличному раздражителю. Таким образом старая система разрушалась, и на ее разрушение уходило приблизительно столько же времени, сколько и на выработку. Далее было доказано, что системность воспроизводится не только при повторении какого-либо из раздражителей, входивших в старую систему (в данном случае, при повторении света, звонка, метронома 120), но и другими раздражителями, не принимавшими участия в образовании системы (например, кожно-механическое раздражение). В опытах с системностью имели значение порядок раздражителей и одинаковый интервал между ними. Но из специальных экспериментов с системностью на разных интервалах следовало, что первостепенная роль здесь принадлежит порядку раздражителей. Поэтому автор полагал, что условные рефлексы на время и явления системности не тождественны, хотя между ними и существует некоторая связь. Он считал, что в физиологической основе явлений системности лежат три закономерности: 1) взаимодействие различных частей функционального целого, 2) свойство сохранения нервных следов и 3) свойство фиксации, закрепления этих следов.
В параллельно проводившихся опытах Г. В. Скипина (4) упрощенная система разнородных раздражителей заменялась одним слабым раздражителем - светом. Также, как и в выше приведенных опытах, здесь выявлялась системность. При замене системы одним сильным раздражителем - стуком метронома - системность могла проявиться только после дальнейшего упрочения стереотипа разнородных раздражителей. При изменении в применяемом порядке условных раздражителей выработанная система разрушалась. В этом случае она восстанавливалась после 10-дневного перерыва. Старая угашенная система вновь восстанавливалась при развитии сонливости, т. е. иррадиированного торможения в коре. На этот факт необходимо обратить особое внимание и повторить такие опыты с целью детального анализа закономерностей снятия торможением поздних нервных следов и выявления более ранних и прочнее зафиксированных.
Из этих экспериментов вытекало, что выявить системность легче удается при помощи слабых раздражителей, так как слабое наличное раздражение менее, чем сильное, маскирует старые нервные следы. Из приведенных опытов также следовало, что при анализе зависимости величины условного рефлекса от силы условного раздражителя необходимо учитывать кроме физической силы раздражителя и всю совокупность нервных процессов, протекающих в этот момент в коре больших полушарий. Это надо считать, по нашему мнению, весьма существенным примечанием к "закону силы". На самом деле, испытание одного повторяемого раздражителя происходит "при прочих равных условиях" с испытанием системы разнородных раздражителей, т. е. при условии одинакового пищевого режима и примерно одинаковой пищевой возбудимости. Однако эффект зависит не столько от наличного раздражения одной и той же физической силы, сколько от конкретных отношений, сложившихся в предыдущей истории опыта.
В других своих опытах Скипин (5) у нескольких собак имел комплексный пищевой раздражитель из свистка с одновременным пассивным подниманием ноги. Один свисток вызывал активное поднятие ноги. Когда этот комплексный раздражитель был отменен, то все раздражители находившиеся на этом месте системы, стали осложняться поднятием ноги. Экспериментатор объяснял это явление, как доминанту: "наблюдаемая системность обусловлена работой других, ранее работавших элементов коры, приводимых в деятельное состояние применяемым нами раздражителем" (стр. 30-31) (5). Таким образом прочность условно-рефлекторных связей обусловлена инертностью, косностью выработанных рефлексов, а косность и инертность нервных процессов одно из основных функциональных свойств высшего отдела центральной нервной системы. Под влиянием изменяющейся внешней среды образованные условно-рефлекторные связи тормозятся, но не разрушаются вновь образующимися рефлексами.
В той же форме, как в работах Асратяна и Скипина, явления системности были установлены и в опытах В. И. Павловой (6).
Вопрос о взаимодействии сильных и слабых условных раздражителей в системе исследовался В. В. Рикманом (7). Он применял систему из двух раздражителей: звонка (сильного) и света (слабого). Оказалось, что свет один, без звонка, дает рефлекс в 15 делений шкалы, тот же свет при применении его в системе со звонком уменьшает свой эффект до 10-12. Надо полагать, что здесь сказался механизм отрицательной индукции от действия сильного раздражителя на слабый в пределах зафиксированной системы.
Опыты Л. О. Зевальда (8) были проверкой предыдущих исследований, но вместе с тем они внесли и кое-что новое. В опытах на двух собаках сначала не было стереотипа и применение одного раздражителя, естественно, какой-либо системности не обнаруживало. Когда же экспериментатор ввел систему (стереотип), тогда могла быть обнаружена и системность. Эта системность отчетливо устанавливалась уже после 9-10 опытов со стереотипом. Системность обнаруживалась при повторении как слабого, так и сильного раздражителя, но резче она выступала при повторении слабого. Так же, как и в предыдущих опытах, системность проявлялась на секреторной и двигательной реакциях. Новое состояло в следующем. Если при наличии системы наблюдались гипнотические фазы, то такие же фазы на тех же местах имелись и при замене системы одним раздражителем, при этом фазы выступали даже резче. У одной из собак было особое явление: при замене системы разных раздражителей одним раздражителем развилось гипнотическое состояние, и тогда старая система была воспроизведена в виде парадоксальной фазы, с соответствующим искажением слабых и сильных мест в системе опыта. У другой возбудимой собаки до введения системы силовые отношения между рефлексами были не вполне ясными, после же введения системы "закон силы" выступил резче. То же потом наблюдали Л. С. Григорович и О. П. Ярославцева.
К опытам по системности примыкали несколько отличные от них исследования Выржиковского и Усиевича. Опыты С. Н. Выржиковского (9) состояли в том, что среди системы раздражителей он применял один раздражитель четыре раза, сопровождая его пищевым подкреплением только в последний, четвертый раз, а первые три раза оставляя без подкрепления. Через некоторое время с трудом условный рефлекс выработался: на первые два-три раздражения был тормозной эффект, а на четвертое - положительный. Затем, проводилась переделка зафиксированных системных отношений: первые три раздражения стали подкрепляться, как и четвертое. Собака сильного типа с трудом, но все же решила эту задачу.
Аналогичные опыты проводил М. А. Усиевич (10). У собаки возбудимого типа он вырабатывал условный пищевой рефлекс, подкрепляемый через три применения в четвертое. Это представляло для данной нервной системы трудную задачу даже при наличии облегченных условий (применение в опыте только одного данного раздражителя). В процессе выработки этого рефлекса имели место "срывы", в силу "сшибки" возбуждения и торможения.
Углубленные исследования системных отношений продолжались и далее (Петрова, Линдберг, Михайлович, Адлерберг, Григорович, Ярославцева и др.). В дальнейшем явления системности были использованы для изучения силы и подвижности нервных систем различных типов (применение систем различной сложности, переделка систем и т. д.).
Одним из физиологических механизмов системности надо считать правильные, периодические колебания корковой возбудимости, обусловленные работой с одинаковыми интервалами. С этой стороны вопрос о системности связан с вопросом об условном рефлексе на время. Последнему была посвящена специальная работа Ф. Д. Василенко (11). Он проводил опыты с пищевыми рефлексами на собаке флегматического типа, отличавшейся большою точностью своей работы. Условные раздражители применялись через одинаковый интервал в 5 мин. Чтобы лучше проследить развитие рефлекса времени, давался один слабый раздражитель - свет. Условный рефлекс на свет постепенно рос, а потом снова снизился. Это значит, что сначала к рефлексу на свет прибавлялся образовавшийся рефлекс на время, а потом последний потерял свое действие благодаря постоянной отрицательной индукции от наличного раздражения. Таким образом, в начале получалась суммация рефлекса времени с рефлексом на свет. Выработанный условный рефлекс на время, уточняясь и совпадая с действием безусловного раздражителя, постепенно терял свое влияние на эффект слабого светового раздражителя в качестве компонента. Из этого следовало важное методическое заключение: при работе с условными рефлексами на равных промежутках рефлекс на время в окончательном итоге существенного значения не имеет, между тем как работа на равных промежутках дает возможность изучить влияние условных рефлексов друг на друга, т. е. взаимодействие элементов системы.
Уточнением вопроса об условном рефлексе на время занимался С. В. Клещ о в (12). Он производил небольшие изменения промежутков времени между раздражителями и определял их влияние на величину условного пищевого рефлекса. Он ставил опыты на трех собаках с обычным интервалом в 5 мин., с удлиненным до 5 мин. 15 сек. (или 5 мин. 20 сек.) и с укороченным до 4 мин. 45 сек. (или до 4 мин. 30 сек.). Изменение прочно усвоенного интервала на 15-30 сек. резко отражалось на величине условных рефлексов, причем удлинение интервала влияло значительно сильнее, чем укорочение. Так, при удлинении наблюдалось уменьшение эффекта на 15-24% при укорочении - увеличение эффекта на 5-14%. Объяснение было дано такое: при Удлинении интервала вводились два обстоятельства, неблагоприятных для условного рефлекса времени: отодвигание безусловного подкрепления для рефлекса времени и увеличение его работы за время удлиняющегося отставления; при укорочении, наоборот, сокращали эту работу и приближали пищевое подкрепление. Это и лежало в основе указанных изменений.
Из ряда предыдущих исследований и, в частности Купалова в Скипина (см. главу XXVII), вытекало предположение о закономерном изменении величины условных рефлексов в зависимости от интервалов между условными раздражителями. Проверка этого предположения составляла задачу опытов С. И. Гальперина (13). На собаке уравновешенного типа повторялся какой-либо один раздражитель (свет и шум из слабых, стук метронома и бульканье из сильных) на обычном интервале в 6 мин. при удлинении его до 10 мин. и укорочении до 4 мин. Сравнение результатов этих опытов дало следующее: 1) При удлинении интервала до 10 мин. было увеличение условных рефлексов на слабые раздражители и уменьшение на сильные, 2) при укорочении интервала до 4 мин. - обратный результат, 3) безусловные слюнные рефлексы при удлинении интервала повышались при слабых условие раздражителях и понижались при сильных, противоположные изменния были при укорочении интервала. Автор представил объяснение этих фактов с точки зрения учения Павлова о пределе работоспособности корковых клеток. "Повышенная возбудимость корковых клеток ведет к увеличению рефлексов на сильные условные раздражители" (стр. 253) (13). Однако, по нашему мнению, автор мог объяснить таким способом только одну, малую часть приведенных фактов, а именно: увеличение условных рефлексов на слабые раздражители, уменьшение на сильные при удлинении интервала до 10 мин., а все остальное является непонятным и даже противоречащим павловской теории работоспособности.
Следовательно, эта работа не завершала исследования вопроса о закономерностях изменения величины условных рефлексов в зависимости от интервалов между ними.
С каждым годом растущее многообразие опытов приносило подтверждение основного количественного "закона силы" и вместе с тем открывало ряд новых исключений к нему. Одно из нарушений правила о связи величины эффекта с силой раздражения было установлена И. Р. Пророковым (14). У собаки возбудимого типа не существовало правильного соотношения величин слабых и сильных условных рефлексов. Причина этого сказалась в слишком большом для данной нервной системы отставлении условных раздражителей (30-45 сек.). Когда было введено более короткое отставление, тогда получилась зависимость эффекта от силы раздражителя, согласно "закону силы". Значит, для возбудимой нервной системы одним из условий для различения условных раздражителей по силе является необходимость работать с коротко отставленными рефлексами, чтобы не создавать трудной нервной задачи в виде сшибки возбуждения и торможения.
В других экспериментах (Н. В. Виноградова) (15) наблюдалось замаскирование "закона силы" под влиянием индукции. У одной собаки сначала зависимость эффекта от силы раздражителя существовала, после же выработки диференцировки на частоту кожно-механическото раздражения произошло нарушение "закона силы", рефлекс на слабый, положительный кожно-механический раздражитель сильно увеличился и стал равен сильным рефлексам. Такая же история произошла и со слабым прерывистым светом после выработки к нему диференцировки на свет другой частоты миганий. В том и другом случае имела место хроническая положительная индукция, увеличивавшая эффект слабых раздражителей до уровня сильных. При снижении пищевой возбудимости удавалось получать нормальные отношения соответственно с "законом силы". Интересно, что в случае кожно-механического раздражителя условный рефлекс на него оставался увеличенным на протяжении нескольких месяцев и после прекращения употребления его диференцировки. "Закон силы" был обоснован преимущество опытами с пищевыми рефлексами. Но существует ли зависимость эффекта от силы раздражителей при кислотных условных рефлексах? Этот вопрос разрешал В. К. Федоров (16). У двух его собак отсутствовала зависимость величины кислотных условных рефлексов от физической силы раздражителей, однако такая зависимость была у тех же животных на пищевых условных рефлексах. Эту зависимость можно было выявить и на кислотных условных рефлексах, когда были применены более слабые концентрации раствора соляной кислоты: вместо 0 15-0,20% (как обычно применяли и другие исследователи до этого) был взят раствор 0,05%. Таким образом, на поставленный вопрос был получен положительный ответ. Закон зависимости величины условных рефлексов от силы раздражителей при нормальной, т. е. средней степени возбуждения подкоркового центра, относится к кислотным условным рефлексам в такой же мере, как и к пищевым. Разница сводится лишь к тому, что эту среднюю степень возбуждения легко поддерживать для пищевого центра и трудно для кислотного в силу их биологических особенностей.
Следовательно, зависимость условно-рефлекторного эффекта от силы раздражителя можно считать универсальной закономерностью корковой деятельности.
В 1912 г. Ерофеева проводила опыты по превращению электрического раздражения кожи в условный пищевой раздражитель (см. главу X). Она получила удивительный факт: слабый электрический ток, подкреплявшийся едой, осложнился сонливостью, после чего и постепенно усиливавшиеся токи (до больших интенсивностей) продолжали сопровождаться сном. Эти опыты в новой вариации были возобновлены Н. А. Подкопаевым (17). У двух животных сначала были выработаны условные пищевые рефлексы на электрическое раздражение кожи фарадическим током, начиная со слабого тока с постепенным увеличением его силы. Потом была образована диференцировка на электрическое раздражение другого места кожи. У одной собаки диференцировка выработалась полностью, у другой - только на 50%, а затем стала сопровождаться оборонительной реакцией. На обеих собаках наблюдалось осложнение слабого электрического раздражителя торможением, как это было и в опытах В. К. Федорова (см. главу XXVII), которое не пропадало и при усилении тока. Условный слюнной рефлекс на ток уменьшался при повторении из опыта в опыт и продолжал уменьшаться при постепенном усилении тока. Автор считал, что слабый электрокожный раздражитель (в качестве условного пищевого) обладает свойствами вообще слабого кожного раздражителя, а именно быстрым Переходом в торможение, вызывающим общую сонливость. При очень постепенном увеличении силы тока он сохранял указанные свойства, несмотря на достижение очень большой физической силы.
Итак, слабый электрический ток, примененный в качестве условного пищевого раздражителя, осложняется торможением, ток продолжает осложняться торможением и тогда, когда постепенно доведен до очень большой силы. Эти факты подчеркивают не "размежевание, разграничение возбуждения и торможения", а их единство, тесную внутреннюю взаимосвязь. К этому времени и И. П. Павлов не был чужд такого взгляда. Так, по поводу опытов А. А. Линдберга, в которых Условный тормоз превратился в условный положительный рефлекс И порядка, и по поводу опытов Г. В. Скипина, в которых после сшибки пропал условный тормоз и превратился в условный рефлекс II порядка, представляла задачу опытов С. И. Гальперина (13). На собаке уравновешенного типа повторялся какой-либо один раздражитель (свет и шум из слабых, стук метронома и бульканье из сильных) на обычном интервале в 6 мин. при удлинении его до 10 мин. и укорочении до 4 мин. Сравнение результатов этих опытов дало следующее: 1) при удлинении интервала до 10 мин. было увеличение условных рефлексов на слабые раздражители и уменьшение на сильные, 2) при укорочении интервала до 4 мин. - обратный результат, 3) безусловные слюнные рефлексы при удлинении интервала повышались при слабых условных раздражителях и понижались при сильных, противоположные изменения были при укорочении интервала. Автор представил объяснение этих фактов с точки зрения учения Павлова о пределе работоспособности корковых клеток. "Повышенная возбудимость корковых клеток ведет к увеличению рефлексов на сильные условные раздражители" (стр. 253) (13). Однако, по нашему мнению, автор мот объяснить таким способом только одну, малую часть приведенных фактов, а именно: увеличение условных рефлексов на слабые раздражители, уменьшение на сильные при удлинении интервала до 10 мин., а все остальное является непонятным и даже противоречащим павловской теории работоспособности.
Следовательно, эта работа не завершала исследования вопроса о закономерностях изменения величины условных рефлексов в зависимости от интервалов между ними.
С каждым годом растущее многообразие опытов приносило подтверждение основного количественного "закона силы" и вместе с тем открывало ряд новых исключений к нему. Одно из нарушений правила о связи величины эффекта с силой раздражения было установлено И. Р. Пророковым (14). У собаки возбудимого типа не существовало правильного соотношения величин слабых и сильных условных рефлексов. Причина этого сказалась в слишком большом для данной нервной системы отставлении условных раздражителей (30-45 сек.). Когда было введено более короткое отставление, тогда получилась зависимость эффекта от силы раздражителя, согласно "закону силы". Значит, для возбудимой нервной системы одним из условий для различения условных раздражителей по силе является необходимость работать с коротко отставленными рефлексами, чтобы не создавать трудной нервной задачи в виде сшибки возбуждения и торможения.
В других экспериментах (Н. В. Виноградова) (15) наблюдалось замаскирование "закона силы" под влиянием индукции. У одной собаки сначала зависимость эффекта от силы раздражителя существовала, после же выработки диференцировки на частоту кожно-механического раздражения произошло нарушение "закона силы", рефлекс на слабый, положительный кожно-механический раздражитель сильно увеличился и стал равен сильным рефлексам. Такая же история произошла и со слабым прерывистым светом после выработки к нему диференцировки на свет другой частоты миганий. В том и другом случае имела место хроническая положительная индукция, увеличивавшая эффект слабых раздражителей до уровня сильных. При снижении пищевой возбудимости удавалось получать нормальные отношения соответственно с "законом силы". Интересно, что в случае кожно-механическою раздражителя условный рефлекс на него оставался увеличенным на протяжении нескольких месяцев и после прекращения употребления его диференцировки. "Закон силы" был обоснован преимущественно опытами с пищевыми рефлексами. Но существует ли зависимость эффекта от силы раздражителей при кислотных условных рефлексах. Этот вопрос разрешал В. К. Федоров (16). У двух его собак отсутствовала зависимость величины кислотных условных рефлексов от физической силы раздражителей, однако такая зависимость была у тех же животных на пищевых условных рефлексах. Эту зависимость можно было выявить и на кислотных условных рефлексах, когда были применены более слабые концентрации раствора соляной кислоты: вместе 0,15-0,20% (как обычно применяли и другие исследователи до этого) был взят раствор 0,05%. Таким образом, на поставленный вопрос был получен положительный ответ. Закон зависимости величины условны: рефлексов от силы раздражителей при нормальной, т. е. средней степени возбуждения подкоркового центра, относится к кислотным условным рефлексам в такой же мере, как и к пищевым. Разница сводится лишь к тому, что эту среднюю степень возбуждения легко поддерживать для пищевого центра и трудно для кислотного в силу их биологических особенностей.
Следовательно, зависимость условно-рефлекторного эффекта от силы раздражителя можно считать универсальной закономерностью корковой деятельности.
В 1912 г. Ерофеева проводила опыты по превращению электрического раздражения кожи в условный пищевой раздражитель (см. главу X). Она получила удивительный факт: слабый электрический ток, подкреплявшийся едой, осложнился сонливостью, после чего и постепенно усиливавшиеся токи (до больших интенсивностей) продолжали сопровождаться сном. Эти опыты в новой вариации были возобновлены Н. А. Подкопаевым (17). У двух животных сначала были выработаны условные пищевые рефлексы на электрическое раздражение кожи фарадическим током, начиная со слабого тока с постепенным увеличением его силы. Потом была образована диференцировка на электрическое раздражение другого места кожи. У одной собаки диференцировка выработалась полностью, у другой - только на 50%, а затем стала сопровождаться оборонительной реакцией. На обеих собаках наблюдалось осложнение слабого электрического раздражителя торможением, как это было и в опытах В. К. Федорова (см. главу XXVII), которое не пропадало и при усилении тока. Условный слюнной рефлекс на ток уменьшался при повторении из опыта в опыт и продолжал уменьшаться при постепенном усилении тока. Автор считал, что слабый электрокожный раздражитель (в качестве условного пищевого) обладает свойствами вообще слабого кожного раздражителя, а именно быстрым переходом в торможение, вызывающим общую сонливость. При очень постепенном увеличении силы тока он сохранял указанные свойства, несмотря на достижение очень большой физической силы.
Итак, слабый электрический ток, примененный в качестве условного пищевого раздражителя, осложняется торможением, ток продолжает осложняться торможением и тогда, когда постепенно доведен до очень большой силы. Эти факты подчеркивают не "размежевание, разграничение возбуждения и торможения", а их единство, тесную внутреннюю взаимосвязь. К этому времени и И. П. Павлов не был чужд такого взгляда. Так, по поводу опытов А. А. Линдберга, в которых Условный тормоз превратился в условный положительный рефлекс И порядка, и по поводу опытов Г. В. Скипина, в которых после сшибки Пропал условный тормоз и превратился в условный рефлекс II порядка, Иван Петрович высказал мысль, что "все раздражители имеют двойное действие"*, что "у каждого условного раздражителя надо предполагать две функции: возбуждение и торможение"**.
* ("Среда", 17 февраля 1932 г.)
** ("Среда", 24 февраля 1932 г.)
Однако на ряду с этими утверждениями продолжало господствовать стремление рассматривать возбуждение и торможение в их раздельности, "размежевании". Свое крайнее выражение эта концепция получила в работах Григорович и Ярославцевой, изучавших вопрос о так называемых "нейтральных пунктах" коры. Еще Фурсиков (см. главу XVIII) и Сирятский (см. главу XIX) указывали на возможность получить пространственное разграничение корковых полей возбуждения и торможения. Л. С. Григорович (18) делала опыты на двух собаках. У одной из них тон 600 (колебаний в сек.) был условным пищевым раздражителем, тон 300 - диференцировочным, тормозным а тон 450 испытывался, как "нейтральный". У другой собаки работа велась на кожном анализаторе: на одном месте кожи была положительная (пищевая) касалка, на другом - диференцировочная, третья касалка была прикреплена ровно на середине между ними и испытывалась, как "нейтральная". Объективными критериями нейтральности тона 450 у первой собаки служили 1) отсутствие пищевой реакции и 2) отсутствие особой двигательной реакции, которая обычно сопровождала диференцировку. Критериями нейтральности средней касалки у второй собаки были 1) отсутствие пищевой реакции и 2) отсутствие последовательного торможения и положительной индукции, что могло бы свидетельствовать о ее тормозном значении. Испытания "нейтральных" раздражителей делались очень редко (через несколько месяцев) и никогда не сопровождались едой. Исследователь считал доказанным существование корковых "нейтральных пунктов", однако отмечал "некоторое непостоянство в действии такого пункта" (стр. 249) и то, что "нейтральное поле есть подвижный пункт, который передвигается то в сторону тормозного поля, то в сторону поля раздражения", (стр.251) (18).
О. П. Ярославцева (19) исследовала "разграничение районов возбуждения и торможения" в звуковом анализаторе. Тон 1500 (колебаний в сек.) был условным пищевым раздражителем, тон 2200 - диференцировочным, тормозным, тон 1760 пробовался, как "нейтральный". Критерии нейтральности были таковы: 1) отсутствие положительной реакции и 2) отсутствие тормозного последействия и положительной индукции. Экспериментатор пришел к такому заключению: "Между районами возбуждения и торможения находится пункт, в котором нельзя обнаружить ни того, ни другого состояния. Этот пункт передвигается то в район раздражения, то в район торможения в зависимости от того, какое из этих двух состояний преобладает в период опытов" (стр. 266) (19).
У нас нет оснований сомневаться в правильности изложенных фактов. Можно допускать пространственное разграничение корковых полей возбуждения и торможения. Но действительно ли существуют между этими функциональными полями "нейтральные пункты", своего рода демаркационные полосы, отделяющие возбуждение от торможения? В интерпретации фактов содержится момент отрыва возбуждения от торможения.

И. П. Павлов на заседании в психиатрической клинике. 1932 г.

И. П. Павлов на заседании в психиатрической клинике. 1932 г.
Ввиду принципиального значения этих исследований необходимо их повторить и провести еще более тщательный экспериментальный анализ указанных явлений, исходя из другой методологической позиции - единства и взаимодействия возбуждения и торможения.
Переходим к следующему вопросу. В развитии учения о высшей нервной деятельности важное место принадлежит способу коротких условных раздражений. Впервые этот способ был применен Л. А. Орбели (1908 г.), затем В. М. Добровольским (1911 г.) и М. К. Петровой (1916 г.). До сего времени способом коротких раздражений пользовались для борьбы со сном у экспериментальных животных. П. С. Куполов применил этот способ в целях более глубокого проникновения в динамику нервных процессов, лежащих в основе условного рефлекса. В связи с этим соответственным образом был модифицирован и самый способ.
В опытах П. С. Купалова и Б. Н. Лукова (20) у двух собак после образования отставления условных пищевых рефлексов на 30 сек. условные раздражители изредка применялись в течение короткого времени: 3-5-10 сек.; после чего раздражитель прекращался, и следовала пауза в 27-25-20 сек. до подкрепления. Короткое применение раздражителя дало в среднем такую же или немногим меньшую условную секреторную реакцию, как и сплошное применение в течение 30 сек. Ход секреции в основном был одинаков в обоих случаях. При удлинении паузы после 3-секундного раздражения до 87 сек. величина условной секреции у возбудимого типа оказалась лишь немногим меньше величины рефлекса при сплошном применении раздражителя в течение 90 сек., ход секреции был одинаков. Другая картина получилась у собак сангвинического типа: короткое раздражение с паузой дало вдвое меньший эффект, чем при непрерывном раздражении. Следовательно, короткий способ раздражений мог бы быть способом для определения особенностей типов нервной системы. При применении короткого способа продолжала сохраняться зависимость эффекта от силы раздражителя. Авторы полагали, что "корковая часть дуги условного рефлекса должна состоять из нескольких различно функционирующих отделов. Первым отделом можно считать комплекс первичных рецепторных клеток. Следующий отдел, вероятно, представляет собой сложную систему невронов, обладающих способностью к длительному Функционированию под влиянием короткого начального раздражения".
Далее проводились опыты (Купалов) (21) по угашению условного рефлекса при длинном и коротком применении раздражителя. Угасание и восстановление рефлекса на свет (у двух собак) протекали в основном одинаково как при коротком раздражении в течение 3 сек., так и при обычном, сплошном применении света в течение 30 сек. У собаки возбудимого типа действие раздражителя в течение 3 мин. дало такую же по величине условную секрецию, как и раздражение в течение 3 сек. с последующей паузой в 2 мин. 57 сек. Условная секреция, вызванная коротким раздражением с последующей паузой, могла быть в несколько секунд нацело заторможена применением тормозного (диференцировочного) раздражителя. П. С. Купалов и Н. Н. Павлов (22) исследовали действие короткого условного раздражения при запаздывающем условном рефлексе. При запаздывающем на 2 мин. рефлексе можно было прекратить условное раздражение через 1 мин., и тогда в следующую минуту условное слюноотделение протекало так же, как и при сплошном применении раздражителя в продолжении 2 мин. Таким образом, внутренняя смена процессов и в этом случае продола происходить так же, как она происходила раньше. В связи с этим авторы полагали, что "запаздывающие условные рефлексы по своему механизму существенно отличаются от следовых условных рефлексов". Более короткое применение условного раздражителя, в течение 5-10 сек., в случае запаздывающего рефлекса оказывалось недействительным и не вызывало секреторной реакции. Короткое применение условного раздражителя при запаздывающих рефлексах вело к положительному эффекту лишь тогда, когда действие раздражителя прекращалось после того, как тормозная фаза сменялась выраженной фазой возбуждения.
Линия исследований с короткими раздражениями была продолжена и далее.
П. С. Купаловым и П. К. Денисовым (23) была выполнена работа установившая ясную зависимость тонуса коры больших полушарий от факторов внешней среды. Опыты производились в затемненной и освещенной камере. Оказалось, что свет в затемненной камере является для собаки более сильным условным раздражителем, нежели в освещенной. Звуковые рефлексы в освещенной камере давали более высокие цифры, чем в затемненной. Причину повышения условных пищевых рефлексов на свет в затемненной камере следует искать в явлениях адаптации глаза. Повышение же рефлексов на звуковые раздражители является результатом тонизирующего влияния на кору зрительных раздражений от окружающей среды. Таким образом "наличие определенного количества падающих на животное раздражителей является одним из факторов повышения тонуса коры больших полушарий и одним из условий бодрого состояния животного".
Давно оставался неразрешенным вопрос о взаимном растормаживании разных видов внутреннего торможения. Одни исследователи: Никифоровский (1910 г.), Потехин (1911 г.), Красногорский (1911 г.), Дегтярева (1914 г.) описали факты растормаживания одного внутреннего торможения другим. Другие исследователи: Лепорский (1911 г.), Горн, Чеботарева (1912 г.) - наоборот, установили, что то или иное соединение двух видов внутреннего торможения дает суммацию торможения, а растормаживания не дает. Однако следует заметить, что и у второй группы сотрудников были отдельные факты растормаживания, которым не придавалось значения. Эта тема получила свое разъяснение в работе А. Г. Иванова-Смоленского (24). Было испытано последовательное взаимодействие всех видов внутреннего торможения: 1) влияние диференцировки на диференцировку, 2) действие условного тормоза на диференцировку и наоборот, 3) влияние угасательного торможения на условное торможение и диференцировочное, 4) действие запаздывательного торможения на условный тормоз и диференцировку. Один вид внутреннего торможения применялся после другого через некоторый интервал. Одинаковые результаты были получены на трех собаках: все виды внутреннего торможения растормаживали друг друга. Эффект растормаживания получался на интервале 5-7 мин. и не получался на интервале в 1 мин. Растормаживание пропадало в случае многократного повторения указанных испытаний, очевидно, в силу развития концентрации торможения при тренировке. Таким образом изложенные выше противоречия в фактах могли зависеть от разницы в интервалах у разных исследователей. Следовательно, надо было считать, что при последовательном взаимодействии видов внутреннего торможения возможны как суммация торможения, так и растормаживание в зависимости от конкретных временных, силовых и прочих отношений.
Однако Павлов продолжал считать, что при одновременном или последовательном суммировании двух видов внутреннего торможения должна получаться суммация торможения, а не растормаживание. Бодрое оставался дискуссионным. Опыты в этом направлении продолжались (Ярославцева, Долин и др.).
"Внутреннее торможение, - писал Иванов-Смоленский, - подобно торможению внешнему имеет действительный эффект, не только тормозя, но и растормаживая". Надо заметить, что складывавшееся представление о растормаживании в этом случае (т. е. при взаимодействии видов внутреннего торможения) не было воскрешением старой, похороненной концепции растормаживания, как "торможения торможения" (по типу внешнего тормоза).
В 1932 г. продолжались дальнейшие исследования типов нервной системы. Иванов-Смоленский (25) описал сильный тип собаки с резко выраженным пассивно-оборонительным рефлексом. Ряд испытаний свидетельствовал о силе нервной системы этой собаки: быстрое образование условных рефлексов, достаточно сильное внутреннее торможение при "угашении с подкреплением", сплошном и прерывистом угашении, выработка трудной диференцировки на частоту касалки при наличии метрономной диференцировки, выработка запаздывания на 3 мин., разрушение диференцировки и производство сшибок на кожно-механических раздражителях без отклонений от нормы. Вместе с этим животное отличалось легкой внешней тормозимостью и пассивно-оборонительным поведением в экспериментальных условиях. Автор предложил два возможных объяснения: или мы имеем особую конституциональную комбинацию сильного типа с резко выраженным пассивно-оборонительным рефлексом или же такое поведение есть результат соответствующего воспитания, по примеру "заключенных" собак Выржиковского - Майорова? Мы должны признать правильным второе объяснение.
Кроме этого, на данной собаке производилось изучение иррадиации тормозного процесса и индукции. К диференцировке М. 50 (положительный М. 100) присоединялся положительный раздражитель - слабый свет - во время действия диференцировки и после ее прекращения. Иррадиация торможения из слухового анализатора в зрительный имела место и при испытаниях прочной диференцировки, но в течение 10-15 сек. иррадиация переходила в концентрацию, что выражалось в яркой положительной индукции на эффекте светового раздражителя.
На той же собаке сильного уравновешенного типа делались опыты (26) с условным запаздывающим рефлексом. Этот рефлекс (на Щи) был выработан среди других обычно отставленных условных рефлексов сразу как запаздывающий на 3 мин., без постепенного его отставления. Применения запаздывающего рефлекса один-два раза за опыт и при том не ежедневно благоприятствовали его выработке. В последействии запаздывающего рефлекса наблюдались гипнотические фазы (парадоксальная и уравнительная). Эти фазы были только в день применения запаздывающего рефлекса. Вместе с окончательной выработкой запаздывания и ослаблением последовательного торможения исчезли и фазовые изменения.
Новая характеристика слабого типа получила свое подтверждение в опытах Г. В. Скипина (27). В полном соответствии с данными Н. В. Виноградова (см. главу XXVII) было доказано, что слабый тип отличается слабостью обоих нервных процессов: и возбуждения и торможения. У исследуемой собаки слабого типа имелась ярко выраженная инертность в течении нервных процессов, что обнаруживалось в легком воспроизведении системности. У собаки была заметна внешняя тормозимость и слабо были выражены процессы выработанного торможения.
Автор описал интересный случай двигательной стереотипии у этой собаки. Когда она вспрыгивала на станок, то всегда поворачивалась задом к кормушке. Это происходило потому, что в течение двух лет у предыдущего работника в другой камере собаке надо было после прыжка делать стереотипный поворот к кормушке именно в эту сторону. Эта двигательная реакция продолжала осуществляться и в новой камере при обратном расположении кормушки.
Данная И. П. Павловым новая характеристика возбудимого (безудержного) типа нервной системы получила свое подтверждение в опытах И. Р. Пророкова (28). У подопытной собаки возбудимого типа были сильны оба процесса - и возбуждение и торможение, но в силу перевеса первого последний в значительной степени маскировался, и ни один вид внутреннего торможения не достигал абсолютности. Доказательства силы возбуждения: 1) собака сразу стала брать еду в изолированной камере и быстро выработала условные рефлексы, 2) быстро выработала условный рефлекс на сверхсильный раздражитель (звук электрического колокола), давала на него рефлекс, равный сильным и выше их, 3) в течение двух месяцев выдержала применение одного сверхсильного раздражителя и трудной диференцировки к нему (на прерывистый звук того же колокола) без отклонений от нормы, 4) вынесла длительное применение больших доз брома, по 3 г ежедневно (114 грм. на протяжении 11/2 мес.) и дала при этом повышение условных рефлексов, 5) угасание (прерывистое и непрерывное) было длительным, и 6) одно- и двух-дневное голодание не изменило величины условных рефлексов, что указывало на способность нервной системы выносить сильное возбуждение без развития запредельного торможения. Доказательства силы тормозного процесса: 1) способность нервной системы выработать диференцировочное торможение на 64% (на тормозной M. 88 к положительному М. 176), 2) выработалась диференцировка на сверхсильный колокол на 50-60% и 3) удалась выработка условного тормоза на прерывистый свет к сверхсильному колоколу на 80%. Таким образом возбудимый тип не есть "специалист возбуждения", у него тормозный процесс тоже сильный и может легко поддаваться тренировке.
В другом виде исследование на сильном, возбудимом типе было проведено В. В. Яковлевой (29). Она продолжила разработку явлений суммации условных рефлексов на возбудимой собаке. Производилось суммирование условных пищевых раздражителей: слабого со слабым (свет слабый плюс касалка), слабого со средним (свет слабый плюс бульканье) и слабого с сильным (свет слабый плюс стук метронома). Эти опыты подтвердили результаты работы Рикмана с суммацией. Суммирование раздражителей доводило их эффект до предельной величины. Повторение суммированных раздражителей непременно окончательном результате" обусловливало торможение всей коры, снижение условных рефлексов и развитие гипнотического состояния. Для экспериментатора остался неясным вопрос: каков механизм развития торможения, при повторении суммации? Обычно здесь принято думать о механизме запредельного торможения, но так ли это? (тем более, если речь идет о суммации двух слабых или слабого со средним). Автор указал на то, что развитию гипнотического состояния предшествовала фаза возбуждения, как "выражение борьбы между раздражением и торможением, между бодрым состоянием и наступающей сонливостью" (стр. 287) (29).
Как показали дальнейшие работы, развитие гипнотического торможения при суммации не является обязательным.
Одним из способов испытания типа нервной системы, силы его тормозного процесса были опыты с перенапряжением торможения. Предел его напряженности определялся путем удлинения времени действия диференцировки. Такие опыты делались Яковлевой и Подкопаевым. В. В. Яковлева (30) у собаки-сангвиника в стереотипной системе условных пищевых рефлексов производила удлинение диференцировки (на М. 50 к положительному М. 100) с обычных 30 сек. до 9 мин. Вначале удалось постепенно дойти до удлинения в 5 мин., что в данный период было пределом напряженности торможения, за которым произошел срыв в сторону возбуждения. Это сменилось гипнотическим состоянием, которое лечилось бромом (3,0 NaBr ежедневно). Когда животное пришло в нормальное состояние, было предпринято второе удлинение и доведено только до 4 мин., что опять вызвало срыв в сторону возбуждения, сменившегося гипнотическим состоянием. Несмотря на это экспериментатор продолжал удлинять диференцировку: при 5 мин. гипнотическое состояние исчезло и далее постепенно действие тормозного раздражителя было продлено до 9 мин. На восемнадцатый день применения 9-минутной диференцировки получился третий срыв, но, в отличие от предыдущих, сразу в сторону торможения. На этот раз применение тех же доз брома не помогло; быстрее, чем в первый раз, наступили явления бромного отравления. Длительный летний отдых и более высокая пищевая возбудимость способствовали восстановлению работоспособности, и задача на предел напряжения тормозного процесса (сразу после перерыва удлинение диференцировки до 9 мин.) была решена. Таким образом под влиянием тренировки сила торможения могла быть значительно увеличена.
Н. А. Подкопаев (31) у собаки умеренно возбудимого типа Удлинял действие диференцировочного раздражителя (М. 60) до 5 мин. 40 сек.; т. е. в 17 раз дольше обычного, не вызывая в коре никаких патологических отклонений. На фоне длительного действия диференцировки применялся какой-либо положительный раздражитель. По мере продления диференцировки наблюдались три стадии: сначала иррадиация торможения, потом концентрация и снова иррадиация, соответственным образом сказывавшиеся на эффекте положительного раздражителя.
Уже предыдущие исследования показали, что сшибки, вызывающие столкновение противоположных нервных процессов, не являются чем-то исключительно трудным для нервной системы. Некоторые типы нервной системы выносят эти сшибки без срывов или сшибки вызывают у них лишь временное нарушение обоих процессов, что сравнительно легко сравнивается. Так было и в опытах О. П. Ярославцевой (32) на собаке с нервной системой средней силы. У нее делались многократные сшибки в звуковом анализаторе: после диференцировочного тона сразу применялся положительный тон. Всего было произведено 39 сшибок с одинаковым результатом. Сшибки отражались только на тоновом и Метрономном "пунктах", не затрагивая остальных рефлексов. Только в более острые периоды иногда растормаживались все три диференцировки (тоновая, метрономная и кожно-механическая).
Перейдем теперь к рассмотрению отдельной работы, касавшейся ранее упоминавшегося нами вопроса об антагонизме секреторного и двигательного компонентов условного пищевого рефлекса. С. В. Клещов (33) наблюдал случай утрированной пищевой двигательной реакции при выработке диференцировки. Процесс выработки диференцировки на М. 88 (к положительному М. 176) сопровождался тем, что собака вылизывала пустую чашку в течение 20 сек. действия тормозного раздражителя. При выработке более тонкой диференцировки на М. 132 удалось получить такую реакцию один раз. При остром прерывистом угашении рефлекса на М. 176 наблюдалась фаза вылизывания пустой кормушки. Эту утрированную пищевую реакцию автор рассматривал как выражение антагонизма секреторного и двигательного компонентов условной пищевой реакции. При выработке диференцировки и при угашении создавались условия для индукционного возбуждения двигательного компонента. С дальнейшим углублением торможения оно захватывало и двигательный компонент, что вело к исчезновению описанной реакции.
Подобные явления в разных вариациях наблюдали и другие исследователи.
Переходим к рассмотрению новых материалов по патологии высшей нервной деятельности. В. В. Рикман (34) наблюдал интересный случай аналогии "травматического невроза" у собаки. Он продолжал опыты Конради (см. главу XXV) на той же собаке. Когда-то у нее применялся электрический ток в качестве безусловного подкрепления. Много времени спустя, у нее наблюдалась оборонительная реакция в экспериментальной обстановке. Обнаружение давних следов этой оборонительной реакции происходило при определенных условиях, а именно - при развитии тормозного, гипнотического состояния. Был проведен тщательный экспериментальный анализ данного факта. Делались опыты с повышением и понижением пищевой возбудимости, опыты в разное время дня, опыты с применением условных пищевых раздражителей и без них, одно стояние собаки в станке. Применение кофеина - 7-8 см3 1% раствора чистого кофеина устраняло оборонительный комплекс. При объяснении физиологического механизма явления экспериментатор не исключал роли иррадиации возбуждения из пищевого центра в оборонительный в первый период работы, когда пищевые реакции осложнялись оборонительной. В дальнейших стадиях развития явления также не исключалась роль взаимной индукции в подкорке. Но при всех этих обстоятельствах главное значение принадлежало развитию гипнотического торможения в коре. Как при, так называемом, "военном или командном неврозе" он обнаруживается во время сна, так и у собаки Рикмана оборонительный комплекс выявлялся при развитии гипнотического состояния и исчезал при устранении этого состояния.
Поэтому данный экспериментальный факт был описан, как "аналог травматического невроза".
Одним из приемов получения патологических отклонений высшей нервной деятельности была кастрация. Большую серию разнообразных исследований на 8 самцах-кастратах сделала М. К. Петрова (35)1. Собаки были разного типа нервной системы и разной степени изношенности. Петрова провела ряд испытаний до кастрации и после нее. Таким образом можно было установить влияние кастрации на высшую нервную деятельность. Все собаки в окончательном результате более или менее слабели после кастрации: у них страдали оба нервных процесса, особенно тормозный. Результат кастрации у разных типов был различный; кроме того, здесь имели значение возраст и степень изношенности нервной системы. Сильные, молодые животные возбудимого и уравновешенного типов после временной слабости восстанавливались, но только для обычных условий эксперимента. Слабые, тормозимые непосредственно после кастрации становились более работоспособными и бодрыми, но лишь временно. Такое временное улучшение высшей нервной деятельности у слабых было непродолжительным: у одной собаки - один месяц, у другой - четыре, а потом развивалась "инвалидность" высшей нервной деятельности. Почти у всех кастратов наблюдалась наклонность к гипнотическим состояниям. У всех кастратов так же имела место периодичность, циркулярность корковой деятельности, т. е. периоды относительно высоких рефлексов чередовались с периодами низких рефлексов. Бром и отдых возвращали животных к норме (до кастрации). Отмечалось различное отношение к дозировке брома до и после кастрации; после кастрации дозы снижались.*
* (Ряд ценных работ на эту же тему был выполнен проф. В. М. Архангельским и его сотрудниками в Днепропетровске.)
Далее, было исследовано изменение тонуса коры больших полушарий у кастратов под влиянием повышения пищевой возбудимости (36). Были повторены и подтверждены соответствующие опыты Рикмана на нормальных собаках (см. главу XXVII). Вместе с тем были получены и новые данные. Высшая нервная деятельность у кастратов при голодании резко отличается от таковой у сильных нормальных собак. Повышение пищевой возбудимости у кастратов сказывается в падении или полном исчезновении условных рефлексов, особому торможению подвергаются слабые рефлексы, при этом растормаживаются диференцировки. Степень этих нарушений зависит от силы нервной системы и от степени повышения пищевой возбудимости. Слабые кастраты реагируют на повышение пищевой возбудимости резче, чем сильные. Голодание можно считать физиологическим индикатором силы нервной системы. Под влиянием голодания исчезла ультрапарадоксальная фаза, бывшая у одной собаки, то же было и при понижении пищевой возбудимости.
Производились также опыты по перенапряжению тормозного процесса у кастратов различного типа нервной системы (37). В системе условных рефлексов делалось удлинение метрономной диференцировки с обычных 30 сек. до 5 мин. У кастратов слабого типа в результате удлинения диференцировки совершенно срывалась их нормальная деятельность в сторону резкого и длительного снижения коркового тонуса. У других собак после продления диференцировки отмечалось повышение условных рефлексов. У большинства кастратов не произошло растормаживания диференцировки при ее удлинении и только у двух она растормозилась. Для экспериментатора остался непонятным вопрос: почему у некоторых кастратов-собак в отсутствии брома диференцировка осталась нерасторможенной, в то время как затягивание ее во времени у многих сильных собак вызывает растормаживание? Мы полагаем, что здесь могут иметь значение два обстоятельства: во-первых, различный предел напряжения тормозного процесса у животных с различными типами нервной системы и, во-вторых, происшедшее вследствие кастрации снижение корковой возбудимости могло облегчать борьбу возбуждения и торможения и этим способствовать выдерживанию диференцировки при некотором ее удлинении.
Опыты на кастратах продолжались и далее.
Влияние полового возбуждения на высшую нервную деятельность исследовал Ф. П. Майоров (38). Нормальная корковая деятельность у молодого самца сильного типа нарушалась под влиянием течки у самок. Экспериментатор различными мерами усиливал и ослаблял действие течки на самца перед опытом и наблюдал, как вызванное таким образом и более или менее дозированное половое возбуждение влияет на условные пищевые рефлексы. При этом было установлено развитие гипнотических фаз: наркотической, уравнительной и парадоксальной. Чем сильнее было половое возбуждение, тем сильнее было торможение условных рефлексов, глубже гипнотические фазы. Эти фазы чаще наблюдались во второй половине опыта, когда нарастало двигательное возбуждение животного с явлениями полового возбуждения. Каков физиологический механизм развития гипнотических фаз в данном случае? Предположение о возможности развития сна отпадает совершенно, так как собака никогда не отличалась сонливостью, была, наоборот, в течение этих опытов возбуждена. Наличие полового возбуждения под влиянием течки было очевидным. Можно было представить себе возникновение гипнотических фаз под влиянием сильного возбуждения подкоркового полового центра, аналогично с влиянием голодания в опытах Рикмана. Но и это предположение к объяснению данных явлений не подходит. При наличии общего возбуждения коры и вмешательства запредельного торможения должно было бы получиться более резкое снижение сильных условных рефлексов, тогда как в этих опытах, наоборот, наиболее резкое падение наблюдалось на слабых рефлексах, сильные же понижались меньше, а частью не меняли своей величины. Поэтому чаще всего здесь имела место наркотическая фаза. Кроме того, ни разу не наблюдалось повышения всех условных рефлексов, как это было в опытах Рикмана. Истинное объяснение заключалось в следующем: течка вызывала возбуждение полового центра, что обнаруживалось во всем поведении животного; это возбуждение полового центра обусловливало отрицательную индукцию, распространявшуюся на большую часть коры больших полушарий; на фоне этого внешнего торможения (отрицательной индукции) могли развиваться гипнотические фазы. Это объяснение подтверждалось еще двумя обстоятельствами: 1) тем, что чаще других фаз встречалась наркотическая: слабый рефлекс, естественно, подвергался легче внешнему торможению, чем сильный; 2) на этой же собаке несколько раз наблюдались случаи внешнего торможения с прямой кишки и тогда тоже были аналогичные наркотические фазы. Позднее опыты В. И. Павловой на одном самце полностью подтвердили данные Майорова.
Но такой нервный механизм влияния полового возбуждения на высшую нервную деятельность, соответствующий половой доминанте, нельзя считать универсальным. Он будет иметь место только в тех случаях, когда половое возбуждение (в зависимости от его интенсивности и типа нервной системы) будет концентрированным. Позднее опыты, проведенные С. Н. Выржиковским* на самце сильного типа, показали, что половое возбуждение сначала вызывало повышение всех условных рефлексов, а потом их падение, как в опытах Рикмана с повышением пищевой возбудимости. В этом случае физиологическая сущность явлений сводилась к перевозбуждению, вмешательству запредельного торможения и развитию гипнотических фаз на фоне этого торможения.
* (Неопубликованные опыты.)
Группа работ относилась к явлениям старения высшей нервной деятельности и к способам восстановления падающей работоспособности (Рикман, Бирюков, Ярославцева, Усиевич и Соловейчик). В. В. Рикман (39) занимался дальнейшей проверкой вопроса о трудности системы разнообразных условных раздражителей для стареющей нервной системы. Его собака не выдерживала системы разных раздражителей, при этом у нее происходило падение сильных рефлексов до уровня слабых. Когда к системе из четырех старых раздражителей был добавлен один новый, то рефлекс на последний быстро начал падать. Собака могла нормально работать сначала при упрощенном стереотипе из двух разных раздражителей, а потом уж только из одного (повторение булькания 8 раз), и условный рефлекс при этом не снижался. Таким образом более упрощенный стереотип соответствовал пониженной работоспособности коры.
У другой старой лабораторной собаки упали все условные рефлексы в силу развития гипнотизации (опыты Д. А. Бирюкова) (40). Для устранения этого был применен описанный ранее способ Подкопаева (см. главу XXII), т. е. замена всех старых, "задолбленных" раздражителей на новые. Получился хороший положительный эффект.
В опытах на старой собаке О. П. Ярославцева (41) определяла влияние брома на старую диференцировку в сложной системе раздражителей. Давно выработанная старая диференцировка (на М. 176 к положительному М. 50) с течением времени ослабела и стала давать положительный эффект, в то время как более молодые диференцировки (на тон и кожно-механическое раздражение) оказывались нулевыми. 45 дней применялся ежедневно бром по 3 г. Бром укрепил молодые диференцировки, в связи с этим усилил процесс возбуждения, но не укрепил старой диференцировки: она стала еще более расторможенной. Экспериментатор объяснял это так: бром усиливает торможение, а так как торможение в метрономном "пункте" ослабело, то бром вызвал в нем перенапряжение торможения и, как следствие этого, растормаживание.
Но верно ли, что бром и в больших дозах усиливает тормозный процесс? Впоследствии отказались от такого представления.
Факт постепенного ослабления старой диференцировки наблюдали и другие сотрудники (Андреев, Соловейчик).
Действие брома на нервную систему старой собаки наблюдал М. А. Усиевич (42). Нормальную работу на одряхлевшей собаке можно было проводить только при хроническом бромировании по 2 г NaBr. Бром уничтожал гипнотическое торможение. Прекращение бромирования быстро возвращало животное в прежнее сонливое состояние. Экспериментатор считал, что бром имеет специальное отношение к тормозному процессу и повышает процессы возбуждения путем положительной индукции. Изменение системы или интервалов на фоне бромирования не являлось очень трудной задачей даже для старой собаки.
Особое значение принадлежит работе Д. И. Соловейчика (43). Он исследовал влияние так называемых операций "омоложения" на высшую нервную деятельность. Опыты производились на двух старых собаках-самцах, тормозимого и уравновешенного типов. У первой дважды была сделана операция омоложения по Штейнаху (перевязка семевыводящего протока) и по Воронову (имплантация молодой семенной железы в мошонку). Оба раза был благоприятный результат сроком на 3 мес. У второй собаки сначала в течение двух недель впрыскивался спермин безрезультатно. Затем была сделана операция по Воронову, вызвавшая заметное улучшение высшей нервной деятельности в течение 3 мес. По истечении этого срока эффект омолаживания исчез у обоих животных. Систематически были произведены на обеих собаках сравнения высшей нервной деятельности до операции и после нее, сравнение величины условных рефлексов, скорости их падения при повторении, опыты с обратным порядком системы раздражителей, "сшибки" касалок и "угашение с подкреплением". В итоге испытаний все изменения высшей нервной деятельности под влиянием операций "омоложения" сводились к временному повышению возбудимости и работоспособности коры больших полушарий.
Исследования д-ра Соловейчика несомненно имеют большой теоретический и практический интерес. Жаль, что к этим опытам потом не возвращались и начатую линию исследований не продолжили.
Общим для приведенной группы работ является доказательство понижения возбудимости и работоспособности коры больших полушарий при развитии старости.
1932 г. был исключительно плодотворным годом как в отношении работ сотрудников павловской школы, так и в отношении работ самого Павлова, имеющих чрезвычайно важное значение.
Работа И. П. Павлова совместно с М. К. Петровой (44) "К физиологии гипнотического состояния собаки" представляет конкретный физиологический анализ гипноза и имеет значение для клиники. В работе дано описание опытов на двух собаках, у которых развивалась гипнотизация в экспериментальной обстановке с постепенным торможением двигательной системы, начиная с языка (как более работавшего в данном случае органа), переходя далее на мышцы шеи и кончая мускулатурой туловища. Таким образом здесь наблюдалась картина постепенного охвата гипнотическим торможением отдельных групп мышц и, наконец, всей мускулатуры в целом. Наряду с торможением условной пищевой двигательной реакции была налицо условная секреторная реакция; наблюдались различные фазы диссоциации между секреторной и двигательной реакциями. Большой интерес представляют описанные здесь отрицательная и положительная фазы негативизма: выдвигание кормушки для пищевого подкрепления вызывало отворачивание от кормушки (отрицательная или первая фаза негативизма), убирание кормушки от собаки вызывало у нее движение в сторону кормушки (положительная или вторая фаза негативизма). Был указан и сложный физиологический механизм, лежащий в основе этих явлений.
Физиологический анализ всех наблюдавшихся при гипнотизации явлений был дан с точки зрения локализованного торможения в коре (в области двигательного анализатора), взаимной индукции и взаимодействия заторможенной коры и расторможенной подкорки.
Статья Павлова "О неврозах человека и животного" (45) является ответом на статью американского психиатра (психоаналитической школы) Шильдера - "Соматическая основа невроза"*. Иван Петрович приветствовал мнение Шильдера о том, что в наших "экспериментальных неврозах" на собаках "находятся все явления неврозов". Статья Павлова была посвящена критике принципиального положения Шильдера о том, что "важные эксперименты Павлова и его школы (над неврозами) могут быть поняты только, если мы рассматриваем их в свете наших экспериментов над неврозами", т. е. с точки зрения "психических механизмов" человека. Иван Петрович исходил из методологического положения, что для объяснения необходимо "сведение более сложных явлений на более элементарные, простые явления", а поэтому человеческие неврозы надо объяснить при помощи более простых данных, полученных в эксперименте на животных, а не наоборот, как предлагает Шильдер.
* (Shilder. The Somatic Basis of the Neurosis. (The Journal of Nervus and Mental Disease, vol. 70).)
Надо согласиться с Павловым в том, что основные физиологические механизмы срывов высшей нервной деятельности легче могут быть установлены на животных путем экспериментального анализа и что едва ли физиологическая сущность "экспериментальных неврозов" на животных станет яснее, если рассматривать их с точки зрения особенных "психических механизмов" у человека. Не может быть также подвергнута сомнению научная законность переноса "самых общих основ" экспериментальной патологии высшей нервной деятельности в клинику неврозов у людей для понимания их патофизиологических механизмов. Однако нельзя признать правильным утверждение Павлова о том, что всякое объяснение заключается в сведении сложных явлений к простым. Иван Петрович и здесь оставался верен своему аналитическому методу.
В указанной статье Павлов формулировал важное обобщение относительно основных причин "экспериментальных неврозов". Все наблюдавшиеся в лаборатории случаи патологических отклонений можно свести к действию трех причин: 1) перенапряжение возбуждения, 2) перенапряжение торможения и 3) сшибка возбуждения и торможения.
Был поставлен вопрос: можно ли получить невроз у собак уравновешенного типа? Дальнейшие опыты В. П. Головиной и др. дали утвердительный ответ на этот давно стоявший вопрос.
Доклад И. П. Павлова (46) на Скандинавском неврологическом конгрессе касался тоже неврозов. Доклад был основан на опытах М. К. Петровой на собаке слабого типа, у которой дважды был получен и излечен невроз. Один раз невроз был вызван перенапряжением торможения путем повторного удлинения диференцировки до 5 мин. вместо обычных 30 сек. Невроз был излечен хроническим применением небольших доз брома (по 0,5 NaBr). Другой раз невроз был получен в результате однократного действия сверхсильной трещотки и вылечен тоже бромом по 0,5. Никакие другие средства лечения не действовали.
Таким образом было доказано, что малые дозы брома оказываются действительными для слабых нервных систем. Поэтому Иван Петрович указал на ошибку в прежних заключениях по данному вопросу: "не дозируя соответственно бром, мы думали, что бром у слабых животных никогда не полезен, а в большой дозе только вреден".
Систематизируя свои взгляды по поводу связи различных форм патологии высшей нервной деятельности с типами нервных систем, Павлов на одной из "сред"* дал следующую классификацию, которую мы считаем интересным привести.
* ("Среда", 3 февраля 1932 г.)

Классификация типов и патология
Конечно, это только черновая схема, связи которой возможны, но необязательны. Но и как схема она нуждается в ряде существенных поправок, часть которых потом была внесена экспериментальными исследованиями на собаках и работами павловских клиник.
Статья Павлова "Ответ физиолога психологам" (47) имеет три раздела. Первый раздел - ответ на статью Гётри (Guthrie) "Обусловливание" как принцип обучения*. Этот автор объяснял процесс обучения (научения), исходя из одного принципа conditioning" ("условности" или "обусловливания"), понимаемого им произвольно по-своему. Он предложил другое толкование основных фактов павловской школы. Так например, по его мнению, условный секреторный рефлекс образуется не на условный сигнал (звонок и др.), а на центростремительные импульсы, сопровождающие ориентировочный рефлекс, который возникает на какой-либо сигнал (звонок и др.); запаздывание Павлов яко бы понимает, как какую-то "таинственную латенцию"; угасание, по мнению автора, зависит не от интервалов, как думает Павлов, а от числа повторений; нарушение угасания посторонними раздражителями американский психолог вольно объяснял тем, что они "дезорганизуют позу и окружение"; он по-своему объяснял генерализацию условного рефлекса, исходя из того, что новые рефлексы тоже вызваны ориентировочной реакцией и т. д. и т. п. Такова "критика". Иван Петрович объяснял ошибки автора склонностью к философской дедукции и игнорированием и непониманием физиологических фактов.
* (Edwin R. Guthrie. Conditioning as a Principle of Learning. Psychological Review, 1930, vol. 37, № 5.)
Второй раздел - ответ на статью Лешли "Основные нервные механизмы в поведении" и другие его работы*.
* (K. S. Las hie y. Basis neural mechanisms in behavior. Psychological Review, vol. 37, № 1. K. S. Lashley. Brain mechanisms and intelligence и др.)
Здесь Павлов защищал рефлекторный принцип против Лешли, отрицавшего рефлекторную теорию. Одновременно с этим Иван Петрович дал сильную критику опытов Лешли на крысах и обезьянах и изложил свои представления о корковой локализации. С этой критикой Лешли пришлось считаться, признать некоторые методологические ошибки, а впоследствии и умерить свои возражения против рефлекторной теории. В этом разделе статьи Павлов формулировал три методологических принципа рефлекторной теории, из которых он исходил: принцип детерминизма, принцип анализа и синтеза и принцип структурности. Этими основными положениями утверждались причинное, каузальное мышление, единство анализа и синтеза и неразрывная связь корковых функций с конкретной мозговой, материальной структурой. Этими материалистическими принципами Павлов руководствовался во всей своей работе по новой физиологии больших полушарий. Методологические принципы Павлова составят предмет нашего дальнейшего обсуждения (в специальной главе).
Третий раздел представляет сопоставление физиологии и психологии на примерах анализа сложного поведения животных. Иван Петрович привел два факта: случай, когда лабораторная собака "научилась" сама вытряхивать сухарный порошок из трубки в кормушку, и случай Келера*, когда собака, видевшая перед решеткой мясо, "научилась" делать обходный путь, - и дал им совершенно ясное физиологическое объяснение. Иван Петрович исходил из существования двусторонней проводимости в коре больших полушарий.
* (Пример взят из книги В. Келера. Исследование интеллекта антропоидов)
В заключение Павлов изложил сущность своего мировоззрения. "Человек есть, конечно, система (грубее говоря - машина)*, как и всякая другая в природе, подчиняющаяся неизбежным и единым для всей природы законам; но система, в горизонте нашего современного научного видения, единственная по высочайшему саморегулированию. Разнообразно саморегулирующиеся машины мы уже достаточно знаем между изделиями человеческих рук. С этой точки зрения метод изучения системы - человека тот же, как и всякой другой системы: разложение на части, изучение значения каждой части, изучение связи частей, изучение соотношений с окружающей средой и в конце концов понимания на основании всего этого, ее общей работы и управления ею, если это в средствах человека. Но наша система в высочайшей степени саморегулирующаяся, сама себя поддерживающая, восстанавливающая, поправляющая и даже совершенствующая" (стр.571) (47). Дважды говоря здесь о "системе-машине", Павлов хотел подчеркнуть верность своему механистическому принципу, неоднократно формулированному им и ранее. Однако, на деле он часто изменял ему!
* (Курсив мой. - Ф. М.)
Исключительное значение принадлежит работе Павлова "Проба физиологического понимания симптомологии истерии" (48). Во-первых в ней в качестве предпосылок для физиологического анализа истерии были систематизированы старые экспериментальные данные и приведено несколько новых физиологических представлений. Во-вторых, дана физиологическая трактовка истерии в целом и отдельных ее симптомов. В-третьих, затронут ряд больших вопросов, как-то: вопрос о корковые "пунктах", основной вопрос о взаимоотношении возбуждения и торможения, вопрос об изменениях высшей нервной деятельности в старости. В-четвертых, в этой работе впервые было изложено учение Павлова о корковых сигнальных системах и о двух "специфических" человеческих типах высшей нервной деятельности.
В таком порядке и рассмотрим содержание работы. При изложение сущности нового учения о работе больших полушарий Иван Петрович писал, что "учение об условных рефлексах, или физиология высшей нерв ной деятельности, занимается изучением законов динамики* этих рефлексов" (т. е. условных и безусловных)** "при нормальной и патологической жизни" (стр. 6) (48). Следует отметить сделанное Иваном Петровичем сопоставление условного рефлекса, как специально корковогс явления с суммационным рефлексом, осуществляющимся в центральной нервной системе благодаря иррадиированию раздражительного процесса и его суммации с очагом местного возбуждения. "В больших полушариях при их усложненной конструкции и чрезвычайной реактивности и запечатлеваемости иррадиирование раздражительного процесса ведет к образованию временной условной связи, условного рефлекса, ассоциации. В то время как суммационный рефлекс есть моментальное и скоропреходящее явление, условный рефлекс есть постепенно укрепляющееся при вышеуказанном условии хроническое явление, представляющее характерный процесс коры"*** (стр. 7) (48). Иван Петрович неоднократно подчеркивал не только тождество общих закономерностей деятельности коры больших полушарий и центральной нервной системы, но и специфическое отличие закономерностей высшей нервной деятельности. В этой работе мы встречаем новую классификацию коркового торможения: 1) торможение "пассивное" - безусловное (все виды внешнего торможения), 2) торможение "активное"**** - внутреннее (все виды внутреннего, выработанного торможения) и 3) торможение "запредельное" (стоящее ближе к первому). В связи с последним было изложено учение о "пределе работоспособности" коры больших полушарий.
* (Курсив мой. - Ф. М.)
** (Примечания мои. - Ф. М.)
*** (Курсив мой - Ф. М.)
**** (Разделение торможения на "пассивное" и "активное" было предложено Павловым еще в 1931 г. ("Среда", 14 октября 1931 г.), что нашло свое отражение в некоторых работах сотрудников до 1932 г.)
С нашей точки зрения, нельзя признать удачной терминологию "пассивного" и "активного" торможения, так как всякое торможение есть активный процесс, и Павлов, по существу, никогда не изменял своему изначальному представлению о торможении, как об активном нервном процессе задерживания, противоположном возбуждению.
Иван Петрович предложил также различать два вида сна: "активный" и сон "пассивный", разные по своему физиологическому механизму. Сон "активный" исходит от коры больших полушарий и основан на иррадиации "активного" торможения. Сон "пассивный" происходит вследствие падения возбудимости коры и подкорки, как например, в известном клиническом случае Штрюмпеля или в опытах проф. А. Д. Сперанского и д-ра В. С. Галкина, когда после разрушения трех рецепторов (обонятельного, слухового и зрительного) собака впала в глубокое и хроническое сонное состояние. К случаям "пассивного" сна Иван Петрович относил и клинические случаи, приведшие некоторых к признанию несуществующего, по его мнению, подкоркового "центра сна".
На основании новых опытов В. В. Рикмана* (аналогичных опытам Конради - см. главу XXV) Иван Петрович указал на то, что различение разнородных положительных условных рефлексов (т. е. связанных с разными безусловными) происходит на основании процесса концентрирования возбуждения. Позднее Иван Петрович должен был согласиться с тем, что и здесь процесс торможения должен принимать участие в форме отрицательной индукции.
* (Неопубликованные опыты.)
На основании опытов Рикмана* с растормаживанием тормозной фазы и торможением возбудительной фазы запаздывающего рефлекса при помощи раздражителей различной силы, Павлов сформулировал также новый закон иррадиации и концентрации возбуждения и торможения в зависимости от силы раздражителя: "при слабом напряжении этих процессов они с первого момента и с места их возникновения иррадиируют, при достаточно сильном - концентрируются, и при чрезвычайно сильном - опять иррадиируют" (стр. 6) (48). Этот закон опирался на многочисленный ряд и других фактов.
* (Неопубликованные опыты.)
Далее было изложено учение о типах в его новой, упоминавшейся ранее редакции. Иван Петрович говорил о слабом типе как "тормозимом", "у которого оба процесса недостаточны, но часто особенно тормозной процесс" (стр. 14) (48).
Мы хотим здесь подчеркнуть точность наименования слабого типа "тормозимым", а не "тормозным", как некоторые писали. Слабый тип именно является "тормозимым", так как отличается легкой внешней тормозимостью, но не является "тормозным", так как совсем не представляет "специалиста торможения" (в смысле выработанного торможения). Наконец, отметим еще один очень важный момент - это идею Павлова о корковом представительстве всех органов и всех тканей и о влиянии коры на трофику.
Вооруженный такими физиологическими данными Иван Петрович произвел блестящий анализ истерии. Основные положения Павлова следующие: 1) истерия - продукт слабой нервной системы, 2) она характеризуется конституциональным преобладанием подкорки над корой и первой сигнальной системы над второй, истерия - продукт - "художественного" типа высшей нервной деятельности, 3) отличается наклонностью к развитию гипнотических состояний, 4) относительным преобладанием подкорки объясняется эмотивность истеричных, 5) симптом внушаемости и самовнушаемости - результат сильной концентрации возбуждения и сильной отрицательной индукции, отсюда вытекает механизм гипнотических и пост-гипнотических внушений, 6) истерия характеризуется симптомами страха и уходом в болезнь, что осуществляется по закону условных временных связей, 7) могут иметь корковое происхождение многообразные соматические симптомы истерии, 8) аналгезии и параличи суть симптомы коркового торможения, 9) наклонность к разлитому торможению коры, выключающему в первую очередь вторую сигнальную систему, объясняет фантастичность, оторванность от реальной жизни и сумеречные состояния истеричных, 10) так как у истеричных субъектов нет слаженной и взаимно уравновешенной деятельности трех систем: подкорковой и двух корковых сигнальных систем, а, наоборот, имеется наклонность к нарушениям их взаимоотношений, к диссоциации их, то отсюда происходят симптомы нарушения "психического синтеза" и раздвоения "я".
Общее резюме физиологической сущности всей симптомологии истерии таково. На фоне слабости коры больших полушарий имеют место три физиологических явления: 1) "легкая подверженность гипнотическому состоянию", 2) "чрезвычайная зафиксированность и концентрированность нервных процессов в отдельных пунктах коры, благодаря преобладанию подкорки", 3) "чрезмерная сила и распространенность отрицательной индукции*.
* (Павловская концепция истерии в несколько видоизмененной и искаженной форме получила развитие в работе румынского невропатолога Маринеско и его учеников.)
Исходя из такого понимания нервного механизма истерии, Иван Петрович коснулся истерических психозов и дал объяснение случая истерического пуэрилизма (инфантилизма).
Рассмотрим несколько отдельных вопросов, связанных с работой об истерии. Во многих местах этой работы мы встречаемся с представлением о корковых "пунктах" или "районах". Здесь эти "пункты" Иван Петрович понимал не только в функциональном, но и в анатомическом смысле слова.
Отметим также важное замечание, сделанное Павловым но поводу "проклятого вопроса" об отношении между возбуждением и торможением: "Несмотря на массу накопленного материала в физиологии нервной системы вообще и в учении об условных рефлексах, в частности, вопрос об отношении между раздражением и торможением остается вопросом, пока упорно не поддающимся решению. Что это: одно ли и то же, превращающееся одно в другое при определенных условиях, или крепко спаянная пара, вращающаяся при определенных условиях, и открывающая то в меньшей, то в большей мере, то сполна ту и другую свои стороны?" (стр. 21) (48). В другом месте Иван Петрович говорил: "Несмотря на множество изученных нами частных отношений между раздражительным и тормозным процессами, общий закон связи этих процессов до сих пор не поддается точной формулировке" (50).
История учения о высшей нервной деятельности показала, что этот основной вопрос не мог быть разрешен одним эмпирическим накоплением хотя бы и бесконечного множества фактического материала*.
* (Дальнейшее рассмотрение этого вопроса см. на стр. 340-342.)
При изложении своих представлений о механизмах истерии Иван Петрович коснулся характеристики корковой деятельности в старости. Основное - это понижение реактивности мозга, "естественное падение раздражительного процесса". В силу этого старческий мозг может реагировать, сосредоточиваясь только на каком-либо одном раздражении и развивая отрицательную индукцию на все остальное. Это ошибочно называют "стариковской рассеянностью", тогда как, на самом деле, это есть "сосредоточенность, но невольная, пассивная, дефектная"*.
* (Еще ранее этого Иван Петрович дал физиологический анализ dementia senilis (старческого слабоумия), исходя из двух установленных опытами на старых животных положений: 1) понижение реактивности коры больших полушарий, 2) ослабление функции коркового торможения, "Среда" 14 мая 1930 г.)
Новая глава физиологии больших полушарий открывалась учением Павлова о корковых сигнальных системах и о двух специфических человеческих типах высшей нервной деятельности. Корковая деятельность базируется на подкорковой. В самой же корковой деятельности надо различать две "корковых сигнальных системы". "Первая сигнальная система" - это система, связанная с непосредственным восприятием внешнего мира посредством органов чувств. Она присуща и животным и человеку. "Вторая сигнальная система "связана с развитием речи, слова, понятий. Это - система словесных сигналов действительности. Она свойственна только человеку. "Этим вводится новый принцип нервной деятельности - отвлечение и вместе обобщение бесчисленных сигналов предшествующей системы" (стр. 31) (48). Вторая сигнальная система базируется на первой. Таким образом высшая нервная деятельность человека складывается из взаимодействия трех систем:
Такова схема высшей нервной деятельности человека. В зависимости от преобладания первой или второй сигнальных систем люди делятся на два типа: художественный и мыслительный. У "художников" преобладает первая корковая сигнальная система, они "захватывают действительность целиком, сплошь, сполна, живую действительность, без всякого дробления, без всякого разъединения". У "мыслителей" преобладает вторая корковая сигнальная система, они разлагают действительность на части, "дробят ее, и тем как бы умерщвляют ее, делая из нее какой-то временный скелет" и затем пытаются соединить все в целое. Это - два крайних человеческих - типа. В большинстве же случаев наблюдаются средние, промежуточные положения.

И. П. Павлов на заседании в психиатрической клинике. 1932 г. Позади И. П. Павлова: И. Р. Пророков и Вс. И. Павлов

И. П. Павлов на заседании в нервной клинике. 1932 г. Сидит рядом с И. В. Павловым проф. С. Н. Давиденков и проф. М. К. Петрова
Это представление Иван Петрович развивал и в следующие годы, связав истерию с художественным типом, а психастению (в его понимании) с мыслительным.
Связывая эволюцию второй сигнальной системы с развитием лобных долей больших полушарий, Павлов допускал, что "у художников деятельность больших полушарий, протекая во всей их массе, затрагивает всего меньше лобные их доли и сосредоточивается главнейшим образом в остальных отделах; у мыслителей, наоборот, преимущественно в первых" (стр. 30) (48). Впоследствии (1934 г.) Иван Петрович отказался от такой локализации.
В предисловии к книге А. Г. Иванова-Смоленского* Павлов (49) высказал свою давнишнюю мечту "о возможности слития субъективного с объективным": "Наступает и наступит, осуществится естественное и неизбежное сближение и наконец слитие психологического с физиологическим, субъективного с объективным - решится фактически вопрос, так долго тревоживший человеческую мысль. И всякое дальнейшее способствование этому слитию есть большая задача будущего науки" (стр. 459) (49).
* (А. Г. Иванов-Смоленский. Основные проблемы патологической физиологии высшей нервной деятельности человека. Госмедиздат, 1933.)
Такое завершение получила проблема физиологии и психологии, красной нитью проходившая через всю историю учения Павлова. Однако оставался неразрешенным существенный вопрос: на основе каких методологических предпосылок должно осуществляться это слияние субъективного с объективным? К этому вопросу мы вернемся.
Доклад Павлова "Физиология высшей нервной деятельности" (50) на XIV Международном физиологическом конгрессе в Риме (на ряду с его работой об истерии и докладом о динамической стереотипии) внес ряд существенных дополнений к систематическому изложению учения, данному в "Лекциях" 1927 г. Многое из изложенного в брошюре об истерии и в докладе о динамической стереотипии было здесь повторено. Поэтому мы остановимся только на некоторых моментах. Иван Петрович предложил отличать понятие "высшей нервной деятельности" (= психической") от "низшей нервной деятельности" (= деятельности нижележащих отделов головного и спинного мозга). Он указал их общие основные законы: закон иррадиирования и концентрирования и закон взаимной индукции. Здесь же мы находим ясное указание на физиологический механизм растормаживания: возбуждение иррадиирует на тормозные "пункты" и, смывая в них торможение, делает их на это время положительно действующими. В докладе было изложено учение о "пределе работоспособности" коры и о "запредельном торможении". При этом была высказана новая идея о возможном единстве внутреннего и запредельного торможения: "Можно думать, что своеобразное внутреннее торможение есть тоже запредельное торможение, при чем интенсивность раздражения как бы заменяется его продолжительностью". Далее, Иван Петрович дал синтез двух теорий сна: корковой теории "сна и теории подкоркового "центра сна"" Факты второй теории он объяснил, как случаи патологического или экспериментального выключения проприоцептивных и интероцептивных импульсов, обычно поддерживающих соответствующий корковый тонус. Тогда происходит падение корковой возбудимости, что благоприятствует развитию сонного торможения в коре и дальнейшей иррадиации его на подкорку.
В этом докладе Иван Петрович сообщил о неопубликованных опытах Рикмана с действием посторонних раздражителей различной силы на запаздывающий рефлекс.
Следует отметить гипотезу Ивана Петровича о внутренней связи торможения с ассимиляцией, а возбуждения с диссимиляцией.
Остался невыясненным вопрос: "каким образом кинэстезическое раздражение связано с соответствующим ему двигательным актом: безусловно или условно?" На этот вопрос нет ясного ответа и до сих пор. В докладе была подчеркнута необходимость теснейшей связи физиологии и психологии и обращено внимание на наметившееся в значительной части американской психологии сближение с физиологией.
"Я полагаю, - говорил Иван Петрович перед конгрессом физиологов мира - что сейчас имею последний случай выступать перед общим собранием моих^ коллег и поэтому позволяю себе предложить вашему вниманию общий, наиболее систематизированный и краткий итог моей, вместе с моими дорогими сотрудниками, последней работы, составляющей целую половину всей моей физиологической деятельности... Передаю я этот итог с горячей мечтой о величественном горизонте все более и более открывающемся перед нашей наукой и об ее все углубляющемся влиянии на человеческую натуру и судьбу" (стр. 602) (50).
В заключение главы отметим важнейшие моменты в развитии учения Павлова, получившие свое освещение в 1932 г. Они следующие:
1) учение о системности в работе коры больших полушарий,
2) теория корковой работоспособности (учение о пределе работоспособности и запредельном торможении),
3) постановка основного вопроса о взаимоотношении возбуждения и торможения,
4) введение "короткого способа раздражения",
5) учение о корковых сигнальных системах и о специфических типах высшей нервной деятельности человека,
6) разрешение противоречий между павловской теорией сна и теорией подкоркового центра сна,
7) физиологический анализ развития гипноза,
8) физиологический анализ "травматического невроза",
9) новое учение об истерии,
10) физиологический механизм старческих особенностей высшей нервной деятельности,
11) вопрос о механизме растормаживания и вопрос о возможности растормаживания одного вида внутреннего торможения другим,
12) проблема физиологии и психологии, как проблема "слития" объективного и субъективного.
Литература
1. И. П. Павлов. Динамическая стереотипия высшего отдела головного мозга. Доклад на X Международном психологическом конгрессе в Копенгагене 24 августа 1932 г. Двадцатилетний опыт, цитировано по изд. 1938 г.
2. Э. А. Асратян. Системность работы больших полушарий головного мозга. Доклад в Обществе физиологов в январе 1933 г. Труды физиологических лабораторий акад. И. П. Павлова, 1938, т. VIII.
3. Э. А. Асратян. Системность работы больших полушарий головного мозга. Доклады Академии наук СССР, 1934, 1, 8, 510.
4. Г. В. Скипин. О системности в работе больших полушарий. Доклад в Обществе физиологов в январе 1933 г. Труды физиологических лабораторий акад. И. П. Павлова, 1938, т. VIII.
5. Г. В. Скипин. Косность, инертность нервных процессов есть одно из основных функциональных свойств высших отделов головного мозга животного. Там же, 1938.
6. В. И. Павлова. Неопубликованные опыты.
7. В. В. Рикман. Неопубликованные опыты.
8. Л. О. Зевальд. Материалы к вопросу о системности. Труды физиологических лабораторий акад. И. П. Павлова, 1941, т. X, стр. 321.
9. С. Н. Выржиковский. Неопубликованные опыты.
10. М. А. Усиевич. О решении нервной системой собаки трудной задачи. Труды физиологических лабораторий акад. И. П. Павлова, 1938, т. VIII.
11. Ф. Д. Василенко. К вопросу об условном рефлексе на время. Труды физиологических лабораторий акад. И. П. Павлова, 1932, т. IV, стр. 310.
12. С. В. Клещов. Влияние небольших изменений промежутков времени между раздражителями на величину условного рефлекса. Труды физиологических лабораторий акад. И. П. Павлова, 1941, т. X, стр. 255.
13. С. И. Гальперин. Значение промежутков времени между условными раздражителями для величины условных рефлексов. Там же, 1941, стр. 249.
14. И. Р. Пророков. Одно из нарушений правила о связи величины эффекта с силой раздражения. Труды физиологических лабораторий акад. И. П. Павлова, 1932, т. IV, стр. 288.
15. Н. В. Виноградов. Замаскирование закона силы раздражителей под влиянием индукции. Там же, 1932, стр. 298.
16. В. К. Федоров. Зависимость эффекта от силы раздражителей при кислотных условных рефлексах. Труды физиологических лабораторий акад. И. П. Павлова. 1938, т. VIII.
17. Н. А. Подкопаев. Выработка условного пищевого рефлекса и диференцировки по месту из слабого и постепенно усиливаемого электрического раздражения кожи. Труды физиологических лабораторий акад. И. П. Павлова, 1932, г. IV, стр. 168.
18. Л. С. Григорович. К вопросу о нейтральном поле между полями возбуждения и торможения в коре больших полушарий мозга собаки. Там же, 1932, стр. 244.
19. О. П. Ярославцева. О разграничении районов возбуждения и торможения в звуковом анализаторе коры больших полушарий собаки. Там же, 1932, стр. 258.
20. П. С. Купалов и Б. Н. Луков. Действие короткого применения условного раздражителя. Доклад в Обществе физиологов, 11 ноября 1932, Архив биологических наук, 1933, т. XXXIII, вып. 5-6, стр. 665.
21. П. С. Купалов. Угашение условного рефлекса при длинном и коротком применении условного раздражителя. Доклад в Обществе физиологов 11 ноября 1932 г. Архив биологических наук, 1933, т. XXXIII, вып. 5-6, стр. 679.
22. П. С Купалов и Н. Н. Павлов. Действие короткого условного раздражения при запаздывающем условном рефлексе. Доклад в Обществе физиологов, 11 ноября 1932 г. Физиологический журнал СССР, 1935, т. XVIII, вып. 5, стр. 734.
23. П. С. Купалов и П. К. Денисов. Величина условных рефлексов собак, в освещенной и затемненной камерах. Доклад в Обществе физиологов 11 ноября 1932 г., Архив биологических наук, 1933, т. XXXIII, вып. 5-6, стр. 689.
24. А. Г. Иванов-Смоленский. Взаимное растормаживание тормозных условных рефлексов. Труды физиологических лабораторий акад. И. П. Павлова, 1932, т. IV, стр. 178.
25. А. Г. Иванов-Смоленский. Пассивно-оборонительные рефлексы и сильный тип нервной системы. Иррадиация и концентрация торможения, как фазы действия тормозного раздражителя. Там же, 1932, стр. 208.
26. А. Г. Иванов-Смоленский. Фазовые изменения в нормальном балансе между раздражением и торможением как последействие запаздывающего рефлекса. Там же, 1932, стр. 229.
27. Г. В. Скипин. К характеристике типов нервной системы собак с функциональной слабостью высших отделов центральной нервной системы. Труды физиологических лабораторий акад. И. П. Павлова, 1938, т. VIII.
28. И. Р. Пророков. К характеристике возбудимого (безудержного) типа нервной системы. Труды физиологических лабораторий акад. И. П. Павлова, т. X, стр. 188.
29. В. В. Яковлева. Влияние суммированных раздражителей на нервную систему собаки возбудимого типа. Труды физиологических лабораторий акад. И. П. Павлова, 1932, т. IV, стр. 267.
30. В. В. Яковлева. Перенапряжение тормозного процесса и влияние на него бромистого натрия и тренировки. Труды физиологических лабораторий акад И. П. Павлова, 1940, т. IX, стр. 193.
31. Н. А. Подкопаев. Соотношение иррадиации и концентрации тормозного процесса в течение необычно длительного применения диференцировки. Труды физиологических лабораторий акад. И. П. Павлова. 1932, т. IV, стр. 254.
32. О. П. Ярославцева. Опыт продолжительного столкновения процессов торможения и возбуждения в звуковом анализаторе собаки. Труды физиологических лабораторий акад. И. П. Павлова, 1938, т. VIII.
33. С. В. Клещов. Случай утрированной пищевой двигательной реакции при выработке диференцировки. Труды физиологических лабораторий акад. И. П. Павлова, 1940, т. IX, стр. 163.
34. В. В. Рикман. Обнаружение давних следов раздражения центров оборонительной реакции как аналог травматического невроза. Труды физиологических лабораторий акад. И. П. Павлова, 1932, т. IV, стр. 102.
35. М. К. Петрова. Влияние кастрации на условно-рефлекторную деятельность и общее поведение собак различного типа нервной системы. Труды физиологических лабораторий акад. И. П. Павлова, 1936, т. VI, стр. 1.
36. М. К. Петрова. Изменение тонуса больших полушарий у собак-кастратов под влиянием повышения пищевой возбудимости. Труды физиологических лабораторий акад. И. П. Павлова, 1937, т. VII.
37. М. К. Петрова. Перенапряжение тормозного процесса у собак-кастратов различного типа нервной системы. Там же. 1937.
38. Ф. П. Майоров. К вопросу о влиянии полового возбуждения на высшую нервную деятельность у собак. Доклад в Обществе физиологов 11 ноября 1932 г. Архив биологических наук, 1935, т. XXXVIII, вып. 2, стр. 223.
39. В. В. Рикман. Неопубликованные опыты.
40. Д. А. Бирюков. К вопросу о восстановлении ослабленной условно-рефлекторной деятельности. Труды физиологических лабораторий акад. И. П. Павлова, 1932, т. IV, стр. 349.
41. О. П. Ярославцева. Влияние брома на старую диференцировку в сложной системе условных раздражителей. Труды физиологических лабораторий акад. И. П. Павлова, 1938, т. VIII.
42. М. А. Усиевич. О действии бромистых препаратов на нервную систему старой собаки. Там же, 1938.
43. Д. И. Соловейчик. Влияние так называемых операций омоложения на высшую нервную деятельность. Экспериментальное исследование на собаке по методу условных рефлексов. Труды физиологических лабораторий акад. И. П. Павлова, 1932, т. IV, стр. 14.
44. И. П. Павлов (совместно с М. К. Петровой). К физиологии гипнотического состояния собаки. Труды физиологических лабораторий акад. И. П. Павлова, 1932, т. IV. Двадцатилетний опыт.
45. И. П. Павлов. О неврозах человека и животного. Статья в The Bulletin of the Battle Creek Sanitarium and Hospital Clinic, 1932. Двадцатилетний опыт.
46. И. П. Павлов. Пример экспериментально произведенного невроза и его излечение на слабом типе нервной системы. Доклад на VI Скандинавском неврологическом конгрессе в Копенгагене, 25 августа 1932 г. Двадцатилетний опыт.
47. И. П. Павлов. Ответ физиолога психологам. Статья в Psychological Review, 1932, vol. 39, № 2.
И. П. Павлов. Двадцатилетний опыт, цит. по изд. 1938 г.
48. И. П. Павлов. Проба физиологического понимания симптоматологии истерии. Брошюра, изд. Академии наук СССР, 1932, (L'Encephale, 1933, XXVIII, 4). (Цитировано по брошюре). Двадцатилетний опыт.
49. И. П. Павлов. О возможности слития субъективного с объективным (предисловие в книге А. Г. Иванова-Смоленского). Двадцатилетний опыт, цитировано по изд. 1932 г.
50. И. П. Павлов. Физиология высшей нервной деятельности. Доклад на XIV Международном физиологическом конгрессе в Риме, 2 сентября 1932 г. Природа, 1932, № 11-12, 1139. Двадцатилетний опыт, цитировано по изд. 1938 г.
При использовании материалов сайта активная ссылка обязательна:
http://anfiz.ru/ 'Анатомия и физиология человека'