Глава XXVI - 1930
Ю. М. Конорский и С. Миллер в 1928 г. в Варшаве делали следующие опыты. Звуковое раздражение они сопровождали пассивным сгибанием ноги собаки и подкрепляли эту комбинацию едой. В результате таких сочетаний одно звуковое раздражение начало вызывать активное сгибание ноги. Авторы были склонны рассматривать эти рефлексы, как принципиально отличные от условных, установленных павловской школой, и описали их, как условные рефлексы второго типа. Павлов не согласился с этим и с целью проверки и физиологического анализа предпринял ряд исследований, в которые потом включились и Конорский с Миллером, работавшие в павловской лаборатории продолжительное время.
Развитие экспериментального анализа произвольных движений в павловской школе имело и свою внутреннюю историю, оно было связано с опытами Н. И. Красногорского (1911 г.), образовавшего условные пищевые рефлексы на проприоцептивные раздражения при сгибании ноги у собак. Эти исследования привели Павлова к важному утверждению, что двигательная область коры больших полушарий есть такой же анализатор ("двигательный анализатор"), как и остальные.
Группа сотрудников параллельно в разных вариациях и на животных разного типа производила тщательное исследование поставленного вопроса. Такая организация экспериментальной работы довольно характерна для стиля научно-исследовательской деятельности павловской школы. Это имело свои несомненные преимущества, так как позволяло в сравнительно короткие сроки всесторонне и глубоко исследовать тот или иной вопрос и притом с гарантией точности благодаря постоянному взаимному контролю.
Для разрешения указанной задачи Г. В. Скипин (1) пользовался секреторно-двигательной пищевой методикой. Учитывались слюнная реакция и движения ноги собаки; и то и другое одновременно записывалось на кимографе. Скипин делал следующие опыты на одной собаке сангвинического типа. Шум на 5 сек. предшествовал пассивному сгибанию правой передней ноги (при помощи специального механизма) и далее оба раздражения (экстероцептивное и проприоцептивное) совпадали 20 сек., подкрепляясь после этого едой. Условный слюнной рефлекс на такой комплекс образовался с 4-5 сочетания. С 8-го раза отставленный (на 20 сек.) шум тоже вызывал активное поднимание ноги. Было сделано более ста таких сочетаний с едой. Примененные после них и все другие условные раздражители стали сопровождаться активным подниманием правой передней ноги.
Дальнейшие опыты заключались в выработке условного тормоза из проприоцептивного раздражения. Положительным раздражителем был стук метронома (М. 120) или звонок. Пассивное сгибание правой задней ноги плюс М. 120 не подкреплялись. Условный тормоз на сгибание выработался медленно и волнообразно и не был абсолютным. Медленность и затрудненность в его выработке, повидимому, зависела от слабости проприоцептивного раздражения по сравнению с основным раздражителем.
На основании полученных кривых Скипин представил анализ состояний коркового двигательного анализатора в разные моменты. Наблюдались явления отрицательной индукции с пищевого центра на корковый двигательный и положительной индукции с заторможенного пищевого центра на двигательный. Были неоднократно констатированы факты самоиндукции (положительной) в двигательном анализаторе. Это выражалось в том, что после фазы задержки движений ноги наступала фаза усиленных движений.
В качестве общего вывода следовало, что раздражения от двигательного аппарата сгибаемой конечности могут быть применены как в качестве компонента при выработке комплексных условных пищевых рефлексов, так и в качестве тормозного агента при выработке условного тормоза. Было установлено, что корковый двигательный анализатор имеет ту особенность, что "в нем заключен и эффекторный, имеющий непосредственную связь с периферией, аппарат" (1). Таким образом вносилось существенное дополнение в понимание физиологической сущности коркового двигательного центра.
В заключении своей работы автор правильно подчеркивал, что, кроме факторов внешнего мира, актуальное значение в высшей нервной деятельности имеют факторы "внутреннего мира" (в данном случае, проприоцептивные раздражения).
Во второй своей работе Г. В. Скипин (2) выяснял условия для выявления ассоциации между двумя возбужденными от экстероцептивного и проприоцептивного раздражений участками коры. Почему в начале указанных исследований сотрудники Павлова (А. М. Павлова, М. К. Петрова и др.) не могли получить активного поднимания ноги на экстероцептивное раздражение, которое получили Конорский и Миллер? Предполагали, что образованию этой корковой связи мешает постоянная отрицательная индукция от старых, прочных условно-рефлекторных связей. Поэтому Скипин проводил свои эксперименты на двух свежих, не бывших ранее в употреблении собаках. Сначала опишем опыты, производившиеся на одной собаке. Вырабатывался условный пищевой рефлекс на комплекс: 5 сек. действия М. 120-30 сек. М. 120 плюс пассивное сгибание правой передней ноги. Рефлекс быстро образовался. При этом один стук метронома имел большой секреторный эффект, а одно пассивное сгибание - незначительный. Активного поднимания ноги на М. 120 не происходило. Тогда было сделано предположение: может быть, слаба раздражительная сила сгибания? Чтобы физиологически усилить это проприоцептивное раздражение, экспериментатор стал применять только сгибание, сопровождая его едой. Однако от этого условный слюнный рефлекс не увеличился и подъема ноги при действии М. 120 не получилось. Было сделано допущение, что образование связи между М. 120 и сгибанием не происходит вследствие тормозящего влияния поднятия лапы при помощи Механизма. С целью устранить это, экспериментатор начал поднимать когу собаки рукой, сияя в камере около станка. Через некоторое время искомая ассоциация была получена: стук метронома стал вызывать активно? поднимание ноги, но только в порядке своего следового действия. Это значит, что активное поднимание ноги производилось собакой не во время действия метронома, а через несколько секунд после его прекращения.
У второй собаки также был быстро образован условный слюнный рефлекс на комплекс: 5 сек. действия света (50 свечей) - 20 сек. света плюс пассивное сгибание ноги механизмом - еда. При отдельных испытаниях свет вызывал незначительный секреторный эффект, а пассивное сгибание - большой. Но активного поднимания ноги при действии света не было. Сила света была увеличена в два раза до (100 свечей). После этого свет тоже не давал подъема лапы. Экспериментатор решил ослабить проприоцептивный раздражитель путем его неподкрепления при нескольких изолированных применениях, т. е. было произведено частичное угашение. Тогда и у этой собаки получили активное поднимание ноги при действии света.
Таким образом удалось выяснить причины, мешавшие образованию временной связи между экстероцептивным и проприоцептивным раздражениями. Они были, во-первых, в разной возбуждающей силе компонентов и, во-вторых, в тормозящем влиянии обстановки. Следовательно, условный пищевой рефлекс на комплекс экстероцептивного и проприоцептивного раздражений вырабатывался легко. Но образование ассоциации между самими компонентами могло происходить при условии некоторого уравнивания их силовых отношений. Так, после угашения пищевого рефлекса на одно пассивное сгибание, свет стал вызывать активный подъем ноги. Следовое раздражение (от стука метронома) также вызывало активное поднимание ноги.
М. К. Петрова (3) производила такие же опыты не в изолированной камере, а сидя около собаки. Работа велась сначала на двух старых лабораторных собаках, а потом на одной молодой. В опытах Скипина экстероцептивный раздражитель всегда предшествовал в комплексе проприоцептивному. В опытах Петровой был применен и тот и другой порядок: 1) треск предшествовал пассивному сгибанию ноги, 2) совпадал с ним и сопровождался пищевым подкреплением, 3) пассивное сгибание предшествовало треску, 4) совпадало с ним и сопровождалось едой. Вырабатывался также условный рефлекс на комплекс: зрительный раздражитель (белый круг на черном фоне) и пассивное сгибание (отставление = 30 сек.). У старых лабораторных животных не удалось выработать активного сгибания ноги на звуковой и зрительный раздражители, несмотря на несколько сот сочетаний. Это объяснялось действием отрицательной индукции на кинестетические корковые клетки с других анализаторов и малой пластичностью нервной системы подопытных собак. У третьей же молодой собаки, у которой ранее не было выработано никаких условных рефлексов, удалось образовать указанную связь. Здесь опыт проводился так: экспериментатор произносил слово "ногу", сгибал рукой лапу и давал еду. После нескольких сочетаний собака стала сама поднимать ногу на слово - раздражитель "ногу"! Затем было сделано угашение активного сгибания правой передней ноги на слово "ногу"!, после чего было выработано активное сгибание левой передней ноги на стук метронома. Далее вырабатывался условный тормоз из пассивного сгибания задней ноги к положительному стуку метронома. У одной из старых собак он выработался, но вместо сгибания собака стала производить активное разгибание ноги и гипнотизироваться.
Выработку условного тормоза из проприоцептивного раздражителя производил и Н. А. Подкопаев (4) на собаке уравновешенного типа. Звонок был условным положительным раздражителем. Условный тормоз: пассивное сгибание ноги 1 сек. - 2 сек. - звонок плюс сгибание 20 сек. (без еды). Результат: после 29 применений тормозной комбинации полного условного тормоза выработать не удалось. Впрочем, автор сам сослался на малое число поставленных опытов. В ответ на пассивное сгибание собака стала держать эту ногу в положении активного разгибания, что расценивалось как проявление активной обороны животного с сигналом "неприятности" - отсутствия еды.
Таким образом, факты, обнаруженные Конорским и Миллером, были подтверждены: 1) была доказана возможность образования корковой временной связи (ассоциации) между экстероцептивным и проприоцептивным раздражениями, 2) была доказана также возможность выработки условного тормоза из проприоцептивного раздражения.
Дальнейший ход этих работ привел к установлению новых фактов и тонкому анализу "произвольных" движений на основе учения о высшей нервной деятельности. Эти исследования открыли необходимость в свободе "произвольных", "волевых" движений.
Опыты с двигательными рефлексами привели также и к более широкому пониманию самого условного рефлекса. В самом начале работы Павлов и его сотрудники подчеркивали "специфичность" условного рефлекса в том смысле, что "условный рефлекс повторяет безусловный". Это положение справедливо в отношении слюнных (секреторных) рефлексов, где, действительно, условный рефлекс в меньшем масштабе воспроизводит рефлекс безусловный. Но оно не справедливо в случае, скажем, условных пищевых двигательных рефлексов, где условный рефлекс не повторяет безусловного, на основе которого он образован.
Таким образом расширилось и уточнилось понятие самого условного рефлекса.
Г. В. Скипин в связи с упомянутыми работами сделал два наблюдения, касавшиеся латентного периода условных рефлексов и процесса угасания.
Первое наблюдение (5) относилось к вопросу о тормозной природе латентного периода. Быков и Петрова (см. главу XXIII) показали, что истинный латентный период условных рефлексов составляет только очень малую часть регистрируемого "латентного периода". Скипин имел возможность одновременно сравнивать течение секреторной и двигательной реакций. У одной из его собак все условные пищевые раздражители сопровождались активным подниманием ноги, которую собака часто поднимала в интервалах и иногда держала приподнятой на протяжении всего интервала. В начале же условного раздражения она сначала опускала ногу, а потом снова ее поднимала (в порядке осуществления положительной двигательной реакции). Опускание ноги вниз свидетельствовало о возникновении торможения во время латентного периода. Но какова была его природа в данном случае? Здесь были возможны два объяснения: развитие запаздывательного торможения или отрицательная индукция от ориентировочной реакции на начало условного раздражения. У исследователя имелись основания признать правильным первое объяснение. Описанный факт потом многократно наблюдали и другие авторы, работавшие с двигательными рефлексами (Ф. П. Майоров и др.). Таким образом было получено новое Доказательство тормозной природы так называемого "латентного периода".
Второе наблюдение Скипина (6) имело отношение к вопросу о природе угасательного процесса. Опыты делались тоже по секреторно-двигательной методике. Случайные поднимания ноги, привязанной к регистрирующему рычагу, экспериментатор подкреплял пищей. После 2-3 подкреплений собака, съев пищу, сейчас же принималась поднимать соответствующую ногу. Когда условный пищевой рефлекс поднимания ноги укрепился, было сделано его угашение. При этом наблюдалось усиление движений ноги. Затем были выработаны условные пищевые рефлексы на комплексы: свет плюс пассивное сгибание лапы и звонок плюс пассивное сгибание. При угашении рефлекса на такой комплекс наблюдалась следующая картина: слюнная секреция уменьшалась, а поднимания ноги увеличивались, затем внезапно обрывались. Следовательно, развитие угасательного торможения сопровождалось взрывами двигательных реакций, свидетельствовавших о возбуждении двигательного анализатора.
Опираясь на этот факт, автор объяснял сущность угасательного торможения, как перевозбуждение, как одну из фаз возбудительного процесса (по Введенскому-Ухтомскому). Рассмотрим изложенные данные со стороны их возможного механизма. Угасание условного секреторного рефлекса здесь происходило, как обычно, по волнообразной затухающей кривой. Эта волнообразность есть проявление борьбы возбуждения и торможения, как антагонистических процессов. Такая же волнообразность имела место и при развитии угасательного процесса на условном пищевом двигательном рефлексе, что свидетельствовало о борьбе возбуждения и торможения в двигательном анализаторе. Фаза торможения двигательных реакций могла сменяться фазой последовательной положительной индукции (самоиндукции) и т. д. Кроме того, в этом случае возможно было действие положительной индукции с заторможенной секреторной части коркового пищевого центра на его двигательную часть.
Если говорить вообще о процессе угасательного торможения, то мы видим в нем не нарастание возбуждения и перевозбуждения, а, наоборот, постепенное затухание рефлекса. Мы думаем, что концепция торможения, как перевозбуждения, к данному виду коркового торможения не применима, так же как она не применима и к запаздывательному торможению (вопреки мнению Г. В. Скипина) (5).
Но это не значит, что парабиотическая теория торможения Введенского не применима вообще к явлениям коркового торможения. Мы вернемся к этому вопросу при рассмотрении так называемого "запредельного торможения".
К вопросу о взаимоотношении возбуждения и торможения относится ряд исследований этого времени. Одним из способов изучения тонких соотношений противоположных нервных процессов было, одновременное применение тормозного и положительного раздражителей (Анохин, Скипин). П. К. Анохин (7) присоединял к метрономной диференцировке тот или иной положительный раздражитель. Первые экстренные испытания положительного процесса на фоне длящегося тормозного показали "обособленное действие обоих процессов с некоторой тенденцией в сторону положительной индукции для положительного". Систематическое неподкрепление комбинации делало ее тормозной, и, если при доведении до нуля эффекта комбинации в нее экстренно вставляли другой положительный раздражитель, то он с места оказывался значительно заторможенным. Если комбинацию подкрепляли, заканчивая ее положительным раздражителем, то последний приходил к своей нормальной величине. При этом происходило и упрочение тормозного раздражителя в силу отрицательной индукции. Если теперь в комбинацию вставляли какой-либо другой положительный раздражитель, то он с места давал положительный эффект. Таким образом, в случае неподкрепления комбинации диференцировки и положительного раздражителя, на последний распространялось последовательное торможение; в случае подкрепления наблюдалось развитие положительной индукции от действия тормозного раздражителя на эффект положительного. Эти опыты позволяли "более глубоко войти в динамическую связь возбуждения и торможения" (7а).
Г. В. Скипин (8) присоединял условный положительный раздражитель к метрономной диференцировке через различные промежутки времени от начала ее действия. В начале действия тормозного раздражителя наблюдалась временная иррадиация тормозного процесса. По мере удлинения времени действия тормоза происходила постепенная концентрация торможения. У собаки сангвинического типа концентрация наступала при 30 сек. действия тормоза и сохранялась при удлинении его действия до 6 мин. Повторение указанной комбинации без подкрепления вело к образованию условного тормозного рефлекса.
Взаимоотношение возбуждения и торможения определяло и результаты опытов С. А. Петрова (9), который сравнивал скорость образования диференцировок в зависимости от продолжительности времени изолированного действия тормозного раздражителя. На одной собаке Петров вырабатывал по очереди несколько диференцировок на разные раздражители. Скорее образовались диференцировки при отставлении в 15 сек., чем при более коротком - в 5 сек. При очень коротком отставлении (1 сек. - 2 сек.) диференцировка не могла быть выработана, несмотря на 68 ее применений. Следовательно, выработка диференцировки тем труднее, чем короче время отставления диференцировочного раздражителя. Это зависит от того, что торможению, как активному процессу задерживания, приходится преодолевать возбуждение при наименее благоприятных условиях: 1) в начальный момент наиболее раздражающего действия агента и 2) в короткий промежуток времени, недостаточный для развития интенсивного тормозного процесса.
Запаздывание можно рассматривать, как переход торможения, в возбуждение. Но этот переход развивается различно в зависимости от процедуры выработки условных запаздывающих рефлексов. Ф. С. Павлов (10) производил образование запаздывающего пищевого рефлекса на звонок на 3 мин. путем постепенного удлинения отставления с 20 сек. на 5 сек. каждый день. Запаздывание вырабатывалось при отставлении в 95 сек., т. е. были тормозная и положительная фазы; при дальнейшем удлинении у собаки получился "срыв", что выразилось в растормаживании первой фазы и в развитии гипнотического состояния и гипнотических фаз в последействии. Перерыв в применении запаздывающего раздражителя и переход на короткое отставление всех рефлексов избавляли животное от гипнотического состояния.
Переходим к рассмотрению следующего вопроса. Как было видно из изложенного, за последние годы стали интенсивно изучать действие брома на высшую нервную деятельность. Эти исследования имели двойное значение: выяснение механизма действия брома и изучение коркового торможения при помощи брома. Так как бром считался, усилителем и концентратором торможения, то его применяли для устранения гипнотического состояния у собак (опыты Ф. П. Майорова) (11). На двух собаках, постоянно гипнотизировавшихся в экспериментальной обстановке, с успехом был использован бром (NaBr). Для устранения гипнотического состояния были необходимы: определенный промежуток времени от момента введения брома (per os) до начала опыта (45-60 мин.) и определенная доза в зависимости от типа нервной системы и других условий. Отметим, что при больших дозах брома (3,0-4,0) наблюдалось значительное растормаживание диференцировок. При длительном применении таких доз автор наблюдал развитие общего возбуждения, переходившего в дальнейшем в явления атаксии и пареза задних конечностей.
В. В. Яковлева (12) под влиянием "сшибок" получила у возбудимой собаки "срыв" и развитие гипнотизации. Для борьбы с гипнотическим состоянием был применен бром (3,0 NaBr ежедневно). Бромирование продолжалось 11/2 месяца без перерыва. Бром урегулировал нервную деятельность животного, сконцентрировал тормозный процесс и, благодаря положительной индукции, поднял процесс возбуждения. Положительное действие брома проявилось прежде всего на сильных раздражителях, затем на средних и, наконец, на слабых. При вторичном применении бром действовал быстрее и сильнее. В этих опытах, как и в предыдущих, было отмечена растормаживание диференцировок при длительном бромировании сравнительно большими дозами. Долгое хроническое применение брома вызвало отравление животных (понижение всех условных рефлексов и нарушение двигательных функций).
И. С. Розенталь (13) изучал влияние брома на развитие процесса угасания условного пищевого рефлекса. Он производил несколько раз (на одной собаке) опыты острого сплошного угашения с введением бромистого натрия (1,0, 2,0 и 4,0) и без него. Приводим сводку полученных результатов (среднеарифметическое количество условно-рефлекторной слюны при угасании рефлекса на звонок):
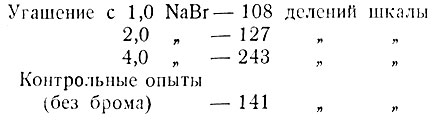
Таким образом, меньшие дозы (1,0 и 2,0) усиливали угасательное торможение, имели тормозящее действие, а большие дозы (4,0), наоборот, ослабляли угасательное торможение, оказывали возбуждающее действие. Последнее экспериментатор объяснял, исходя из павловской теории тормозного действия брома: бром концентрировал торможение, следствием чего была вторичная положительная индукция, вызывающая повышение корковой возбудимости.
В то время Павлов объяснял и возбуждающее действие больших доз брома в опытах Майорова и Яковлевой, как проявление положительной индукции в коре вследствие концентрации торможения бромом. Однако, при этом не было объективных признаков концентрации тормозных процессов, наоборот, диференнировки оказывались значительно расторможенными. Сомнительным было также предположение о существовании в коре необнаруживаемых нами очагов концентрированного торможения (в порядке постоянной жизненной "мозаики" положительных и тормозных "пунктов").
Представление о том, что и большие дозы брома тоже усиливают и концентрируют торможение, впоследствии (1934 г.) пришлось оставить (под влиянием фактов, обнаруженных одновременно Ф. П. Майоровым и С. Д. Каминским на обезьянах и М. К. Петровой на собаках).
Новые материалы были получены при изучении типов нервной системы. Действие сверхсильного раздражителя на нервную систему собаки сангвинического типа изучал И. О. Нарбутович (14). В стереотипе условных раздражителей вырабатывался условный пищевой рефлекс на сверхсильную трещотку. В начале у собаки наблюдалось резкое торможение двигательных реакций, но скоро рефлекс выработался. При этом секреторная реакция была налицо, а двигательная пищевая оказывалась еще заторможенной. Сначала слюнной рефлекс На трещотку был равен слабым рефлексам, потом средним и сильным и, наконец, стал давать постоянный эффект больше сильных. Путем тренировки сверхсильного раздражителя удалось ослабить явления торможения и довести их до полного исчезновения. Итак, даже неразрешимые на первый взгляд для данной нервной организации задачи при помощи постепенной тренировки могли быть разрешены. Эти опыты еще раз подтвердили большую пластичность центральной нервной системы.
Новое в учение о типах внесла работа, проведенная С. Н. Выржиковским и Ф. П. Майоровым (15) на Биостанции им. акад. И. П. Павлова. Они исследовали вопрос о влиянии воспитания и внешней среды на склад и формирование высшей нервной деятельности собак. С этой целью 8 щенят одного помета с самого раннего возраста воспитывались в противоположных условиях: четверо в условиях полной свободы и четверо взаперти, в клетках, из которых их никуда не выпускали. Таким образом первые росли в условиях постоянного взаимодействия со всем многообразием внешнего мира, соприкасаясь с другими животными и людьми; вторые жили в однообразной обстановке клеток собачника, по одиночке. Условия питания были примерно одинаковыми у той и другой группы. Когда эти животные стали взрослыми (около 2 лет), мы приступили к изучению их высшей нервной деятельности. Обнаружилась резкая разница в поведении тех и других собак. У "заключенных" собак в отличие от "свободных" сохранился и во взрослом состоянии тип поведения, свойственный щенятам. Пассивно-оборонительный рефлекс у первых был выражен гораздо резче, чем у вторых. Такая же разница была и в ориентировочном рефлексе. "Заключенные" собаки были сильнее подвержены всякому внешнему торможению, чем собаки "свободные". Однако первые лучше смогли приспособься к условиям изолированной от внешнего мира экспериментальной Остановки, чем вторые. "Свободные" собаки обнаружили наклонность к гипнотизации в однообразной обстановке камеры.
Указанное различие в поведении было результатом разного воспитания, а не врожденного типа нервной системы, так как по счастливому (для экспериментаторов) стечению обстоятельств все животные - заключенные", обнаруживавшие пассивно-оборонительный тип поведения слабых, при систематическом их испытании в опытах оказались животными не слабого, а сильного типа нервной системы.
На основании этих исследований было принято различать тип высшей нервной деятельности (или нервной системы), что составляет генотип и соответствует "темпераменту", и склад высшей нервной деятельности, что является фенотипом и соответствует "характеру".
Разберем теперь несколько отдельных, несвязанных друг с другом вопросов.
В плане изучения безусловных, врожденных рефлексов, как фундамента условных, была поставлена работа И. С. Розенталя (16). Он представил материал наблюдений над поведением щенят в возрасте от первых дней до 6 месяцев. Розенталь прослеживал развитие ориентировочного и оборонительного рефлексов и их взаимодействие. Он предложил различать агрессивно-оборонительный рефлекс (в основе которого лежит возбуждение), пассивно-оборонительный рефлекс тормозного происхождения и пассивно-оборонительный рефлекс с убеганием (основанный тоже на возбуждении).
Это деление может быть нами принято лишь как схема, но и она, как таковая, имеет принципиальный недостаток, состоящий в отрыве возбуждения от торможения. Так, агрессивно-оборонительный рефлекс связан с отрицательной индукцией, пассивно-оборонительный рефлекс тормозного характера может сопровождаться явлениями возбуждения и т. д.
Исследования С. В. Клещова (17) относились к вопросу о том, могут ли животные различать отношения тонов независимо от их высоты. Известный психолог Штумпф считал, что животные этого делать не могут. Был выработан условный пищевой рефлекс на аккорд из двух тонов фисгармонии (квинта и соль 1 октавы). Эта квинта применялась среди систем других условных раздражителей. Опыты заключались в угашении четырех пар других аккордов, действовавших в порядке генерализации: квинты и терции каждой октавы попарно. Угашение квинт всегда давало больший секреторный эффект, чем угашение терций. Это различие возрастало по мере приближения угашаемой пары аккордов к основной квинте. Итак было доказано, что животные (в данном случае собаки) могут различать отношения тонов. Экспериментатор был склонен в этом видеть как бы биологическую предпосылку для возникновения музыки.
В. В. Строганов (18) провел работу, доказавшую важное значение метода условных рефлексов в физиологии труда. Он делал опыты с условными пищевыми рефлексами на собаках до работы и после работы, заключавшейся в перевозке тележек с грузом кирпичей. В процессе приспособления к такой работе наблюдались некоторые изменения в условных рефлексах (увеличение рефлексов на звуковые раздражители и уменьшение на кожные). С образованием установки на работу условные рефлексы возвращались к норме. После работы отмечалось повышение коркового возбуждения: увеличение условных рефлексов, укорочение латентных периодов и ускорение слюнной реакции. Наблюдались также колебания величины рефлексов на протяжении некоторого времени после произведенной работы. Строганов пришел к заключению, что "метод условных рефлексов, выявляющий интимнейшие процессы коры больших полушарий головного мозга, должен получить широкое применение в области вопросов, связанных с физиологией труда".
В дальнейшем работы на эту тему были проведены в лаборатории проф. К. М. Быкова*.
* (Александров. Влияние физической работы на условные рефлексы. Архив биологических наук, 1932, т. XXXII, вып. 5-6.)
Доклад И. П. Павлова на Международном психологическом конгрессе 1929 г. в Нью-Хэвене (США) был напечатан под названием "Краткий очерк высшей нервной деятельности" (19) (статья в американском сборнике "Психология 1930 г."). Здесь были изложены основные законы высшей нервной деятельности, которые Иван Петрович назвал "правилами". Все многообразие явлений можно было свести к следующим шести основным правилам:
1) правило нервного замыкания, ассоциации,
2) правило зависимости величины эффекта от силы раздражения,
3) правило суммации условных рефлексов,
4) правило перехода корковой клетки в тормозное состояние,
5) правило - иррадиирования и концентрирования нервных процессов,
6) правило взаимной индукции нервных процессов.
Первое правило указывает на основную закономерность образования корковой временной связи или ассоциации путем нервного замыкания.
Правило зависимости величины эффекта от силы раздражения ("закон силы") было сформулировано следующим образом: "Чем условный раздражитель сильнее, чем более энергии поступает с ним в большие полушария, тем, при прочих равных условиях, более условно-рефлекторный эффект, т. е. тем энергичнее пищевая двигательная реакция и тем обильнее слюнотечение" (стр. 424) (19). К этому правилу было добавлено новое существенное исключение: "Как можно судить, по некоторым опытам, эта связь эффекта с силой раздражения должна быть довольно точной. Но при этом всегда имеется предел, за которым еще более сильный раздражитель не увеличивает, а начинает уменьшать эффект"*. Мы подчеркиваем это исключение ввиду его принципиального значения. К этому времени Иван Петрович стал обращать внимание на то, что очень сильные и слабые условные раздражения почему-то осложняются торможением.
* (Курсив мой - Ф. М.)
С тем же пределом и вмешательством торможения имели дело в случае суммации условных рефлексов. При комбинировании слабых раздражителей обычно получалась арифметическая сумма. Комбинация слабого раздражителя с сильным давала увеличение эффекта до какой-то предельной величины. При суммировании же двух сильных эффект "как выходящий за предел"* делался меньше каждого из компонентов. Таково было правило суммации. Оно вытекало из опытов В. В. Рикмана** и других.
* (Курсив мой - Ф. М.)
** (Неопубликованные опыты.)
Правило перехода корковой клетки в тормозное состояние подчеркивало, что, кроме процесса раздражения "тот же внешний условный раздражитель может вызывать в корковой клетке противоположный процесс - процесс торможения", если он действует один без сопровождения безусловным раздражением (стр. 424) (19).
Иррадиация и концентрация нервных процессов и взаимная индукция рассматривались в их единстве и динамическом взаимодействии. Из этого вытекало представление о сложном, волнообразном характере иррадиации (стр. 426) (19).
Павловские шесть правил были тем логическим каркасом, на котором в более или менее систематическом порядке можно было расположить большой фактический материал.
Для Павлова являлось чрезвычайно характерным стремление понять физиологические факты с биологической точки зрения. Так, им был поставлен вопрос о биологическом значении последовательного торможения. Каков биологический смысл длительного распространения торможения и иногда на все полушария? "Есть ли это недостаток косность аппарата или здесь определенное нарочное явление, биологическое значение которого мы еще не понимаем?" (стр. 427) (19).
Иван Петрович указал на три основных темы, стоявшие перед физиологией больших полушарий: 1) изучение "сложнейших безусловных рефлексов", как фундамента внешней деятельности животного организма, 2) изучение деятельности коры и 3) способа соединения и взаимодействия подкорковых центров и коры.
По этим трем основным линиям и шло развитие экспериментальной работы.
Лекция Павлова "Физиология и патология высшей нервной деятельности" (20), прочитанная врачам Государственного института усовершенствования врачей в Ленинграде, по существу повторяла содержание предыдущей статьи, только в несколько другой форме*. Остановим наше внимание лишь на некоторых моментах этой работы. Павлов подчеркивал свое понимание торможения, как "активного нервного процесса" (стр. 11) (20). Обычная для него пространственная теория корковых "пунктов" и здесь получила свое надлежащее выражение. Иван Петрович говорил также об экспериментальном проявлении "неврастении" и "истерии" у собак, дал физиологическое объяснение некоторых симптомов шизофрении. Была высказана важная идея о сохранении нервных следов не только в коре, но и в подкорке. Высшая нервная деятельность рассматривалась, как взаимодействие коры и подкорки, в котором существенное значение принадлежит взаимно-индукционным отношениям. Надо отметить, насколько расширилось и усложнилось представление о взаимной индукции со времен ее открытия в явлениях корковой деятельности.
* (Эта лекция не вошла в сборник И. П. Павлова "Двадцатилетний опыт" и была издана Медгизом отдельной брошюрой. 1930 г.)
Мы позволим себе здесь несколько отступить от текста рассматриваемой работы Ивана Петровича и представить систему взаимной индукции в деятельности больших полушарий в виде следующее схемы (рис. 5):
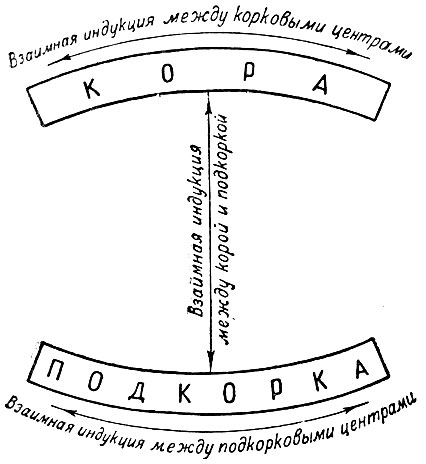
Рис. 5. Схема взаимной индукции
Эта схема показывает сложное динамическое переплетение индукционных влияний в горизонтальном и вертикальном направлениях, развертывающихся в деятельности больших полушарий головного мозга. К этому времени Иван Петрович также установил деление корковой взаимной индукции на "внешнюю" (с одного центра на другой) и "внутреннюю" (в пределах одного и того же центра), т. е. "самоиндукцию".
Большой задачей физиологии и патологии больших полушарий является поставленная Павловым задача дальнейшего изучения взаимодействия коры и подкорки. Ни одно корковое явление не может нами рассматриваться вне этого основного взаимодействия.
Лекция Ивана Петровича заканчивалась рассмотрением вопроса о соотношении физиологии и психологии в изучении психической деятельности. Классическая ассоциативная психология представляла анализ, а ее соперница Gesltaltpsychologie - одностороннее выражение синтеза. Физиология высшей нервной деятельности, по мнению Павлова, давала объединение того и другого направления, т. е. и анализ и синтез. Это нашло свое конкретное выражение в новом учении Павлова о "системности". "Для нас совершенно ясно, что кора больших полушарий представляет собой сложнейшую функциональную мозаику из отдельных элементов, каждый из которых имеет определенное физиологическое действие - положительное или тормозное. С другой стороны, также несомненно, что все эти элементы объединены в каждый данный момент в систему, где каждый из элементов находится во взаимодействии со всеми остальными" (стр. 35-36) (21).
С этого времени начинаются смелые и все более и более углубляющиеся "экскурсии физиолога в область психиатрии". В своей статье "Пробная экскурсия физиолога в область психиатрии" Павлов (21) дал новое физиологическое объяснение кататонических симптомов шизофрении. Он считал, что кататоническая стадия шизофрении представляет по своему нервному механизму "хроническое гипнотическое торможение", предохраняющее ранимые корковые клетки слабой нервной системы больного от дальнейшего разрушения. Доказательства хронического гипнотического состояния у таких больных Иван Петрович видел в наличии парадоксальной фазы (на сильный голос не отвечают, а на шопот реагируют), негативизма (как проявления ультрапарадоксальной фазы), стереотипии в различных формах, эхолалии, эхопраксии, в развитии явлений каталепсии и кататонии и т. д. Развитие этого торможения имеет, по мнению Ивана Петровича "охранительное" значение. "Таким образом это состояние с одной стороны - патология, так как оно лишает пациента возможности нормальной Деятельности, с другой - по существу самого механизма есть еще физиология, физиологическая мера, потому, что оно предохраняет корковые клетки против угрожающего нарушения вследствие непосильной Работы" (стр. 442) (21). Здесь была блестяще представлена мысль о единстве физиологии и патологии.
Из изложенных физиологических соображений следовал прямой вывод практическое порядка о соответствующем содержании психически больных. Для таких больных, естественно, необходимы покой, тишина и самое деликатное обращение. "Следовательно, - заключал Иван Петрович - нужно как можно скорее, своевременнее как бы, переводить таких душевно-больных на положение больных, страдающих всякими другими болезнями, которые не истязают так непосредственно чувство человеческого, достоинства" (стр. 443) (21).
В дополнение к этой экскурсии сделаем еще одну, касающуюся физиологического анализа аменции (amentia)*. У больной, которую Иван Петрович видел в психиатрической клинике проф. П. А. Останкова, имелся симптом отождествления вновь встречающегося лица с кем-либо из давно известных ей людей. Таким образом уподобление у аментика превращалось в тождество. Может быть, мы имеем здесь две фазы: первую, когда больная только находит сходство, и вторую, когда происходит уже прямое отождествление одного лица с другим. Физиологическое объяснение Ивана Петровича сводилось к следующему. Большие полушария ребенка начинают свою работу с первичного синтеза. Анализ основан на применении коркового торможения. Может быть, аментики в силу своего заболевания переходят до некоторой степени в первобытное состояние ребенка: они воспринимают предметы, лица целиком, как дети, не в состоянии произвести различение в силу ослабления диференцировочных тормозных функций коры. К такому пониманию аменции близко подходил и Вернике.
* ("Среда", 4 июня 1930 г.)
Литература
1. Г. В. Скипин. Анализ высшей нервной деятельности собаки по усложненной секреторно-двигательной методике. Сообщение I. Труды физиологических лабораторий акад. И. П. Павлова, 1941, т. X, стр. 5.
2. Г. В. Скипин. То же. Сообщение II. Одно из условий для выявления ассоциации между двумя возбужденными участками коры больших полушарий собаки. Там же, 1941, стр. 17.
3. М. К. Петрова. Материалы к познанию физиологического механизма произвольных движений. Там же, 1941, стр. 41.
4. Н. А. Подкопаев. Выработка условного тормоза из проприоцептивного раздражителя. Труды физиологических лабораторий акад. И. П. Павлова, 1940, т. IX, стр. 400.
5. Г. В. Скипин. О характере действия положительного условного раздражителя в первые моменты его применения. Труды физиологических лабораторий акад. И. П. Павлова, 1941, т. X, стр. 172.
6. Г. В. Скипин. К вопросу о природе тормозного (угасательного) процесса, развивающегося в высших отделах центральной нервной системы собаки. Труды физиологических лабораторий акад. И. П. Павлова, 1940, т. IX, стр. 459.
7. П. К. Анохин. Соотношение процессов возбуждения и торможения при их одновременном протекании в коре больших полушарий. Труды IV Всесоюзного съезда физиологов, 1930, стр. 279.
7-а. П. К. Анохин. Соотношение между возбуждением и торможением при их одновременном протекании в коре больших полушарий. Труды физиологических лабораторий акад. И. П. Павлова, 1932, т. IV, стр. 322.
8. Г. В. Скипин. К вопросу об иррадиации и концентрации тормозного процесса. Там же, 1930, стр. 215.
8-а. Г. В. Скипин. То же. Труды физиологических лабораторий акад. И. П. Павлова, 1932, т. IV, стр. 318.
9. С. А. Петров. К вопросу о выработке диференцировки при коротком и длительном отставлении условного раздражителя. Труды физиологических лабораторий акад. И. П. Павлова, 1940, т. IX, стр. 148.
10. Ф. С. Павлов. К вопросу об образовании запаздывающих условных рефлексов: Там же, 1940, стр. 391.
11. Ф. П. Майоров. Опыт устранения гипнотического состояния у собак при помощи брома. Доклад в Обществе психиатров и невропатологов в Ленинграде, 22 ноября 1930 г. Обозрение психиатрии, неврологии и рефлексологии 1930, т. V, № 5.
11-а. Ф. П. Майоров. Устранение гипнотического состояния у собак при помощи брома. Труды физиологических лабораторий акад. И. П. Павлова, 1933, т. V, стр. 133.
12. В. В. Яковлева. Действие бромистого натрия на центральную нервную систему собаки возбудимого типа. Там же, 1933, стр. 97.
13. И. С. Розенталь. К влиянию различных доз бромистого натрия на высшую нервную деятельность. Там же, 1933, стр. 167.
14. И. О. Нарбутович. Действие сверхмаксимального раздражителя на нервную систему собаки уравновешенного сангвинического типа. Труды физиологических лабораторий акад. И. П. Павлова, 1938, т. VIII.
15. С. Н. Выржиковский и Ф. П. Майоров. Материалы к вопросу о влиянии воспитания на склад высшей нервной деятельности у собак. Доклад в Обществе физиологов 6 октября 1930. Труды физиологических лабораторий акад. И. П. Павлова, 1933, т. V, стр. 171.
16. И. С. Розенталь. К характеристике ориентировочных и оборонительных рефлексов. Архив биологических наук, 1930, т. XXX, вып. 1, стр. 119.
17. С. В. Клещов. Отношение звуков, как условно-рефлекторный раздражитель. Труды физиологических лабораторий акад. И. П. Павлова, 1933, т. V, стр. 213.
18. В. В. Строганов. Метод условных рефлексов в физиологии труда. Архив биологических наук, 1930, т. XXX, вып. 2, стр. 125.
19. И. П. Павлов. Краткий очерк высшей нервной деятельности. Статья в американском сборнике "Psychologies 1930". Двадцатилетний опыт, изд. 1932 г.
20. И. П. Павлов. Физиология и патология высшей нервной деятельности. Лекция врачам ГИДУВа, 12 января 1930 г., Брошюра. Медгиз, 1930.
21. И. П. Павлов. Пробная экскурсия физиолога в область психиатрии. Archives Internationales de Pharmacodynamic et de Therapie, 1930. Двадцатилетний опыт, изд. 1932 г.
При использовании материалов сайта активная ссылка обязательна:
http://anfiz.ru/ 'Анатомия и физиология человека'