Глава XXI - 1925
В первые годы работы Павлов и его школа придерживались той схемы условного рефлекса, которая была изложена в диссертации Бабкина 1904 г. (см. главу II). Согласно этому представлению, условно-рефлекторное замыкание происходит не прямо между корковым центром условного раздражителя и подкорковым пищевым центром, а через посредство коркового пищевого центра. В дальнейшем, в связи с экспериментальными опровержениями существования коркового пищевого центра Горшкова (1910 г.) и другими обстоятельствами, от указанного представления отказались. Стали полагать, что условно-рефлекторное замыкание осуществляется непосредственно между корковым центром условного раздражителя и подкорковым пищевым центром. Этот второй период продолжался примерно до 1925 г., когда в связи с новыми исследованиями Д. С. Фурсикова и М. Н. Юрман (1-5) опять вернулись к первоначальному взгляду. Они делали опыты на трех собаках, у которых было удалено одно полушарие головного мозга. Производилась выработка условного электрооборонительного рефлекса на звуковое раздражение с пораженной и здоровой стороны. Со здоровой стороны рефлекс был выработан, а с поврежденной это оказалось невозможным. С поврежденной стороны не удалось также получить и условных пищевых тактильных рефлексов. Такой отрицательный результат был и при повышении возбудимости введением стрихнина и кофеина: на неповрежденной стороне условные рефлексы были повышены, а на поврежденной стороне их не было. Расстройства этого рода не восстанавливались в течение длительного срока (на протяжении 9 месяцев после операции). При применении в качестве безусловного раздражителя очень сильного тока, вызывавшего иррадиированную оборонительную реакцию, вырабатывались "добавочные" условные рефлексы. Условная реакция, начинавшаяся в этих случаях с отдергивания неповрежденной лапы, могла затем проявляться и на "поврежденной" лапе, достигая в некоторых случаях большой интенсивности. Из приведенных данных авторы сделали заключение, что условно-рефлекторные замыкания осуществляются исключительно в коре головного мозга, что они происходят между воспринимающими корковыми центрами условного и безусловного раздражителей. С удалением полушария целиком удалялась и его моторная зона, почему авторы полагали, что "при выработке условного рефлекса требуется представительство в коре головного мозга не только для условного раздражителя, но и для безусловного".
Несомненно, из изложенных экспериментальных материалов следует вывод о том, что кора есть орган условных рефлексов; что же касается представления об условно-рефлекторном замыкании, то оно должно быть нами расценено не как непосредственный вывод из фактов, а скорее как их интерпретация с новой точки зрения. Эту интерпретацию надо считать правильной, она получила многочисленные подтверждения в дальнейших исследованиях. Такой точки зрения Павлов держался до конца своих дней, и мы до настоящего времени продолжаем думать так же.
Таким образом, в истории представлении Павлова и его школы схеме условно-рефлекторного замыкания мы отмечаем три следующих этапа (рис. 4).
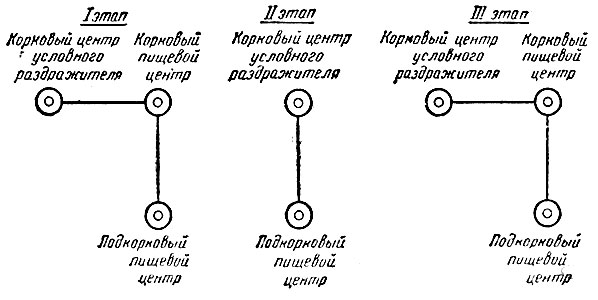
Рис. 4. Схема условного рефлекса
В связи с этим в работах Фурсикова и Юрман был поднят и принципиальный вопрос о корковых представительствах безусловных (врожденных) реакций вообще.
Для понимания механизма условного рефлекса существенно было выяснить: что делается в корковой клетке условного раздражителя во время применения безусловного? Разрешением этого вопроса было поручено заняться П. К. Анохину (6). Два выработанных условных раздражителя (свет в 16 свечей и сильный звонок) подвергались так называемому "покрытию", т. е. сначала давалась еда, а потом присоединялся условный раздражитель и продолжал действовать на фоне безусловного раздражения. После серии таких "покрытий" испытывались обычным порядком условные рефлексы на эти раздражители. Оказалось, что условный рефлекс на слабый раздражитель - свет - уменьшился, а на сильный - звонок - увеличился. Автор заключал, что во время действия безусловного раздражителя в клетке условного развивается торможение по принципу отрицательной индукции. Применение прочного условного раздражителя после начала безусловного ведет в борьбе возбуждения и торможения в корковой клетке условного раздражителя. Эта борьба противоположных нервных процессов вызывает "срыв", в случае света в сторону торможения, в случае звонка в сторону возбуждения. Поэтому в результате "покрытий" Уменьшился первый рефлекс и увеличился второй. Таким образом, было признано возможным "локальное нарушение функций корковых клеток", которое даже и при положительном действии вело к развитию, тормозного процесса в функционально поврежденных клетках и иррадиации его на соседние участки коры больших полушарий.
Этот вопрос, связанный с работой Крестовникова (см. главу XI), составил предмет ряда последующих исследований.
Как мы уже говорили, открытие и изучение явлений взаимной индукции привели к установлению принципа "функциональной мозаики", как принципа функциональной дробности в работе коры больших полушарий. Существование функциональной мозаики было доказано Сирягским на звуковом анализаторе (см. предыдущую главу). Этот принцип получил экспериментальное подтверждение и на примере кожного анализатора в опытах д-ра П. С. Купалова (7, 8, 9). Исследования производились им на чрезвычайно точно работавшей собаке сильного уравновешенного типа. На одной стороне тела у нее была выработана мозаика из 9 кололок, расположенных на равном расстоянии друг от друга, 5 из них были положительными пищевыми, а 4 - диференцировочными:
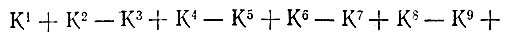
Положительные кололки всегда давали постоянный положительный эффект, а диференцировочные - нули.
Таким образом была образована "мозаика возбудимых и тормозных пунктов" в корковом кожном анализаторе, связанных между собой взаимной индукцией. При выработке такой системы условных раздражителей (применявшихся тогда на разных интервалах) оказалось, что стоило образовать только несколько чередующихся положительных и тормозных пунктов, как остальные стали действовать почти с первых же проб, как положительные и отрицательные, соответственно расположению. "Мы создали, - писал автор, - на определенном пространстве коры полушарий правильный ритм, и этот ритм захватывает и новые участки коры, на которые мы ничем извне непосредственно не воздействовали" (стр. 13) (8). Такое же явление наблюдал Сирятский на тоновой мозаике.
Из прежних исследований было известно, что работа с одними кожно-механическими раздражителями обычно способствовала развитию сна у подопытных животных. Опыты Купалова сопровождались полным отсутствием сна, что объяснялось тренированной мозаикой, содействовавшей концентрации торможения в отдельных функциональных очагах коры.
Далее, было установлено, что один и тот же положительный пункт, находящийся на определенном расстоянии от очага торможения, переживал периодическую смену состояний, то впадая в торможение, то переходя в состояние повышенной возбудимости, то снова впадая в торможение и т. д., и потом приходил в устойчивое нормальное состояние. При этом положительные пункты, расположенные на различном расстоянии от тормозного очага, испытывали прямо противоположные изменения. Такова была картина периодической смены возбудимости клеток коры в связи с механизмом взаимной индукции и последовательного торможения.
Были сделаны эксперименты для выяснения механизма взаимодействия тормозных и положительных пунктов в коре при функциональной мозаике (9). Положительные пункты кожной мозаики, примененные сразу же после тормозных, давали увеличенный эффект, т. е. на них сказывалась положительная индукция. Тормозные места, взятые после положительных (сразу и на разных интервалах), никогда не обнаруживали повышения возбудимости (отрицательная индукция). Применение положительных пунктов сразу после положительных (через 15 сек. их длительности) давало увеличенный эффект, т. е. наблюдалась иррадиация возбуждения и его суммация. При этих условиях эксперимента
было обнаружено отрицательной индукции, как в опытах Сирятного. После угашения одного положительного пункта оставалось последовательное торможение на других положительных пунктах, тормозные же нисколько не страдали.
Данные Купалова и Сирятского характеризуют не только функциональную дробность работы коры больших полушарий, но показывают также и взаимосвязь ее функциональных элементов. В этих опытах мы фактически уже имеем то, что потом вошло в содержание учения Павлова о "системности". В исследованиях Купалова каждый элемент представлял органическую часть целого. Им было показано, что практика одного пункта функциональной мозаики ведет к тренировке, укреплению всей мозаики в целом, как системы.
Как видно, в изложенных работах Купалова фигурирует павловская "теория корковых "пунктов", понимаемых здесь не только функционально, но и анатомически. И это имело под собой фактические основания.
Кроме того, в работах Купалова был снова поднят вопрос о числовом соотношении между возбуждением и торможением (см. работу Николаева в главе IX). Считалось необходимым установить "определенную пропорцию между числом применений в одном и том же опыте положительных и тормозных мест" (стр. 16) (8). Однако какой-либо определенной количественной закономерности установить не удалось, и исследователь действовал "чисто эмпирически"*.
* (Этим вопросом занимались, кроме Николаева, Крылов и Скипин.)
Волнообразный характер положительной индукции и последовательного торможения был также констатирован в опытах д-ра Н. А. Подкопаева (10), определявшего зависимость положительной фазы индукции от расстояния между положительным и тормозным пунктами кожной поверхности. У собаки на одной стороне тела были выработаны один условный пищевой рефлекс на кололку (на запястье передней лапы) и четыре диференцировки на кололки, расположенные на разных расстояниях от положительной (на середине передней ноги, сзади лопатки, в паху и на середине задней ноги). Испытывался положительный пункт через 0 сек. (практически через 0,5-1 сек.) после окончания действия того или иного тормозного раздражителя. В результате получилось, что процесс положительной индукции распределяется по кожному анализатору неравномерно. После применения крайних тормозных мест (самого ближнего и самого дальнего) обнаруживалась на положительной кололке положительная индукция (28,5 и 43%). После же применения средних диференцировок получался либо нормальный эффект, либо колеблющийся в ту и другую стороны.
Накопившийся материал наблюдений, касавшихся развития сна и гипнотизации у собак в экспериментальной обстановке, выдвинул необходимость систематического исследования вопроса о нервном механизме сна нормального и гипнотического, этому вопросу были посвящены опыты д-ра Б. Н. Бирмана (11, 12)*. Кроме обычной слюнной методики им была применена также графическая регистрация движений головы животного (по Рожанскому). У двух собак тон фисгармонии "до" - 256 был сделан условным пищевым раздражителем, а остальные 22 тона фисгармонии - диференцировками: вниз от 256 колебаний, до 85,3 и вверх до 768. У бодрых, нерасположенных ко сну животных можно было вызвать экспериментальным путем сон, применения одни тормозные тона. Последние вызывали сон тем глубже, длительнее было их действие и чем меньше промежутки времени между ними. При этом сон был глубже в промежутках между применениями тормозных раздражителей, чем во время их действия. При развитая торможения в нескольких анализаторах сон наступал быстрее. Но можно было образовать в коре и "сторожевой пункт", сопротивляющийся иррадиирующему внутреннему торможению. Этот пункт был связан с действием положительного тона 256, который будил собаку. Благодаря образованию такого "сторожевого пункта" в коре больших полушарий, сон мог быть достаточно глубоким, но он не был полным. Это был "сон с частичным бодрствованием". Для понимания нервного механизма сна надо иметь в виду две стороны его: глубину и полноту. Глубина обусловливается интенсивностью внутреннего торможения, а полнота - его экстенсивностью. Таким образом, удалось получить сон со "сторожевым пунктом". Автор считал, что этот "экспериментальный сон собак имеет черты, общие с гипнозом животных и человека, представляя собой промежуточную степень между тем и другим". Он наблюдал случаи, когда действие положительного тона на усыпленную собаку вызывало у нее каталептоидное состояние вследствие фиксирования торможения в двигательной области коры, при наличии условного секреторного эффекта. Диференцировочные тона, применявшиеся один за другим, способствовали возникновению тормозных очагов во многих местах и иррадиации торможения по всей коре, за исключением очага возбуждения, связанного с тоном 256. Но совпадая несколько раз с сонным состоянием, указанные диференцировочные тона превращались тем самым в условных гипнозогенных раздражителей. Вызванный таким образом сон осуществляется посредством того же механизма, что и условные "отрицательные" (тормозные) рефлексы, описанные Фольбортом (см. главу X). Данные Бирмана привели к заключению, что "гипноз и сон имеют в своей основе однородный нервный механизм - процесс внутреннего торможения", что "гипноз отличается от сна ограниченностью в распространении тормозного процесса", различные же фазы и формы гипноза обусловливаются "различием в распространении и локализации внутреннего торможения".
* (Предварительное сообщение было сделано еще в 1923 г.)
"Работа Б. Н. Бирмана, - писал Павлов в предисловии к его диссертации, - значительно приближает к окончательному решению вопрос о физиологическом механизме гипноза. Еще две-три добавочных черты - и в руках физиолога окажется весь этот механизм, так долго остававшийся загадочным, окруженным даже какою-то таинственностью" (12).
Изучение переходных состояний при развитии наркотического сна составило задачу опытов С. И. Лебединской (13). Она применяла хлоралгидрат и уретан на собаке сангвинического типа. Доза в 2 г обычно вызывала сон и была наиболее подходящей для наблюдения переходных состояний от бодрствования ко сну. Большие дозы не представляли такого удобства, так как переход ко сну происходил очень быстро, и, кроме того, затягивался период полного сна. Прослеживались изменения в величине условных пищевых рефлексов (на стук метронома, звонок, кожно-механическое раздражение, свет) и изменения в тормозном эффекте диференцировки (на более редкий стук метронома). Наркотики вводились клизмой перед опытом. Оказалось, что при усыпании животного происходило постепенное падение величины всех условных рефлексов, при этом слабые уменьшались больше и исчезали раньше, чем сильные. При переходе от сна к бодрствованию сильные раздражители начинали действовать раньше слабых. Таким образом и при засыпании и пробуждении наблюдалась наркотическая фаза. Что же касается диференцировки, то она оставалась полной, если только не было промежуточного слюнотечения. На этом основании автор полагал, что тормозная функция коры при применении таких доз хлоралгидрата и уретана сохраняется. Здесь не было отмечено установленного позже нарушения выработанного коркового торможения в начальных стадиях действия наркотиков.
Вопросу лечения экспериментальных неврозов у собак была посвящена работа М. К. Петровой (14). Лечился ранее описанный "срыв" у возбудимого "Пострела" и тормозимого "Милорда". Применялся бром с успехом у первого и без результата у второго. На основании наблюдений над "Пострелом" было сделано заключение; что бром действует, не понижая возбудимости, как это трактовалось в учебниках фармакологии, а восстанавливая и усиливая тормозной процесс и через это регулируя нервную деятельность. У "Милорда" же, у которого срыв произошел в сторону торможения, после полуторамесячного бесплодного применения бром был оставлен. Впоследствии было выяснено, что этот отрицательный результат зависел от ненадлежащей дозировки брома. Таким образом, в то время "Милорда" никакими средствами не удалось избавить от хронического патологического состояния. Следовательно, одни и те же приемы у собак с разными типами нервной системы вели к различным отклонениям от нормы, как в отношении их симптомов, так и в отношении их излечимости.
В опытах Н. В. Зимкина (15) срыв у подопытного животного произошел в сторону торможения с развитием хронического сонливого состояния в камере; при этом наблюдались уравнительная и парадоксальная фазы, как гипнотические фазы. Последние исчезали под влиянием подкожных инъекций 6-8 см3 2% раствора кофеина. Такой же эффект получался в результате применения прочной, концентрированной диференцировки. Однако механизм действия обоих средств был разный: в первом случае гипнотическое торможение устранялось повышением общей возбудимости коры, во втором - благодаря концентрации рассеянного торможения. В дальнейшем последний прием получил широкое применение.
Классическая "сшибка", описанная Разенковым (см. предыдущую главу), заключалась в непосредственном переходе от тормозного раздражителя к положительному. А. М. Павлова (16) применяла одновременно тормозный и положительный раздражители и таким образом изучала взаимодействие торможения и возбуждения при их одновременном возникновении в коре больших полушарий. На одной собаке со слабым тормозным процессом на протяжении 7 месяцев было произведено 9 проб одновременного применения диференцировочного метронома с разными положительными условными раздражителями (пищевыми): свисток, бульканье, кололка - на протяжении 30 сек. Эти испытания вызывали колебания величины условных рефлексов, преимущественно в сторону торможения. При повторении проб наблюдалось ослабление диференцировки, которая стала чаще растормаживаться. При одновременном возникновении двух противоположных процессов торможение имело наклонность иррадиировать еще во время своего действия.
Взаимодействие возбуждения и торможения получило также свое отражение на примере изучения динамики условной слюнной секреции (опыты П. С. Купалова) (17). Он производил регистрацию условной пищевой секреции за каждые 10 и 5 сек. в течение всего периода отставления (30 сек.). Нормальный тип условной секреции - это правильное прогрессивное нарастание эффекта, например:
при измерении по 10 сек.
при изменении по 5 сек.
Но динамика условной секреции не выражена всегда по одному типу. Помимо приведенного обычного хода секреции имеется другой с периодически меняющейся скоростью. Такая картина наблюдалась в процессе образования диференцировок. Экспериментатор сделал предположение, что "указанная периодичность бывает в том случае, когда в коре больших полушарий положительным пунктам противопоставлены тормозные. Колебания скорости условного слюноотделения указывают на периодическую смену состояний центров больших полушарий, на постоянную борьбу между возбуждением и торможением при установлении правильного баланса между этими процессами" (17).
Впоследствии Павлов пришел к обобщению, что всякая волнообразность в явлениях высшей нервной деятельности есть результат борьбы противоположных нервных процессов: возбуждения и торможения.
С основной идеи взаимодействия возбуждения и торможения начинает изложение своей работы Е. М. Крепс (18). "Из работ последнего времени можно сделать тот основной вывод, что состояние возбуждения коры и состояние торможения не представляются совершенно самостоятельными, независимыми, резко друг от друга отграниченными, но что между ними существует целый ряд переходных ступеней" (стр. 175) (18). Исследования Крепса заключались в определении влияния продолжительности отставления условного раздражителя на возбудимость больших полушарий. Условные рефлексы, отставленные на 30 сек., переводились на совпадающие и это вызывало очень сильное повышение возбудимости коры, даже у собаки тормозного типа. При этом растормаживались условный тормоз и диференцировка. Восстановление их являлось задачей большой трудности и у тормозной собаки требовало очень длительной тренировки. Изменения корковой возбудимости при переходе от отставленных рефлексов к совпадающим и обратно совершались очень быстро, в течение одного опыта. Переход на совпадающие рефлексы надо считать удобным методическим приемом для повышения тонуса коры. В согласии с предыдущими исследованиями Андреева, Валькова и Разенкова (см. главу XX) автор установил, что для нормального развития корковых тормозных процессов требуется некоторая средняя оптимальная степень возбудимости коры.
Как мы писали, уже давно стали применяться наряду с отдельными условными раздражителями всякие синтетические, комплексные. Важно было определить силу отдельных компонентов, входящих в сложный раздражитель. Это составило задачу опытов В. В. Яковлевой (19). Сначала были выработаны условные пищевые рефлексы отдельно на стук метронома, кололку и свет (слабый). Потом эти раздражители были соединены в комплекс и снова разъединены. На комплексный раздражитель условный рефлекс был больше, чем на отдельные раздражители. Величина условных рефлексов отдельных раздражителей после продолжительного совместного их применения в комплексе уменьшалась. Наиболее резко уменьшался рефлекс на слабый раздражитель - свет. Позднее Яковлевой же был выяснен механизм данного явления. Таким образом, путем разложения комплексного раздражителя можно определять силу отдельных раздражителей, входящих в него. В приведенном случае раздражители располагались по силе в следующем порядке: метроном, кололка, свет. При повышении корковой возбудимости введение нового условного раздражителя затрудняло развитие внутреннего торможения. Далее было констатировано, что при понижении возбудимости в одних анализаторах повышалась возбудимость в других. Как и в опытах М. К. Петровой, применение брома оказало не только благоприятное действие на выработку процессов внутреннего торможения, но в некоторых случаях и повышало величину условных рефлексов.
Вопросу угасания условных рефлексов были посвящены работы Подкопаева, Фролова и Виндельбанд и Анохина.
Н. А. Подкопаев (20) изучал влияние пищевого безусловного рефлекса на восстановление угашенного рефлекса на стук метронома тремя известными способами: 1) естественное восстановление временем, 2) восстановление посторонними раздражителями, в том числе и разнородными безусловными раздражителями с полости рта и 3) восстановление 'своим безусловным раздражителем (т. е. пищевым). Из полученных данных следовало, что нет никакой принципиальной, разницы между восстановлением угашенного условного рефлекса однородным безусловным раздражителем (еда) и разнородным безусловным раздражителем (соляная кислота - 0,5%). Автор считал, что свой безусловный раздражитель растормаживает, как и всякий другой любой растормаживающий агент, "по правилу внешнего торможения". Здесь мы встречаем последние отголоски старой концепции растормаживания как внешнего торможения торможения - концепции, уже сданной историей дела в архив.
Ю. П. Фролов и О. А. Виндельбанд (21) описали "особый вид" угасания. Были произведены эксперименты обычного угашения условного пищевого рефлекса на стук метронома. Получены волнообразные кривые процесса угасания. Эта волнообразность и составляла "особенность" угасания в данных опытах. Однако, как ясно из предыдущего, волнообразный характер присущ в той или иной степени всякой кривой угасания, что имело место в первых опытах угашения у Бабкина (см. главу II). Фролов и Виндельбанд наблюдали, что периоды низких цифр угашаемого рефлекса совпадали с наступлением сонливости животного; это состояние исчезало с появлением волны более высоких цифр. Здесь наблюдался один из многочисленных случаев перехода угасательного торможения в рассеянное, сонное. Когда чередовали опытные дни, в которые производились угашения, с днями, свободными от угашения, тогда указанная волнообразность сохранялась в течение довольно долгого времени. Если же ставили опыты с угашением несколько дней подряд, то получалась кривая угасания без выраженной волнообразности.
Вся картина волнообразного хода угасания заставляла предположить наличие положительной индукции в том же самом пункте коры. Это - самоиндукция. "Она играет, - по мнению автора, - крупную роль в установлении ритмических процессов, благодаря наличию которых становится возможным важнейшее явление отсчета времени в высших отделах центральной нервной системы животных".
В опытах П. К. Анохина (22) были выработаны условные пищевые рефлексы на стук метрономов частотою 40, 104 и 200 ударов в минуту. После 16 сочетаний рефлексы уравнялись по своему эффекту. Делалось острое прерывистое (через 3 мин.) угашение рефлекса на метроном 104, а затем через 3 мин. после одного нуля производились испытания метрономов 40 и 200 на вторичное угашение. В обоих случаях (М. 40 и М. 200) получался положительный эффект в 30-50% от обычной величины рефлекса. Таков факт. Объяснение же автора довольно спорно. Он рассуждал так: все три метронома адресуются к одной и той же корковой клетке, положительный эффект на М. 40 и М. 200 после угашенпя до нуля М. 104 объясняется растормаживанием клетки от новизны, получающейся при смене одной частоты метронома на другую. Дело, повидимому, заключается здесь не в новизне, которая должна была бы вызывать какую-либо ориентировочную реакцию и на это нет объективных оснований. Законно поставить вопрос и иначе: не почему М. 40 и М. 200 растормаживались, а почему они оказывались заторможенными? И на это можно ответить: потому, что возможно последовательное торможение в одной и той же корковой клетке, что и сказывалось при переходе от угашенной частоты метронома на неугашенную положительную. Последовательным торможением можно объяснить указанный факт и при предположении, что М. 40, М. 104 и М. 200 связаны не с одной корковой клеткой, а с несколькими.
Павлов сосредоточивал свое внимание и внимание своей школы на работе с собаками, как наиболее удобными экспериментальными объектами, позволяющими итти вглубь исследования динамики высшей нервной деятельности.
Но некоторые из его сотрудников расширили круг экспериментальных объектов и занялись изучением условных рефлексов у других видов животных: у рыб (Фролов), асцидий (Крепс), у птиц (И. А. Попов), низших обезьян (Фурсиков и др.), а также детей (Красногорский, Иванов-Смоленский и др.).
Ю. П. Фролов (23) произвел опыты на треске и бычке - рыбах, не имеющих коры больших полушарий: образовал условный двигательный электро-оборонительный рефлекс на свет и звук и получил угашение. По разработанной им методике он образовал у рыб условный оборонительный двигательный рефлекс на красный цвет и диференцировку на зеленый. Подкреплением служили индукционные удары электрического тока, пропускавшегося через воду аквариума (23а). Эти исследования были им продолжены и в дальнейшем.
Е. М. Крепсу (24) удалось получить нечто вроде условного рефлекса у асцидий. Индиферентный раздражитель (звонок), не вызывавший никакой двигательной реакции, после ряда сочетаний с механическим раздражением тела асцидий падающей каплей - сделался возбудителем защитного рефлекса. Исследователь считал, что "последнее может толковаться, как повышение возбудимости к данному раздражению или как указание на образование условного рефлекса. У форм прибивных и бедных реакциями между условной реакцией и простым помещением возбудимости к данному раздражению трудно провести резкую границу".
Таким образом был поставлен на очередь вопрос: как быть с законом условных рефлексов у низшие животных, не имеющих коры больших полушарий?
А. Г. Иванов-Смоленский (25) провел сравнительное изучение высшей нервной деятельности собаки и человека. На собаке опыты делались по слюнной пищевой методике и также по двигательной электро-оборонительной, на людях (студентах) - по "хватательной" методике автора. Задача заключалась в выработке условного рефлекса на синтетический (цепной) раздражитель и диференцировки на тот же раздражитель, только с перестановкой двух средних элементов ряда.
Положительный раздражитель: шум 1 сек. - пауза 1 сек. - низкий тон 1 сек. - пауза 1 сек. - высокий тон 1 сек. - пауза 1 сек. - звонок 1 сек. - перерыв 5 сек., повторение ряда 3 раза.
Диференцировочный раздражитель - то же самое с переставленными тонами: шум - высокий тон - низкий тон - звонок (с соответствующими паузами и повторениями).
В результате оказалось, что в отношении диференцирования таких синтетических раздражителей собака остается далеко позади человека. Задача, неразрешенная ею с помощью слюнных рефлексов, не могла быть решена и, с помощью двигательных оборонительных рефлексов. Выработка различения указанных синтетических раздражителей являлась для нервной системы собаки труднейшей задачей, вызвавшей явления "срыва".
Вначале новые представления Павлова о работе больших полушарий были "им оформлены в виде "теории условных рефлексов", но с самых первых опытов условные рефлексы изучались на базе безусловных и в их взаимодействии с безусловными, что было ясно видно из первых работ. Впоследствии систему своих взглядов Иван Петрович стал чаще именовать "учением о высшей нервной деятельности", понимая под высшей нервной деятельностью совокупность и взаимодействие Функций коры и подкорки, т. е. условных и безусловных рефлексов. Вся работа павловской школы по условным рефлексам осуществлена на основе пищевого и оборонительного инстинктов. Проблема инстинкта разрешалась Павловым в плане рефлекторной теории. По своему нервному механизму инстинкты суть сложные или сложнейшие безусловные Рефлексы. Систематическое изложение представлений павловской школы по этому вопросу было дано в книге Ю. П. Фролова: "Физиологическая природа инстинкта с точки зрения учения об условных и безусловных рефлексах" (26)*.
* (Первая статья автора "Современное состояние учения об инстинкте" была Напечатана в 1913 г. в Известиях Военно-медицинской академии.)
В ней были изложены история учения об инстинкте, история вопроса в павловской школе и представления школы о природе инстинкта, показана связь учения И. П. Павлова с учением Ч. Дарвина. В книге дана критика зоопсихологии и взглядов Русского биолога В. А. Вагнера, стоявшего на дуалистической позиции и в то время отрицательно относившегося к учению об условных рефлексах.
Труд Фролова мы рассматриваем лишь как установку для дальнейшего систематического анализа физиологического механизма инстинктов. Эта работа должна проводиться с точки зрения современных представлений о нейрогуморальной регуляции.
К 1925 г. относятся три работы И. П. Павлова, касавшиеся нескольких основных вопросов физиологии и патологии высшей нервной деятельности.
Фундамент всего учения - взаимоотношение возбуждения и торможения. Это было темой статьи Павлова: "Отношения между раздражением и торможением. Размежевание между раздражением и торможением и экспериментальные неврозы у собак" (27). Первое основное отношение между возбуждением и торможением - факт образования условных положительных и отрицательных связей. "Второй общий пункт отношения между раздражением и торможением в коре больших полушарий есть их взаимное пространственное разграничение, размежевание"*. "Очевиднейшую демонстрацию" этого Иван Петрович видел в вышеописанных опытах с механическим раздражением различных пунктов кожной поверхности. Таким образом "каждому процессу (т. е. возбуждению и торможению - Ф. М.) отводится свое место, свое время".
* (Курсив мой. - Ф. М.)
Здесь был подчеркнут именно момент "размежевания", раздельности возбуждения и торможения (в пространстве и во времени), а не их взаимодействие, внутренняя связь и взаимный переход друг в друга. Это соответствовало теории равновесия, которой старался придерживаться Павлов и которая учитывала только один момент внешнего баланса, количественного равновесия и неравновесия возбуждения и торможения. Дальнейшая логика фактов показала принципиальную недостаточность такой концепции.
В своей статье Павлов также изложил представление о "переходных состояниях" (или гипнотических фазах). Была намечена следующая градация состояний коры:
возбужденное состояние нормальное состояние (= состояние равновесия) уравнительное состояние парадоксальное состояние полное тормозное состояние.
Иван Петрович считал, что в конечном счете "настоящую теорию всех нервных явлений даст нам, только изучение физико-химического процесса, происходящего в нервной ткани" (стр. 372) (27).
"Мы делаем теперь уже 25-летний опыт понимать всю без остатка высшую нервную деятельность животного чисто физиологически, совершенно не пользуясь психологическими понятиями и терминами" - писал Павлов в своей работе о "здоровом и больном состоянии больших полушарий" (стр.372) (28). Тут были изложены не только основы физиологии высшей нервной деятельности, но и новые данные экспериментальной патологии высшей нервной деятельности. Иван Петрович указал на "высшую степень реактивности" и "стремительную функциональную разрушаемость" корковых нервных клеток. Было подчеркнуто физиологическое значение торможения, как "экономического процесса", ограничивающего "дальнейшее функциональное разрушение" и способствующего "восстановлению истраченного раздражимого вещества".
На основании накопившихся наблюдений поведения экспериментальных животных в норме и при отклонениях от нее впервые был ясно поставлен вопрос о типах нервных систем.
Первое же более или менее развернутое учение о типах было дано Павловым в его парижском докладе о "тормозном типе нервной системы собак" (29). Им была намечена такая классификация типов (в зависимости от преобладания возбуждения или торможения): сангвинический и меланхолический типы, как крайние, и вариации уравновешенного типа в середине. Таков был первоначальный несовершенный набросок.
Выступая тогда во французском психологическом обществе, Иван Петрович заявил: "Я убежден, что рано или поздно физиологи с их изучением нервной системы и психологи должны будут соединиться в тесной, дружной работе".
Литература
1. Д. С. Фурсиков и М. Н. Юрман. Условные рефлексы у собак без одного полушария. Архив биологических наук, 1925, т. XXV, вып. 4-5, стр. 147.
2. Д. С. Фурсиков и М. Н. Юрман. Значение коры головного мозга при образовании условных рефлексов. Медико-биологический журнал, 1925, вып. 1-2, стр. 117.
3. Д. С. Фурсиков. Последствия удаления коры одного полушария головного мозга у собак. Сообщение I. Методика. Общие наблюдения. Русский физиологический журнал, 1925, т. VIII, вып. 1-2, стр. 80.
4. Д. С. Фурсиков и М. Н. Юрман. Последствия экстирпации коры одного полушария. Сообщение II. Значение коры при выработке условных рефлексов. Там же, 1925, т. VIII, вып. 1-2, стр. 117.
5. Д. С. Фурсиков. Последствия экстирпации коры одного полушария. Сообщение III. О генерализации и выработке условных рефлексов на тактильное раздражение. Там же, 1925, т. VIII, вып. 5-6, стр. 231.
6. П. К. Анохин. Взаимодействие клеток условного и безусловного раздражителей в течение применения последнего. Доклад в Обществе физиологов 17 декабря 1925 г. Труды физиологических лабораторий акад. И. П. Павлова, 1927, т. II, вып. 1, стр. 107.
7. П. С. Купалов. Периодическая смена возбудимости клеток коры в связи с механизмом индукции и последовательного торможения. Доклад в Обществе физиологов 5 марта 1925 г. Реферат - Русский физиологический журнал, 1926, т. IX, вып. 1, стр. 130.
8. П. С. Купалов. Функциональная мозаика в кожном отделе коры головного мозга и ее влияние на ограничение сна. Доклад в Обществе физиологов 2 апреля 1925 г. Реферат - Русский физиологический журнал, 1926, т. IX, вып. 1, стр. 147. То же. Труды физиологических лабораторий акад. И. П. Павлова, 1929, т. III, вып. 2-3, стр. 3.
9. П. С. Купалов. О механизме взаимодействия тормозных и активных пунктов в коре больших полушарий при функциональной мозаике. Доклад в Обществе физиологов 9 апреля 1925 г., Реферат - Русский физиологический журнал, 1926, т. IX, вып. 1, стр. 155.
Тоже. Труды физиологических лабораторий акад. И. П. Павлова, 1929, т. III, вып. 2-3, стр. 27.
10. Н. А. Подкопаев. Зависимость положительной фазы индукции от расстояния между активным и тормозным пунктами. Доклад в Обществе физиологов, 5 марта 1925 г. Реферат - Русский физиологический журнал, 1926, т. IX, вып. 1, стр. 132.
10а. Н. А. Подкопаев. К вопросу о распределении положительной фазы индукции на различных пунктах кожного анализатора. Труды физиологических лабораторий акад. И. П. Павлова, 1940, т. IX, стр. 76.
11. Б. Н. Бирман. Экспериментальный подход к проблеме гипнотизма. Доклад в Обществе физиологов в декабре 1923 г. Реферат - Русский физиологический журнал, 1924, т. VII, стр. 362.
12. Б. Н. Бирма и Экспериментальный сон. Госиздат, 1925.
13. С. И. Лебединская. Действие хлоралгидрата и уретана на условные рефлексы (неопубликованная работа).
14. М. К. Петрова. Лечение экспериментальных неврозов у собак. Архив биологических наук, 1925, т. XXV, вып. 1-3, стр. 3.
15. Н. В. Зимкин. Отклонение от нормы баланса между возбуждением и торможением в коре больших полушарий и восстановление его под влиянием кофеина и диференцировок. Доклад в Обществе физиологов 17 декабря 1925 г. Реферат - Русский физиологический журнал, 1926, т. IX, вып. 1, стр. 176.
16. Л. М. Павлова. Взаимодействие тормозного и возбудительных процессов при их одновременном возникновении в коре больших полушарий. Труды физиологических лабораторий акад. И. П. Павлова, 1940, т. IX, стр. 119.
17. П. С. Купалов. Периодические колебания скорости условного слюноотделения. Доклад в Обществе физиологов 29 января 1925 г. Архив биологических наук, 1925, т. XXV, вып. 4-5, стр. 167.
18. Е. М. Крепс. О влиянии продолжительности отставления условного раздражителя на возбудимость больших полушарий. Доклад в Обществе физиологов 19 марта 1925 г., Архив биологических наук, т. XXV, вып. 4-5, стр. 175. 1
19. В. В. Яковлева. Опыт применения комплексного раздражителя. Доклад в Обществе физиологов 19 марта 1925 г. Реферат - Русский физиологический журнал, 1926, т. IX, вып. 1, стр. 139.
19а. В. В. Яковлева. Отдельные условные раздражители, продолжительно примененные в виде одновременного комплекса и затем снова разъединенные. Труды физиологических лабораторий акад. И. П. Павлова, 1927, т. II, вып. 1, стр. 37.
20. Н. А. Подкопаев. Влияние пищевого безусловного рефлекса на восстановление угашенного условного рефлекса. Доклад в Обществе физиологов 5 марта 1925 г. Реферат - Русский физиологический журнал, 1926, т. IX, вып. 1, стр. 133.
20а. Н. А. Подкопаев. Дальнейшие материалы к вопросу о восстановлении угашенного условного рефлекса своим безусловным. Труды физиологических лабораторий акад. И. П. Павлова, 1927, т. II, вып. 1, стр. 117.
21. Ю. П. Фролов и О. А. Виндельбанд. Особый вид угасания искусственного условного рефлекса. Архив биологических наук, 1925, т. XXV, вып. 4-5, стр. 153.
22. П. К. Анохин. Новизна, как особый раздражитель на примере растормаживания. Доклад в Обществе физиологов 5 марта 1925 г. Реферат - Русский физиологический журнал, 1926, т. IX, вып. 1. стр. 128. Труды физиологических лабораторий акад. И. П. Павлова, 1941, X, 163.
23. Ю. П. Фролов. Условные двигательные рефлексы у пресноводных и морских рыб. Труды физиологических лабораторий акад. И. П. Павлова, 1941, т. X, стр. 156.
23а. Ю. П. Фролов. О диференцировании световых условных раздражителей у рыб. Доклад в Обществе физиологов, 1925 г. Реферат - Русский физиологический журнал, 1926, т. IX, вып. I, стр. 113.
24. Е. М. Крепс. О реакции асцидий на внешние раздражения. Архив биологических наук, 1925, т. XXV, вып. 4-5, стр. 223.
25. А. Г. Иванов-Смоленский. Опыт сравнительного изучения высшей нервной деятельности человека и собаки. Медико-биологический журнал, 1925, № 3, 84.
26. Ю. П. Фролов. Физиологическая природа инстинкта с точки зрения учения об условных и безусловных рефлексах. Изд. "Время", 1925.
27. И. П. Павлов. Отношения между раздражением и торможением. Размежевание между раздражением и торможением и экспериментальные неврозы у собак. Статья на немецком языке в Scand. Archiv fur Physiologie, 1925; Двадцатилетний опыт, изд. 1932 г.
28. И. П. Павлов. Здоровое и больное состояние больших полушарий. Сообщение на французском языке в Сорбонне, декабрь 1925 г. Двадцатилетний опыт, изд.
29. И. П. Павлов. Тормозной тип нервной системы собак. Доклад на французском языке в Парижском психологическом обществе, декабрь 1925 г. Двадцатилетний опыт, изд. 1932 г.
30. И. П. Павлов. Предисловие к русскому переводу книги Цур-Штрассена "Поведение человека и животных в новом освещении" (перевод с немецкого языка Подкопаева. Изд. "Время", 1925).
При использовании материалов сайта активная ссылка обязательна:
http://anfiz.ru/ 'Анатомия и физиология человека'