Глава XX - 1924
Выход в свет 1-го выпуска I тома Трудов физиологических лабораторий акад. И. П. Павлова и Сборника, посвященного 75-летию акад. И. П. Павлова, дал возможность опубликовать сразу большую группу работ, свидетельствовавших о дальнейшем успешном развертывании исследований высшей нервной деятельности. Укажем на следующие основные вопросы этих работ: 1) проблема возбуждения и торможения, 2) экспериментальные неврозы, 3) типы нервной системы, 4) движение нервных процессов и взаимная индукция, 5) функциональная мозаика коры больших полушарий, 6) вопрос об относительной силе разных видов внутреннего торможения, 7) вопрос о старческих изменениях корковой деятельности, 8) образование условного рефлекса на раздражитель через кровь, 9) подтверждение резонаторной теории Гельмгольца, 10) вопрос о парной работе больших полушарий, 11) перенос экспериментальных данных на человека.
Анализ соотношения процессов возбуждения и торможения с точки зрения теории равновесия дал в своих исследованиях Д. С. Фурсиков (1). Данные его предварительного сообщения были нами изложены в главе XVII, остановимся здесь только на основных фактах и принципиальной стороне вопроса. Производились опыты с условными тормозами и устанавливалась зависимость между физической силой тормозного агента и выработкой условного тормоза. Было доказано, что для образования условного тормоза требуется известное соотношение в силе условного раздражителя и тормозного агента.
При физически слабом тормозном агенте, вызывающем незначительную ориентировочную реакцию, условный тормоз вырабатывается относительно медленнее, торможение при этом непостоянное и неабсолютное. При физически сильном тормозном агенте с резкой ориентировочной реакцией условный тормоз вырабатывается быстрее, торможение при этом постоянное и абсолютное. Раздражитель одной и той же интенсивности (напр., t 45° С) может быть вполне достаточным для выработки условного оборонительного рефлекса и совершенно недостаточным для выработки из него абсолютного условного тормоза к пищевому рефлексу на стук метронома. Было также отмечено, что один и тот же тормозной агент оказывает различное тормозящее действие на условные рефлексы различной силы.
Другой ряд фактов относится к установлению зависимости образования условного тормоза от интервала между тормозным агентом и условным раздражителем. Была дана следующая схема последовательности различных значений тормозного агента в зависимости от длины паузы:
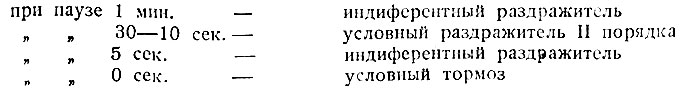
При паузе в 5 сек. прибавляемый агент теряет свойства условного раздражителя II порядка, но и не делается тормозным, т. е. в этом случае получается "уравновешенность возбуждения и торможения". При непосредственном переходе от тормозного агента к условному раздражителю удается выработать условный тормоз. Выработанный таким образом условный тормоз сохраняется и при постепенном удлинении паузы до 1 мин. Оказалось также, что применение тормозного агента через 3-5 сек. после начала действия положительного раздражителя не мешает выработке условного тормоза.
Почему же в одном случае образуется условный тормоз, в другом получается условный рефлекс II порядка, а в третьем нет ни того, ни другого? Автор объяснял это равновесием возбуждения и торможения и ставил в связь с большим или меньшим совпадением во времени следов первого и второго раздражителей. Однако он считал, что "несмотря на громадное преимущество метода условных рефлексов, которым уже более двадцати лет пользуется школа проф. И. П. Павлова, вопрос о соотношении процессов возбуждения и торможения в коре головного мозга Остается до сих пор самым темным. Очевидно, ближайшей задачей является накопление фактического материала, имеющего отношение к данному вопросу" (стр. 4) (1).
Дальнейшая история дела показала, что одно эмпирическое накопление даже и бесчисленного материала не могло разрешить и не разрешило этого основного вопроса.
Нарушения нормального равновесия возбуждения и торможения были исследованы И. П. Разенковым, А. Д. Сперанским и М. К. Петровой. С их работ начинается систематическое изучение экспериментальных неврозов.
Д-р И. П. Разенков (2) изучал изменения раздражительного процесса коры полушарий головного мозга собаки при трудных условиях*. Им была осуществлена классическая "сшибка" (квалифицированная в работе, как "трудная задача") и в результате "сшибки", столкновения возбуждения и торможения получен "срыв" нормальной высшей нервной деятельности (обозначенный им, как "нарушение деятельности полушарий головного мозга"). "Сшибка" заключалась в непосредственном переходе от тормозной (диференцировочной) кололки (частотою 12 прикосновений в минуту) к положительной, пищевой (частотою 24 прикосновения в минуту) на одном и том же месте. При такой очень быстрой смене одного процесса другим, противоположным, произошло нарушение равновесия возбуждения и торможения и отклонение от нормы. В течение некоторого времени после этого наблюдались четыре последовательные стадии изменений в соотношении условных рефлексов: 1-я стадия - "тормозная" (отсутствие всех положительных рефлексов), 2-я - "парадоксальная" (слабые условные раздражители вызывали больший эффект, чем сильные), 3-я стадия характеризовалась тем, что на все "условные раздражители получались одинаковые по своей величине условные рефлексы" (т. е. то, что потом стали называть "уравнительной" фазой), и 4-я стадия - "промежуточная к норме", когда "максимальные условные рефлексы получались от раздражителей средней силы". В таком порядке наблюдался постепенный по дням переход нервной системы собаки от тормозного "срыва" к норме. Впоследствии эти стадии были многократно установлены и при других обстоятельствах и вошли в содержание ученая Павлова о гипнотических фазах. Тормозную, парадоксальную и уравнительную фазы видели очень многие экспериментаторы. Что же касается "промежуточной" стадии то ее, кроме Разенкова, имели в своих опытах только В. В. Ринкман**, И. С. Розенталь*** и П. О. Макаров****. В опытах Рикмана был случай, когда не действовали ни сильные, ни слабые раздражители, а положительный эффект давали лишь раздражители средней силы. Аналогичные явления наблюдал и Розенталь. Открытый Разенковым факт надо считать доказанным. Однако ввиду его чрезвычайной редкости он не вошел в систему павловских гипнотических фаз.
* (Доложено на II съезде психиатров и невропатологов, 7 января 1924 г.)
** (См. Труды Физиологических лабораторий акад. И. П. Павлова, 1928, т. III, вып. 1, стр. 24 и 30 (оп. № 156).)
*** (См. Архив биологических наук, 1932, т. 32, вып. 3, стр. 188.)
**** (См. Труды Физиологических лабораторий им. И. П. Павлова, 1940, т. IX, стр. 112.)
Указанная трудная задача была повторена четыре раза и потом еще раз уже с дозированными по силе раздражителями, чтобы лучше выявить фазовые состояния (сильный и слабый свисток, сильный и слабый свет). При этом оказалось, что повторение "сшибок" вызывало некоторую тренировку, упражнение нервной системы и вело к постепенному ослаблению и исчезновению нарушений высшей нервной деятельности. Таким образом данные эксперименты показали и важное значение тренировки нервной системы.
"Нарушение деятельности полушарий головного мозга при трудных условиях" автор рассматривал как "результат нарушения баланса между возбуждением и торможением".
В заключение он имел основание указать на значение его работы для педагогики, учения о нервных и душевных болезнях и жизни вообще.
В опытах Разенкова в яркой форме был показан "закон силы" .(см. опыт № 68 - стр. 104, опыт № 226 - стр. 112 и др.).
Приведем для иллюстрации один протокол:

Как видно из протокола, физически сильный раздражитель вызывает больший условный рефлекс, чем слабый раздражитель.
В работе Разенкова было дано вполне обоснованное сопоставление выше описанных стадий изменения высшей нервной деятельности с тремя стадиями парабиоза Н. Е. Введенского.

Здесь мы имеем не только одинаковую терминологию или внешнюю аналогию, но и родство явлений по существу. Явления, наблюдавшиеся Введенским на нерве, были обнаружены Разенковым, а впоследствии и многими другими сотрудниками павловской школы, в деятельности корковых центров. Таким образом в опытах по условным рефлексам получила подтверждение мысль Введенского о приложимости открытого им закона парабиоза к нервным центрам.
Систематическая разработка учения об экспериментальных неврозах была стимулирована большим ленинградским наводнением осенью 1924 г. Территория Института экспериментальной медицины была затоплена водой, уровень которой достигал метра, и экспериментальных собак пришлось спасать из затопленного собачника. В результате пережитого потрясения у многих собак потом наблюдались отклонения от нормальной деятельности: у одних в сторону преобладания возбуждения с нарушением всех тормозов, у других, наоборот, было общее заторможенное состояние с исчезновением всех положительных условных рефлексов. Изменение взаимоотношений процессов возбуждения и торможения у собаки после наводнения в то время наблюдал д-р А. Д. Сперанский (3). У его "Августа" сильный раздражитель (наводнение) вызвал длительное нарушение равновесия возбудительного и тормозного процессов с преобладанием последнего, нарушил правильную реакцию животного, не только количественно, но и качественно изменив самый характер реакции: вместо обычной пищевой была оборонительная (с отказом от еды). Все поведение собаки характеризовалось "резко выраженной пассивного типа оборонительной реакцией". По своему внешнему проявлению оно было близко к состоянию травматического невроза или невроза страха у людей. После того, как собака пришла к нормальному состоянию, были сделаны опыты с искусственным наводнением: в комнату, где находилась в станке собака, пускали струю воды. В результате тоже получили "срыв" высшей нервной деятельности в сторону торможения. Для восстановления нормы благотворное действие имел "социальный раздражитель", т. е. присутствие самого экспериментатора около собаки во время опыта. Потом достаточно было для успокоения животного оставлять в камере только пиджак экспериментатора, т. е. действовать одной лишь частью сложного "социального раздражителя". Тогда по характеру "срыва" был определен тип нервной системы "Августа" как слабый, тормозный. Мы еще вернемся в дальнейшем к этому вопросу, ввиду его принципиального значения.
М. К. Петрова (4) получила "срыв" у двух собак разного типа ("Милорда" и "Пострела") в результате особо трудных условий эксперимента. Вырабатывались все четыре вида внутреннего торможения: запаздывание 6 условных пищевых рефлексов на 3 мин., диференцировка к ним продолжительностью 3 мин., условный тормоз на 3 мин. и угасание. Труднее всего вырабатывался у обеих собак условный тормоз. Поставленная задача собаками была выполнена, но с трудом и каждой по-разному. У обеих собак ее решение сопровождалось общим возбуждением, сильнее выраженным у "Пострела". Но, в конце концов, "Милорд" обнаружил преобладание тормозного процесса, а "Пострел" - преобладание возбуждения. В зависимости от разного типа Нервной системы этих собак и срыв у них получился в разные стороны.
Ранее было установлено, что удлинение отставления (т. е. периода изолированного действия условного раздражителя) может вести, вследствие иррадиации торможения, к сонливости. Однако в результате тренировки запаздывания (на 1 мин. и 3 мин.) сонливость может и не развиваться, как это было в приведенных опытах Петровой, а также В. М. Архангельского, работавшего в то же время. В этом случае, очевидно, имеет место развитие концентрации торможения.
При работе М. К. Петровой (5) с "живым" "Пострелом" и "спокойным" "Милордом" оба они впали в сонное состояние, благодаря однообразному применению одного и того же условного раздражителя - стука метронома. Они были выведены из этого положения выработкой других пищевых условных рефлексов. Затем, собаки снова были вогнаны в сон в процессе образования из всех 6 раздражителей, запаздывающих на 3 мин. Этот сон удалось устранить применением короткого отставления всех раздражителей на 5 сек. У "Пострела" выработка запаздывающих рефлексов сопровождалась некоторое время общим возбуждением. Таким образом, еще раз было доказано, что сон есть иррадиированное внутреннее торможение, что способ разнообразных и коротко-отставленных условных раздражителей оказался действительным в смысле поддержания бодрого состояния у обеих собак, несмотря на противоположный характер их типа нервной системы; практика же локализации торможения в виде постепенного отставления до 3 мин. имела благотворное влияние на "Пострела" в смысле уравновешивания его нервной системы и выполнения предложенной жизненной задачи.
К вопросу о типах примыкает также исследование д-ра Е. М. Крепса. (6), представившего опыт индивидуальной характеристики экспериментального животного. В начале его работы говорится, что данная собака "подходит под тип "солидных" собак". Однако приведенное описание соответствует слабому, тормозному типу. Правда, автор стремился скорее к физиологическому описанию типа, чем к его квалификации, как такового. Недостаточно ясно было в то время и представление о "солидном" (т. е. флегматическом) типе собаки (ввиду его сравнительной редкости). Собаке Крепса плохо давались все виды внутреннего торможения. Но характерно, что в результате тренировки и после летнего перерыва это торможение улучшилось. Это животное - по мнению исследователя - представляло тип нервной системы с низкой возбудимостью, со слабостью раздражительного процесса. Вследствие постепенного приближения частоты ударов положительного метронома (76) к частоте диференцировочного (М-144)* у собаки произошел "срыв" в сторону торможения. Благодаря применению брома (2% раствора NaBr 100 см3 per rectum) нервная деятельность была урегулирована: улучшился как процесс торможения, так и возбуждения.
* (Это - тема Шенгер-Крестовниковой, Усиевича и Фурсикова, только поставленная наоборот. Следовало бы сделать систематическое исследование взаимодействия возбуждения и торможения именно при такой вариации опытов, когда производится постепенное сближение М+ и М-, идя не от М-, как это делали, а от М+ к М-.)
Автор писал о двух возможных видах "срыва": в сторону возбуждения и в сторону торможения. В причинах преобладания того или другого процесса при этом "повидимому, главную роль играют особенности каждой нервной системы" (стр. 129) (6).
Дальнейшие исследования типов нервной системы шли в тесной связи с изучением экспериментальных неврозов и экспериментальной патологии высшей нервной деятельности вообще. В первое время и то и другое основывалось на количественном представлении о равновесии возбуждения и торможения и нарушении этого равновесия.
К вопросу о движении нервных процессов в коре - относятся работы Н. А. Подкопаева, А. Г. Иванова-Смоленского и Е. М. Крепса.
Д-р Н. А. Подкопаев (7) разрешал задачу "в каком состоянии находятся пункты данного анализатора в то время, когда в каком-нибудь другом пункте этого же анализатора происходит процесс торможения" (стр. 297) (7). У двух собак был выработан прочный условный пищевой рефлекс на кожно-механическое раздражение различных участков кожи вдоль одной стороны тела. Производились опыты с острым прерывистым (через 2 мин.) угашением какой-нибудь кололки до нуля. Получив нуль эффекта за 30 сек. изолированного действия угашенной кололки, автор присоединял на 30-й секунде раздражение другой кололки (то более дальней, то ближайшей к угашенному пункту), не прекращая действия угашенной кололки. Каждое испытание занимало 1 мин.: первые 30 сек. действовала одна угашенная кололка, вторые 30 сек. продолжалось действие угашенной совместно с какой-нибудь другой положительной кололкой. Был подтвержден факт Когана (см. главу XII) относительно того, что концентрация угасательного торможения происходит медленнее иррадиации. В опытах Подкопаева время концентрации было в 35-40 раз больше времени иррадиации; у Когана же это отношение было 1:4. В результате своих экспериментов Подкопаев пришел к заключению, что "процесс внутреннего торможения, возникший в коре больших полушарий под влиянием тормозного условного раздражителя, остается строго концентрированным в очаге своего возникновения во все время, пока действует этот тормозной раздражитель". Во время действия тормозного агента на периферии очага торможения имеется положительная индукция. Возникающий в коре очаг внутреннего торможения концентрирует на себя все те следы внутреннего торможения, которые имеются налицо в данном анализаторе к моменту возникновения этого очага. Иррадиация же внутреннего торможения наступает лишь по окончании действия тормозного раздражителя и происходит очень быстро. Это положение в Дальнейшем укрепилось, как основное, и по отношению к другим выработанным тормозам (диференцировке и условному тормозу). Оно верно, но его. нельзя считать абсолютным, так как иррадиация внутреннего торможения иногда может происходить и во время действия тормозного раздражителя, что зависит от целого ряда обстоятельств: длительности тормозного раздражителя, его характера и физической интенсивности, типа нервной системы животного и др. И в работе Подкопаева мы находим указание на существование небольшого торможения (12-18%) положительной кололки во время применения угашенной, хотя им утверждается, что "во время действия тормозного раздражителя процесс торможения держится в пункте своего возникновения, не иррадиируя". Автор объяснял это тем, что "возбуждение с данного пункта каждого анализатора должно по пути к слюнной железе пройти через пищевой центр. А этот последний находится в это время всегда в состоянии торможения, вследствие ряда предшествующих угашений" (стр. 301) (7). Почему же тогда этого эффекта торможения нет, когда условные раздражители применялись им же не на фоне угашения непосредственно?
В исследованиях Подкопаева отчетливо выступала волнообразность угасательного торможения, "напоминающая по форме кривую затухающего гармонического колебательного движения" (стр. 299) (7). Эту волнообразность наблюдали также Крепе, Фролов и Виндельбанд. Ока была установлена еще в самых первых опытах угашения, производившихся Бабкиным (см. главу II). Волнообразность характерна и для других корковых процессов. Так, Сирятокий в тот же период (1924 г.) указал на волнообразный характер иррадиации и концентрации угасательного торможения.
Можно сказать, что все корковые нервные процессы протекают не линейно, а волнообразно.
Исходя из павловского положения о том, что звуковой анализатор есть тоже корковая проекция периферического органа слуха д-р А. Г. Иванов-Смоленский (8) сделал опыты угашения звуковых раздражителей. Им были выработаны условные пищевые рефлексы на 4 тона разной высоты, на шум и стук метронома. Подобно тому, как в кожном анализаторе, здесь тоже наблюдались явления иррадиации и концентрации торможения. Делалось обычное угашение какого-либо одного раздражителя и после этого на разных интервалах испытывались другие. На тонах, резко отличавшихся по высоте от угашаемого, торможение сказывалось позднее и держалось меньше, чем на тонах, по высоте близких к нему. Торможение, возникавшее при угашений условного рефлекса на какой-либо тон, отзывалось на шумах и стуках значительно слабее, чем на тонах. Эти последние как бы были окружены по периферии шумами и стуками. Угашение рефлекса на шум или стук давало более сильную волну торможения в сторону тонов, чем обратно. Те же явления автор наблюдал и на людях, здоровых (у которых они протекали быстрее, чем у собак) и психически больных.
Уже в то время ясно выступила связь явлений иррадиации и концентрации с явлениями взаимной индукции. Это можно было видеть и в данных д-ра Е. М. Крепса (9). Он изучал последействие диференцировочной кололки на пяти положительных кололках, расположенных на расстоянии 10, 20, 40, 60 и 90 см от первой. В начале опытов он получил развитие сонливости вследствие монотонного применения одних кожно-механических раздражений. Сонливость удалось устранить при помощи других условных раздражителей. По мере упрочения диференцировки наблюдалось укорочение периода концентрации торможения. В связи с концентрированием диференцировочного торможения выявилась положительная индукция на положительных пунктах после паузы в 0 сек. и 5 сек. после диференцировки. На паузах в 15 сек., 30 сек., мин., 2, 3 и 5 мин. - сначала имела место фаза иррадиации торможения с его максимумом на 30-й сек., потом постепенная концентрация торможения, заканчивавшаяся после 5 мин. Результаты Крепса несколько расходились с данными Когана (см. главу XII): не было постепенного охватывания торможением сначала ближних мест, потом дальних, как в опытах последнего. Некоторая разница получилась и с опытами Красногорского (см. главу IX). Факты Крепса ближе совпадали с данными Анрепа (см. главу XV). В этих разногласиях, по мнению Крепса, "могла играть роль индивидуальность собаки, а затем и различная методика" (стр. 328) (9).
Последовательная положительная индукция была описана д-ром Ю. П. Фроловым (10), проводившим наблюдения на одной собаке, служившей для изучения желудочного пищеварения. У нее возникали взрывы возбуждения после длительного стояния в станке - явление "нервной самоиндукции". Это обусловливалось еще и тем, что собака отличалась пассивно-оборонительные рефлексом и, следовательно, наклонностью к затормаживаемости. Оказалось, что чем период стояния ее в станке был дольше, тем реакция возбуждения наступала быстрее.
Подобные явления могут наблюдаться и у людей, особенно у детей (например, в школах).
Влияние положительного раздражителя на развитие отрицательной индукции было показано в опытах И. Р. Пророкова и И. Н. Журавлева.
И. Р. Пророков (11) установил, что для развития отрицательной индукции наиболее благоприятным является предшествование отрицательному (диференцировке) положительного раздражителя. На двух собаках получился следующий одинаковый результат: диференцировочный метроном после положительного метронома не давал, растормаживания, даже и при введении 0,05 чистого кофеина и при однодневном голодании; тот же диференцировочный метроном, примененный после положительного звонка или тона, чаще растормаживался и, если растормаживание с течением времени пропадало, то оно снова обнаруживалось при даче 0,05 кофеина и при однодневном голодании. Таким образом сохранению тормозного действия диференцировки в первом случае способствовала отрицательная индукция от положительного метронома.
Аналогичный факт был и в экспериментах д-ра И. Н. Журавлева (12). Он работал на собаке с удаленными gyrus coronariusi и g. ectosylvius с одной стороны. Наряду с разными условными пищевыми раздражителями практиковались положительный комплексный раздражитель из 3 кололок (применявшихся одна за одной по 5 сек. с интервалом 3 сек.) и диференцировочный комплекс из тех же кололок в обратном порядке. Опыты ставились на больной и здоровой стороне. Такая диференцировка могла быть выработана при условии предшествования ей положительного комплекса кололок, что свидетельствовало о влиянии отрицательной индукции. Тормозной эффект диференцировки был больше, если она применялась после положительного комплекса кололок, чем если она бралась после других положительных раздражителей.
Явление взаимной индукции сказалось в яркой форме в исследованиях д-ра В. В. Сирятского (13) с "функциональной мозаикой"*. На одной собаке была образована тоновая (от фисгармонии) мозаика положительных и тормозных пунктов на одинаковых интервалах в 3 тона, на другой-то же на нервных интервалах (1, 11/2, 2 и т. д. тона). Благодаря такой функциональной мозаике в звуковом анализаторе были обнаружены четыре вида взаимной индукции между возбудимыми и тормозными пунктами коры (рис. 3).
* (Доложено на Всероссийском съезде психиатров и невропатологов 7 января 1942 г.)
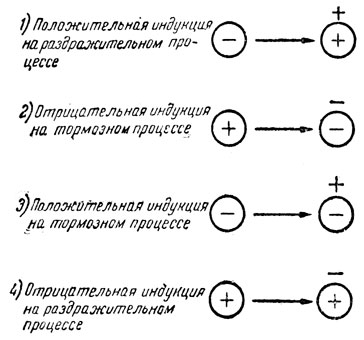
Рис. 3. Взаимная индукция
При образовании указанной функциональной мозаики из положительных и тормозных (диференцировочных) звуков Сирятский (14) получил патологические отклонения в высшей нервной деятельности, вследствие "трудного балансирования процессов возбуждения и торможения". Характер развившегося неврастенического состояния зависел от типа нервной системы: у возбудимого типа - сдвиг в сторону возбуждения, у тормозного - в сторону торможения. Для лечения собак были использованы бром (2,0), tinctura Valerianae и даже алкоголь в малых дозах.
Аналогичные явления автор наблюдал у 8-летнего ребенка при выработке у него функциональной мозаики на кожном анализаторе.
Уже давно назрела необходимость разрешить вопрос об относительной силе разных видов внутреннего торможения. Этим и занялся д-р В. М. Архангельский (15). Он ставил опыты на одной собаке с резко выраженной двигательной реакцией. Изменения в соотношении секреторной и двигательной реакции решено было использовать в качестве критерия силы разных видов внутреннего торможения. Наиболее сильным в этих опытах оказалось диференцировочное торможение: им тормозилась как слюнная, тал и двигательная реакция. Самым слабым было запаздывательное торможение: им задерживалась слюнная реакция, но не двигательная. Условное торможение занимало среднее место: оно тормозило, хотя и с трудом, как секреторную, так и двигательную реакции.
Опытами Архангельского не завершилось исследование данного вопроса: относящийся к нему материал мы найдем и в дальнейших работах.
Старческие изменения высшей, нервной деятельности изучались д-ром Л. А. Андреевым (16). Он собрал ценный материал в опытах на одной старой собаке. Старческие изменения организма вызвали у нее понижение возбудимости коры больших полушарий, вследствие чего условные слюнные рефлексы на почве пищевого, как довольно слабого раздражителя, не могли быть выработаны. Но возможно было образовать условные оборонительные рефлексы при подкреплении 0,25% раствором соляной кислоты. Одновременно с ослаблением процессов возбуждения с еще большей резкостью выступили расстройства торможения, что сказалось в невозможности выработки даже грубой диференцировки и в ослаблении угасательного торможения. Весьма далеко зашедший старческий маразм нервной системы обусловил последовательное исчезание слюнных рефлексов, в первую очередь искусственных, затем натуральных. По мнению исследователя, dementia senilis и старческий распад высшей нервной деятельности собаки имеют не только общие анатомо-гистологические изменения атрофического характера, но также и функциональные расстройства. Было констатировано совпадение картины угасания жизнедеятельности коры больших полушарий при старении с данными, полученными ранее Розенталем и Фроловым на собаках при голодании (см. главу XVIII).
Общие результаты исследований Андреева соответствовали тому, что получил д-р А. В. Вальков (17) при изучении высшей нервной деятельности у щенка с удаленной щитовидной железой. Удаление было сделано в возрасте 6 недель. Работа началась через 10 месяцев после этого. Другой щенок был контрольным. Условный пищевой рефлекс у тиреоидектомированной собаки вырабатывался с большим трудом и не достигал достаточной прочности. Его приходилось подкреплять от 2 до 4 раз, чтобы он достиг своей обычной величины. Условный, кислотный рефлекс вырабатывался и сделался прочным, но диференцировку к нему образовать не удалось. Однако удалось выработать диференцировку на условном электро-кожном рефлексе, при этом была необходима известная интенсивность тока. Таким образом был установлен важный факт зависимости коркового торможения от уровня возбудимости коры, от ее тонуса. "Единственный вывод, к которому мы, - писал автор, - позволяем себе притти, заключается в том, что процессы возбуждения и торможения в коре больших полушарий находятся в какой-то зависимости друг от друга" (стр. 367).
Указанной факт наблюдал также И. П. Разенков (18) на собаке с ослабленной корой больших полушарий, вследствие односторонней экстирпации gyri coronarius et ectosylvius. В основном он имел то же, что и Андреев и Вальков, т. е. выработать диференцировку удалось только на оборонительном раздражении, следовательно, при более высоком тонусе коры. "Значит, для проявления торможения, - считал он, - требуется определенное состояние возбуждения. Таким образом, между торможением и возбуждением устанавливается прямая функциональная зависимость. И такую функциональную зависимость мы понимаем не в математическом смысле, что всякое изменение возбуждения или торможения обязательно вызывает изменение торможения или возбуждения. Такую функциональную зависимость мы понимаем только таким образом, что для проявления торможения требуется определенное состояние возбуждения. Достаточно возбудимости мозга понизиться: ниже определенного состояния, как торможение не проявляется, точно также не проявляется оно и при очень высокой возбудимости мозга. Значит, для проявления тормозного процесса требуется определенный оптимум состояния возбудимости мозга" (стр. 145) (18).
Следовательно, данные Андреева, Валькова и Разенкова указывают на внутреннюю, интимную связь противоположных нервных процессов - возбуждения и торможения.
Кроме этого, в опытах на той же собаке Разенков наблюдал парадоксальную, уравнительную и промежуточную фазы, задерживающее влияние раздражения больных мест кожи, а также предсудорожных и судорожных состояний на условные рефлексы.
Д-р В. В. Строганов (19) изучал образование условного пищевого рефлекса и диференцировки на синтетический раздражитель. Условным раздражителем был сделан следующий комплекс: свет 1 сек.- пауза 1 сек. - кололка 1 сек. - пауза 1 сек. - бульканье 1 сек. - пауза 3 сек. и повторение данного ряда раздражителей 4 раза. Диференцировка вырабатывалась на тот же комплекс в обратном порядке. Образование условного рефлекса произошло легко. Автор полагал, что при этом происходит "образование сложного функционально-рассыпанного центра в коре больших полушарий, существенные элементы которого соответствуют компонентам раздражителя". Выработка же диференцировки являлась весьма трудной задачей, лежавшей на границе возможности больших полушарий, по крайней мере, данной собаки. Сложность происходившего нервного процесса была причиной крайней неустойчивости торможения, отчего легко растормаживалась диференцировка. С укреплением синтетической диференцировки торможение стало проявляться и на отдельных компонентах, захватывая сперва более слабые. Было обращено внимание на разное значение диференцировочных нулей: в одних случаях они сопровождались последовательным торможением, в других - положительной индукцией.
Особый род условных рефлексов на раздражитель через кровь (автоматический раздражитель) был описан д-ром В. А. Крыловым (20), а ранее его установлен Н. А. Подкопаевым*, образовавшим условный рвотный рефлекс на тон, покрепляемый апоморфином. Опыты Крылова были им начаты еще в Ташкенте на собаках с морфием, но по другому поводу. Он обратил внимание на то, что эффект действия этого яда не уменьшался при повторных его введениях. Непонятный факт объяснялся участием в этих явлениях условных рефлексов на самую процедуру введения морфия. Этот условный морфийный рефлекс, воспроизводивший в ослабленной степени симптомы морфийного отравления, был потом детально исследован Крыловым в лаборатории акад. Павлова. Было доказано угасание такого условного рефлекса путем нескольких повторных инъекций физиологического раствора. Была также получена диференцировка: Крылов делал инъекцию морфия в одной обстановке в заднюю ногу, другое лицо производило инъекции физиологического раствора в другой обстановке в переднюю ногу; в результате в первом случае приготовления к введению яда вызывали картину морфийного отравления, во втором нет. Так же были сделаны эксперименты с хлоралгидратом. После нескольких вливаний в прямую кишку 2-3 г, вызывавших у собаки сон, уже одни приготовления к вливанию стали вызывать сонное состояние.
* (В 1914 г.)
В приведенных опытах с морфием и хлоралгидратом автор усматривал некоторый "аналог внушения".
Исследование условных рефлексов на различные автоматические раздражители было потом продолжено рядом лиц.
"Едва ли можно оспаривать, что действительно все вопросы, составляющие доселе область так называемой физиологии органов чувств, могут разрабатываться на животных при помощи условных рефлексов" - утверждал Иван Петрович в "Лекциях о работе больших полушарий головного мозга" (стр. 132).
Как образец применения метода условных рефлексов в физиологии органов чувств можно привести работу Л. А. Андреева (21), представившего новые доказательства резонаторной теории слуха Гельмгольца. Почти три года проводились опыты на одной собаке, у которой были разрушены частично кортиевы органы с обеих сторон; разрушения были сделаны в области нижней тон-скалы. В качестве условных раздражителей употреблялись чистые тона (от прибора Е. А. Ганике и катодного прибора Термена). Оказалось, что механическое разрушение улитки дает возможность произвести, при соответствующей оперативной технике, в достаточной степени изолированное повреждение, нашедшее свое четкое выражение в условных рефлексах. При таком интракапсулярном разрушении тонкой иглой улитки на границе средней и верхней трети выпали тоны от 315 колебаний в секунду и ниже. Была точно установлена физиологическая граница между зоной здоровой и поврежденной: в данном случае она лежала между тоном 317 колебаний и 309-315.
Переходим к вопросу о парной работе больших полушарий (опыты К. М. Быкова) (22). На одной собаке последовательно производилась выработка ДВУХ диференцировок на кололку на симметричных местах кожи. Результат получился отрицательный: диференцировку образоать не удалось. "Различение двух пунктов кожной поверхности одной Стороны не представляет затруднений, тогда как различение симметричных пунктов разных сторон, если не совсем невозможно, то во всяком случае весьма затруднительно" (стр. 294) (22). Таково заключение автора, сославшегося при этом на факты Л. С. Григорович и Н. А. Подкопаева, которым тоже удалось образовать такую диференцировку (хотя она и была нестойкой).
В связи с этим ставились эксперименты на собаках с Перерезанным corpus callosum (К. М. Быков и А. Д. Сперанский) (23). Ими была разработана оперативная методика перерезки, и у такой собаки был выработан условный электро-оборонительный рефлекс на кололку на одной стороне, а потом она пробовалась на другой. Их заключение сводилось к тому, что у собаки с перерезанным corpus cajlosum процесс возбуждения, возникающий в одном полушарии, не переходит на другое, тогда как распространение возбуждения по одному полушарию совершается так же, как и у кооперированных животных.
Представления павловской школы о физиологической природе инстинкта были изложены в докладе Г. П. Зеленого (24). По своему нервному механизму инстинкт это безусловный рефлекс. В инстинктах имеет место автоматическое возбуждение наряду с рефлекторным; например, в пищевом инстинкте - "голодная кровь", в половом - половой гормон.
В трех работах Г. П. Зеленого, А. Г. Иванова-Смоленского и А. К. Ленца был представлен переход исследований на человека.
В своей статье Зеленый (25) привел опыт физиологического анализа неврозов*. Основой этого анализа было соотношение возбуждения и торможения. Симптом "потери функции реального" при психастении П. Жанэ получил толкование, как явление парабиоза и парадоксальной фазы. Автор касался также ряда нервных и психических заболеваний и возможной их классификации на физиологической основе.
* (Материал двух докладов на Всероссийском съезде психиатров и невропатологов, в январе 1924 г.)
Работа А. Г. Иванова-Смоленского (26) была посвящена методу условных рефлексов в применении к людям и значению его для психиатрии. Он исходил из положения о применимости этого метода в эксперименте на человеке, при строго объективном наблюдении за его поведением. В статье было дано описание предложенной автором "речевой хватательной методики", по которой работа на детях и взрослых людях (больных и здоровых) ведется сотрудниками Иванова-Смоленского до сих пор.
Психиатр А. К. Ленц (27) писал о поведении душевно-больных с точки зрения физиологии высшей нервной деятельности. Он считал, что "физиология высшего отдела нервной системы" есть "фундамент психиатрии". Исследование касалось условных рефлексов у прогрессивных паралитиков различных стадий заболевания. У них было отменно ослабление тормозов. Бред преследования трактовался, как проявление иррадиированного оборонительного рефлекса. Терапия психозов с физиологической точки зрения сводится или к стремлению понизить процессы возбуждения в мозгу и усилить процессы торможения (успокаивающая терапия), или к стремлению повысить процессы коркового возбуждения и понизить всякие тормозящие влияния (стимулирующая терапия).
Особый интерес имеют три публичных лекции И. П. Павлова "Об уме вообще и русском в частности", прочитанные им в 1924 г. Эти лекции не были опубликованы.
Литература
1. Д. С. Фурсиков. О соотношении процессов возбуждения и торможения. Труды физиологических лабораторий акад. И. П. Павлова, 1924, т. I, вып. 1, стр. 3.
2. И. П. Разенков. Изменение раздражительного процесса коры полушарий головного мозга собаки при трудных условиях. (Там же, стр. 103) 1926. Реферат - Русский физиологический журнал, т. IX, вып. 5-6, стр. 571. Доклад в Обществе физиологов 3 октября 1924 г.
3. А. Д. Сперанский. Изменение взаимоотношений процессов возбуждения и торможения у собаки после наводнения. Доклад в Обществе физиологов 11 декабря 1924 г. Реферат - Русский физиологический журнал, 1925, т. VIII, вып. 3-4, стр. 139.
А. Д. Сперанский. Влияние сильных разрушительных раздражителей на собаку тормозного типа нервной системы. Труды физиологических лабораторий акад. И. П. Павлова, 1927, т. II, вып. 1.
4. М. К. Петрова. Различные виды внутреннего торможения при особенно трудном условии. Труды физиологических лабораторий акад. И. П. Павлова, 1924, т. I, вып. 1, стр. 61.
5. М. К. Петрова. Борьба со сном. Труд уравновешивания раздражительного и тормозного процессов. Сборник, посвященный 75-летию акад. И. П. Павлова, 1924, стр. 277.
6. Е. М. Крепс. Опыт индивидуальной характеристики экспериментального животного. Труды физиологических лабораторий акад. И. П. Павлова, 1924, т. I, вып. 1, стр. 119.
7. Н. А. Подкопаев. О моменте начала иррадиации тормозного процесса. Сборник, посвященный 75-летию И. П. Павлова, 1924, стр. 297. Реферат - Русский физиологический журнал, 1924, т. VII.
8. А. Г. Иванов-Смоленский. О звуковой проекции в коре больших полушарий. Сборник, посвященный 75-летию И. П. Павлова, 1924, стр. 387. А. Г. Иванов-Смоленский. Об иррадиации угасательного торможения в слуховом анализаторе собаки. Реферат - Русский физиологический журнал, 1924,. т. VII, стр. 310.
А. Г. Иванов-Смоленский. То же. Труды физиологических лабораторий, акад. И. П. Павлова, 1926, т. I, вып. 2-3, стр. 229.
9. Е. М. Крепс. Положительная индукция и иррадиация торможения в коре больших полушарий. Доклад в Обществе физиологов, 19 декабря 1922 г. Сборник, посвященный 75-летию И. П. Павлова, 1924, стр. 323.
10. Ю. П. Фролов. Пассивно-оборонительный рефлекс и его последствия. Там же, стр. 331.
Ю. П. Фролов. О рефлексе покорности и его последствиях. Реферат* - Русский физиологический журнал, 1924, т. VII, стр. 292.
* (Краткое изложение доклада, содержание которого вошло в вышеназванную работу (10).)
11. И. Р. Пророков. О влиянии положительного раздражителя на развитие отрицательной индукции. Труды физиологических лабораторий акад. И. П. Павлова, 1940, т. IX, стр. 78.
12. И. Н. Журавлев. Материалы к вопросу об отрицательной индукции и влиянии частичной экстирпации кожного анализатора у собаки. Там же, 1940, стр. 89.
13. В. В. Сирятекий. О мозаичных свойствах коры больших полушарий. Врачебное дело, 1925, вып. 1-2, стр. 7.
В. В. Сирятский. Положительная и отрицательная индукция в связи с выработкой функциональной мозаики. Реферат - Русский физиологический журнал 1926, т. IX, вып. 1.
14. В. В. Сирятский. О патологических отклонениях в деятельности центральной нервной системы при трудном балансировании процессов возбуждения и торможения. Доклад в Обществе физиологов 11 декабря 1924 г. Реферат - Русский физиологический журнал, 1925, т. VIII, вып. 3-4, стр. 136.
15. В. М. Архангельский. Относительная сила разных видов внутреннего торможения. Труды физиологических лабораторий акад. И. П. Павлова, 1924, т. I, вып. 1, стр. 71.
16. Л. А. Андреев. Материалы к изучению функциональных старческих изменений центральной нервной системы. Там же, стр. 93.
17. А. В. Вальков. Опыт изучения высшей нервной деятельности у тиреоидектомированных щенков. Сборник, посвященный 75-летию И. П. Павлова, 1924, стр. 363. Реферат - Русский физиологический журнал, 1924, т. VII, стр. 324.
18. И. П. Разенков. К вопросу о соотношении процессов возбуждения и торможения у собаки с односторонней экстирпацией gyri coronarius et ectosylvius. Архив биологических наук, 1924, т. XXIV, вып. 1-3.
19. В. В. Строганов. Образование условного рефлекса и диференцировки на синтетический раздражитель. Сборник, посвященный 75-летию И. П. Павлова, 1924, стр. 369.
20. В. А. Крылов. О возможности образования условного рефлекса на раздражитель через кровь (автоматический раздражитель). Там же, стр. 397.
21. Л. А. Андреев. Резонаторная теория слуха Гельмгольца в свете новых данных, определяющих деятельность периферического конца звукового анализатора собаки. Там же, стр. 339. Реферат - Русский физиологический журнал, 1925, т. VIII, вып. 3-4.
22. К. М. Быков. Опыты по вопросу о парной работе полушарий. Сборник, посвященный 75-летию И. П. Павлова, 1924, стр. 291. Реферат - Русский физиологический журнал, 1924, т. VII, стр. 294.
23. К. М. Быков и А. Д. Сперанский. Собака с перерезанным corpus callosum. Труды физиологических лабораторий акад. И. П. Павлова, 1924, т. I, вып. 1, стр. 47. Реферат - Русский физиологический журнал, 1924, т. VII, стр. 351.
24. Г. П. Зеленый. К теории так называемых инстинктов и ассоциативных мозговых процессов. Русский физиологический журнал, 1924, т. VII, стр. 296.
25. Г. П. Зеленый. Опыт физиологического анализа неврозов. Сборник, посвященный 75-летию И. П. Павлова, стр. 239.
26. А. Г. Иванов-Смоленский. О методе условных рефлексов в применении к людям и о значении его для психиатрии. Журнал психологии, неврологии и психиатрии, 1924, т. V, стр. 7.
27. А. К. Ленц. Поведение душевно-больных с точки зрения физиологии высшей нервной деятельности. Сборник, посвященный 75-летию И. П. Павлова, стр. 307.
При использовании материалов сайта активная ссылка обязательна:
http://anfiz.ru/ 'Анатомия и физиология человека'