Глава IX - 1911
Уже в конце 1910 г. произошел поворот в понимании Иваном Петровичем условного торможения. В своем докладе (как уже мы писали) он говорил об условном торможении как о внутреннем торможении. Поводом для изменения представления послужили опыты д-ра Н. И. Лепорского, изложенные в его диссертации "Материалы к физиологии условного торможения" (1). Сравнивая действие условного тормоза на другие виды внутреннего торможения (угасание и запаздывание) с действием гаснущих и простых тормозов, Лепорский пришел к заключению, что "в основе действия условного тормоза лежит процесс внутреннего торможения, вполне сходный с тем же процессом при угасании и запаздывании условных рефлексов" и, что условный тормоз, таким образом, есть "особый вид внутреннего торможения".
На основании чего же теперь стали считать правильным выделение условного тормоза из группы внешнего торможения? Аргументами для этого были многочисленные факты сходства процесса обобщения тормозящего действия условного тормоза и угасания и факты отсутствия растормаживания при действии условного тормоза на угасший условный рефлекс и на тормозную фазу запаздывающего рефлекса. Согласно неверной концепции растормаживания, как "торможения торможения", явление растормаживания тогда считалось характерным для действия внешних тормозов. Упомянутые же ранее данные Никифоровского о растормаживании запаздывающего рефлекса при помощи условного тормоза рассматривали, как исключение, как временное, непостоянное явление (стр. 90) (1).
Таким образом действие условных тормозов на внутреннее торможение считалось отличным ют действия внешних тормозов. Возникла новая классификация видов коркового торможения.
1) гаснущий тормоз,
I. Внешнее торможение: 2) простой тормоз,
3) общий или снотворный тормоз.
1) угасание,
2) запаздывание,
II. Внутреннее торможение: 3) внутреннее торможение, лежащее в основе
диференциации,
4) внутреннее торможение, лежащее в основе
следовых условных рефлексов,
5) условный тормоз.
Лепорский в полном согласии с предыдущими исследователями показал, что условный тормоз, выработанный для одного условного рефлекса, тормозит и другие однородные условные рефлексы (явление генерализации условного тормоза). Суммарный условный рефлекс тормозится условным тормозом, выработанным к одному из слагаемых раздражителей. Далее, чем сильнее условный раздражитель при одном и том же условном тормозе, тем меньше тормозящий эффект. Эти силовые взаимоотношения были подтверждены в последующих работах (Фурсиков и др.). Лепорский производил также опыты с диференцированием условных тормозов путем подкрепления некоторых из них. Опыты с суммацией условных раздражителей установили, что суммарный условный рефлекс больше, чем отдельные. Как уже известно, явление суммации стало изучаться в первые годы исследований высшей нервной деятельности и представляет собою факт обще-физиологического значения.
Если в диссертации П. Н. Николаева (см. главу VIII) речь шла об условном торможении как о внешнем торможении, то в 1911 г. в его статье "К анализу сложных условных рефлексов" (2) оно трактуется как торможение внутреннее. Содержание этой работы было изложено в докладе Павлова в 1910 г. Сделаем здесь только несколько дополнительных замечаний. Важно отметить, что в работе проводилась мысль о качественной равноценности возбуждения и торможения и активном характере торможения. У одной собаки свет был положительным условным раздражителем, свет плюс тон - условным тормозом и свет плюс тон плюс метроном - положительным. У другой - вертушка была положительным условным раздражителем, вертушка плюс тон - условным тормозом и вертушка плюс тон плюс чесание - положительным раздражителем. Результат опытов на обеих собаках был одинаковый: "При диференциации условно-тормозной комбинации с каким-либо раздражителем, в котором эта тормозная комбинация повторяется как общая часть, количества испытаний обоих раздражителей не могут быть произвольны, а должны находиться в таком взаимоотношении, чтобы число повторений диференцируемого раздражителя было бы, по крайней мере, в два раза меньше числа повторений условного тормоза, отношение между ними не должно быть больше 1/2". Мы уже говорили о том, что этот количественный "закон" в дальнейшем не получил подтверждения. Однако необходимо подчеркнуть в связи с опытами Николаева несомненно важное положение: возбуждение и торможение в каждом конкретном случае своего взаимодействия находятся в определенном количественном соотношении; торможение менее устойчиво, чем возбуждение, и поэтому нуждается в большем Укреплении, иначе перевес берет возбуждение. Этим объясняется давно установившаяся практика - применять тормоза и положительные условные раздражители в более или менее определенном соотношении их в опыте.
Кроме упомянутых двух работ, к проблеме коркового торможения относятся еще диссертации Белякова, Потехина и Добровольского.
Работа д-ра В. В. Белякова (3) была посвящена систематическому исследованию диференцировки. Изучалось растормаживание диференцировок и их последовательное торможение. Наличие последнего рассматривалось как доказательство того, что в основе диференцировки лежит процесс внутреннего торможения. Была получена диференцировка на 1/8 тона. Факторами, влияющими на выработку диференцировки, оказались: промежуток времени между раздражителями и степень пищевой возбудимости. Чем выше была степень диференцировки, тем последовательное торможение было сильнее. Диференцировочный тон, примененный на первом месте в опыте, тормозил сильнее, чем в сере дине опыта. Если диференцировка повторялась в течение опыта, то последовательное торможение после нее делалось больше. Был отмечу интересный и теперь факт: свежая диференцировка вызывала более сильное последовательное задерживание, чем окрепшая (явление иррадиации и концентрации диференцировочного торможения). Далее было установлено, что из посторонних раздражителей сильнее растормаживали диференцировку однородные. Последовательное торможение после диференцировки сказывалось на условных рефлексах из однородного анализатора сильнее, чем из разнородного. Более слабое диференцировочное торможение задерживало условные рефлексы однородного анализатора и не тормозило рефлексов другого анализатора. С точки зрения пространственного анализа явлений высшей нервной деятельности важен следующий вывод: "Задерживающий от диференцировки процесс, распространяясь по коре головного мозга на другие анализаторы, скорее покидает эти последние и дольше остается в однородном анализаторе; отсюда следует, что центр задерживания от диференцировки находится именно в однородном анализаторе (3). Таков ответ на вопрос: где локализовано диференцировочное торможение - в мозговом конце анализатора или в пищевом центре?
Допускалась мысль о возможности иррадиации торможения: "предполагалось, что диференцировочное задерживание может выходить за пределы мозговой части ушного анализатора и распространяться также и на другие отделы головного мозга" (стр. 147). В этой работе мы находим зародыш идеи об иррадиации торможения и возбуждения. "Путь, по которому должно пройти возбуждение, прежде чем достигнуть пункта торможения, вызванного диференцировкой, будет различен для раздражителя однородного и разнородного, для первого, очевидно, он будет короче, чем для второго" (стр. 142).
Задачей работы д-ра С. И. Потехина (4 и 5) было изучение развития запаздывания и взаимодействия угасания и запаздывания. В свое время Завадский исследовал искусственные запаздывающие условные рефлексы; Потехин работал с натуральными запаздывающими пищевыми условными рефлексами и подтвердил основные данные Завадского. При выработке таких рефлексов как тот, так и другой наблюдали фазу исчезания условных рефлексов (фазу нулей). Угасание рассматривалось как более элементарный процесс, нежели запаздывание. Запаздывание натуральных условных рефлексов оказалось труднее, чем искусственных. По данным Потехина, угасание и запаздывание, являясь видами одного и того же основного процесса - внутреннего торможения, в то же время представляют два самостоятельных процесса различной сложности и до известной степени антагонистичны один другому. Потехиным было доказано, что можно выработать диференцировку в отношении однородных условных рефлексов, совпадающих и запаздывающих (на 3 минуты). В этих опытах угасание одного условного рефлекса вело не к полному угасанию другого, а лишь к понижению его величины. В связи с этим могла быть выработана диференцировка в отношении угасания однородных условных рефлексов.
Исключительное значение имеет глава III диссертации Потехина об угасании запаздывающих условных рефлексов. Было установлено два типа угасания таких рефлексов у разных собак: 1) с обратным изменением характера кривой условной секреции и 2) без изменения характера кривой, но со снижением величины секреции.
Первый тип угасания наблюдался у собак, у которых запаздывающий рефлекс был недостаточно прочно выработан, второй - у собак, у которых он был выработан прочно. Описанное явление не получило удовлетворительного объяснения, и "вопрос о характере угасания запаздывающих условных рефлексов остался открытым" (стр. 92) (5).
Я полагаю, что этот вопрос должен стать темой дальнейшего исследования, ибо здесь лежит один из экспериментальных подходов к разрешению основного вопроса о взаимодействии возбуждения и торможения в высшей нервной деятельности. Следует не только повторить опыты Потехина на новом уровне наших знаний, но произвести ряд вариаций их, применяя кофеин и бром?
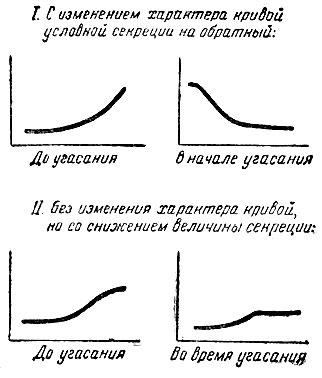
Рис. 2. Кривые угасания запаздывающих рефлексов
Эксперименты д-ра В. М. Добровольского (6) представляли дальнейшую разработку вопроса об условных следовых рефлексах. До этого Пименов и Гроссман имели дело со следовыми рефлексами, образованными на следах чесания и подкреплявшимися раствором соляной кислоты. Задачей Добровольского было выяснить возможность получения условных следовых рефлексов с другого анализатора и притом на почве пищевого подкрепления. Условными раздражителями были свет и вертушка. Образование следовых рефлексов производилось по способу Гроссмана (см. главу VII). Добровольский пришел к выводу, что этот метод образования следовых рефлексов из наличных предпочтительнее непосредственного образования (как это было у Пименова). В его опытах, как и в опытах предыдущих исследователей, наблюдались три фазы образования условного следового рефлекса: фаза угасания, фаза нулей и фаза выработки следового рефлекса, вторая фаза сопровождалась развитием сонливости. В первые две фазы имело место положительное действие экстрараздражителей. На возбудимых собак экстрараздражители действовали все время. Действие эстрараздражителей при пищевых следовых рефлексах было слабее, ем при следовых кислотных. Угасание происходило с одного раза, как в прежних опытах. Отмечено было медленное восстановление угасшего следового рефлекса. Добровольский полагал, что при осуществлении следового рефлекса происходит развитие внутреннего торможения коре. В его опытах пищевые следовые рефлексы тормозились гаснущими тормозами. Следовые пищевые рефлексы отставлялись легче, чем следовые кислотные. Добровольский производил опыты с укорочением действия условных раздражителей* и обнаружил, что это укорочение оказывало заметного влияния на величину следового пищевого рефлекса и на следовой промежуток времени (что впоследствии было подтверждено в работе Б. И. Стожарова в 1939 г.).
* (См. предыдущие опыты Л. А. Орбели (1908 г.) и последующие работы проф. Купалова и его сотрудников (Лукова, Стожарова и др.).)
Эффект генерализации экстрараздражителей объяснялся тогда, как "торможение торможения" (стр. 62) (6), чему противоречили факты, полученные самим автором (стр. 63, 64, 83), в них было проявление простой иррадиации возбуждения. Идея не только помогает видеть факты, но также способствует иногда (как в данном случае) тому, чтобы их не видеть.
Может ли играть положительную роль в деле специализации условного рефлекса само по себе многократное повторение основного раздражителя? Этот вопрос уже давно стоял в павловской лаборатории, и утвердительный ответ на него был дан в работе д-ра Ю. В. Снегирева (7). Он делал опыты с многократным на протяжении более или менее длительного времени повторением одного основного тона, подкреплявшегося едой. В начале таких опытов "необычные" тона действовали положительно, а потом нет. Таким образом удалось получить отличение других тонов, отличавшихся от основного на 21/2 тона. В данном случае специализация условного звукового рефлекса возникла (как теперь нам ясно) в результате развития процесса концентрации возбуждения*.
* (См. диссертацию П. С. Купалова, 1915 г.)
Подобно некоторым другим, автор неудачно пытался сочетать теорию условных рефлексов с идеалистической философией Маха и Авенариуса: "Проф. Павлов встал на путь исключения интроекции... В этом случае, устраняя интроекцию, мы вместе с тем устраняем и целый ряд ложных проблем и, с этой точки зрения, учение об условных рефлексах является практическим применением эмпириокритических начал" (стр. 44) (7). На это мы должны сказать, что исключение Павловым субъективно-психологического рассмотрения явлений высшей нервной деятельности ничего общего не имеет с субъективно-идеалистической системой Авенариуса и Маха. Правильность этого утверждения можно иллюстрировать и на работе самого Снегирева, не приведшего никаких доказательств своей точки зрения.
Исследования д-ра Я. Е. Егорова (8) о влиянии пищевых условных рефлексов друг на друга являлись продолжением работ Бабкина и Былины над простым торможением. Егоров выработал ряд условных рефлексов на почве разных по качеству пищевых подкреплений и изучал влияние одних условных рефлексов на другие, а также влияние того или иного безусловного пищевого раздражения на разнородный условный рефлекс. Таким образом он обнаружил, что предварительно примененный "искусственный" "сырный" условный рефлекс (например, на вертушку) уменьшал условный рефлекс, связанный с подкреплением мясо-сухарным порошком. Это уменьшение было заметно как при подкреплении первого рефлекса едой сыра, так и без подкрепления. В случае неподкрепления условного рефлекса сыром уменьшение условного пищевого рефлекса, по мнению автора, зависело не от угасания, а от специфических свойств самого сыра. С этим выводом полностью едва ли можно согласиться, так как здесь, кроме отрицательной индукции, все-таки должно было сказываться и влияние угасания "сырного" рефлекса вследствие его неподкрепления. Автор правильно рассматривал указанное торможение условного пищевого рефлекса, как "простое торможение", т. е. как вид внешнего торможения. Далее, предварительная еда сахара уменьшала условный "сырный" рефлекс, а еда сыра уменьшала натуральный мясо-сухарный условный рефлекс. Одноминутная еда сахарного песка также уменьшала натуральный мясо-сухарный условный рефлекс.
Кроме этих фактов внешнего торможения, в данной работе мы находим и указание на возможность "иррадиации раздражения из одной части пищевого центра в другую" (стр. 31) (8), и (как на следствие этого) на эффект суммации (стр. 85).
Значение исследований Егорова заключается: 1) в разработке павловского учения о пищевом центре, 2) в развитии учения о простом (внешнем) торможении. Работа Егорова дала фактический материал к вопросу об анатомической локализации корковых функций в связи с развитием представления о мозаичной структуре коркового пищевого центра. "Значит, та группа клеток пищевого центра, которая заведует поступлением в организм сахара, так же самостоятельна и различна, как и группа клеток, заведующих поступлением сыра" (стр. 77) (8). Если стать на эту точку зрения, то надо предположить, что в корковом пищевом центре существует столько же отдельных специфических групп клеток, сколько в природе различных вкусовых веществ. И, значит, должны быть специфические клетки для таких веществ, которые мы, может быть, никогда и не попробуем. По мнению автора диссертации, существуют корковые клетки специально для сыра, сахара, колбасы, масла, хлеба, мяса и т. д., и т. д. Такое упрощенное представление должно нас привести только в тупик грубо-механического и атомистического понимания связи корковых функций с материальной структурой. Правильнее, конечно, все указанные факты объяснить с точки зрения позже развитого Павловым принципа "функционального динамического структурного комплекса"*. В связи с другими исследованиями Иван Петрович писал: "Едва ли можно думать о том, что каждому нами употребляемому звуку отвечает особая группа нервных клеток, воспринимающих элементарные звуковые раздражители, из которых слагается наш звук. Вероятнее, что в случае каждого нашего звукового раздражителя дело идет о динамическом структурном комплексе, элементы которого, соответствующие клетки, входят и в другие динамические комплексы при применении других сложных звуков"**. По поводу того, как представлять себе корковые "пункты", Иван Петрович говорил: "Представлять их анатомически трудно, надо представлять их динамически. Одни и те же клетки могут выполнять разные функции. Нарушается не деятельность отдельных клеток, а динамический комплекс"***.
* (См. И. П. Павлов. Экспериментальная патология высшей нервной деятельности 1935. Отдельная брошюра, стр. 23, Изд. Академии наук СССР.)
** (См. И. П. Павлов. Экспериментальная патология высшей нервной деятельности 1935. Отдельная брошюра, стр. 23, Изд. Академии наук СССР.)
*** (Стенограмма Павловской "среды" 23 мая 1934 г.)
Именно с этой точки зрения надо понимать факты Егорова. Диссертация д-ра И. С. Цитовича (9) была посвящена происхождению и образованию натуральных условных Рефлексов. Решался вопрос: есть ли натуральные условные рефлексы слюнных желез результат наследственной передачи или жизненно опыта животного? Цитович пришел к выводу, что натуральные условные рефлексы есть результат индивидуального жизненного опыта животного. Мы вправе это заключение распространить на все условные рефлексы.
Для опытов было взято пять щенят одного помета. После того как их отняли от матери, они воспитывались на одном коровьем молоке. Для сравнения с натуральными рефлексами у них были образованы и "искусственные" условные рефлексы на звуки, запахи и световые раздражения. Таким образом было установлено отсутствие у них натуральных условных рефлексов на вид и запах хлеба и мяса. После же кормления их хлебом, сухарным порошком и мясом эти натуральные рефлексы выработались. Для сравнения исследовались контрольные взрослые собаки и щенята.
Число сочетаний для образования условных натуральных рефлексов было очень незначительно. Цитович полагал, что двигательная реакция на запахи незнакомых съедобных веществ может иметь инстинктивную основу, если только это не приобретено в течение первых недель жизни. Ограничивая знакомство животного с безусловными раздражителями, можно было получить экземпляры с очень упрощенными сложно-нервными процессами. Образование условных натуральных рефлексов происходило постепенно, прежде всего образовывалась связь безусловного раздражителя с обонятельным сигналом, позднее в эту связь вступали слуховые и зрительные условные раздражения. Цитовичу удалось получить "искусственные" условные рефлексы и на желудочные железы. Он показал, что главные свойства условных рефлексов остаются в полной силе и для желудочных желез, что происхождение и образование натуральных условных рефлексов на желудочные железы таково же, как и в отношении слюнных. Рефлекс с полости рта на желудочные железы рассматривался, как рефлекс безусловный.
С. И. Потехин (10) (после Завадского и Никифоровского) продолжил разработку фармакологии условных рефлексов. Он делал опыты с этиловым алкоголем, морфием и апоморфином. Доза:, алкоголя была равна 3,0-4,0 абс. алкоголя, вводимого клизмой в 3-10% разведении при 37° С. Как и Завадский, Потехин наблюдал растормаживание угасания условного пищевого натурального рефлекса под влиянием алкоголя. При применении этилового алкоголя (согласно выводам Потехина) необходимо иметь в виду побочное его действие на слюнную железу, а именно: "слюнотечение, наблюдаемое непосредственно за введением в организм собаки этилового алкоголя, есть натуральный условный рефлекс на запах алкоголя, зависящий от раздражения периферических окончаний обонятельных нервов полости носа". Автор считал, что "этиловый алкоголь является условным отвергаемым раздражителем полости носа собаки". По поводу указанного побочного действия алкоголя можно возразить, что здесь мы имеем дело не с условным, а с безусловным рефлексом с рецепторов полости носа, и поэтому правильнее говорить не об угасании этого побочного эффекта, а об адаптации к запаху. На основании своих опытов с морфием и апоморфином Потехин полагал, что необходимо также считаться с побочным действием этих веществ на рвотный центр. Падение условных рефлексов зависело не от наркотического действия морфия, а от возбуждения рвотного центра. Автор писал, что "морфий, применяемый в малых дозах (1/4-11/2 миллиграмма), действует на ход условного слюноотделения в качестве простого тормоза".
К этому же (1911) году относится работа д-ра М. Н. Ерофеевой (11) с раздражением кожи фарадическим токам, как условным возбудителем слюнных желез. На основании этих экспериментов ею была написана диссертация, рассмотрением которой мы займемся в следующей главе.
Д-р Н. И. Красногорский (12) изучал иррадиацию и концентрацию торможения и вопрос о локализации кожного и двигательного анализаторов в коре. Он подтвердил, что кортикальные торможения подчиняются законам иррадиации и концентрирования. Было доказано, что торможение, возникшее в любом пункте анализатора, весьма быстро иррадиирует, захватывает всю кору больших полушарий. Красногорский считал, что "появление активных диференциаций обусловливается образованием в коре локализованных условных очагов (центров) раздражения", то же самое относится и к "инактивной диференциации" очагов торможения. Иррадиированное торможение медленно исчезает, но может быть с различной скоростью сконцентрировано. Изучая иррадиацию и концентрацию торможения от диференцировок на покалывание кожи, Красногорский натолкнулся на непонятный факт, а именно: в зрительном анализаторе последовательное торможение от "инактивного" (тормозного) покалывания было, а при этих же условиях в том же (кожном) анализаторе его не было (см. таблицу XI на стр. 57-59 и вывод 5) (12). Тогда это было объяснено тем, что рассеянное последовательное торможение тем меньше влияет на условные очаги раздражения, чем выше организованы эти очаги раздражения. Следовательно, в приведенном случае очаг условного возбуждения в кожном анализаторе оказался более устойчивым, чем в зрительном. Во всяком случае, этот факт говорил против простого линейного представления о процессах иррадиации и концентрации.
Далее наблюдалось, что при повторении диференцировок возникает сон. Функция какой-либо "диференциаций", возникшей на почве последовательного торможения от другой, извращается, т. е. имели место факты растормаживания "при взаимодействии двух разнородных диференциаций": 1) при одновременном действии двух диференцировочных раздражителей и 2) при применении одной диференцировки на фоне последовательного торможения от другой (см. стр. 71-73 и вывод 8) (12). Таким образом, уже в то время Красногорский Доказал возможность растормаживания при взаимодействии двух внутренних торможений; поэтому неправильно было считать явление растормаживания специфичным только для действия внешних тормозов, на чем основывалась, как было нами упомянуто, диссертация Лепорского.
На основании исследований Красногорского было установлено правило распространения локальных "диференциаций" на симметрические пункты - вопрос, послуживший предметом для экспериментирования и дальнейших дискуссий.
Вторая часть разбираемой работы относится к проблеме корковой локализации. В результате опытов с экстирпацией gyri sygmoidei, coronarii et et ectosylvii Красногорский пришел к следующим заключениям. Моторная зона коры есть тоже анализатор - двигательный анализатор. В области gyrus sygmoideus локализирован корковый двигательный анализатор (и часть кожного). Cyri coronariua et ectosylvius суть части кожного анализатора. Если производилось раздражение кожной поверхности с выпавшей условно-рефлекторной функцией, то оно вызывало торможение. Было описано несколько фаз восстановления условных рефлексов после удаления соответствующих корковых областей. Красногорский считал, что элементы в пределах одного коркового анализатора функционально равны. Работа Красногорского положила начало исследованию проприоцептивных условных рефлексов.
Дальнейшим исследованием нормального и поврежденкого звукового анализатора собаки занимался д-р Б. П. Бабкин (13 и 14). Он делал опыты с грубыми и тонкими диференцировками (на порядок тонов, музыкальные фразы с разным порядком пауз) до операции и после удаления коркового слухового анализатора с обеих сторон. Вначале наблюдался общий факт - уточнение работы анализатора по мере практики. После мозговых операций было констатировано постоянное явление ослабления тормозного процесса. Грубые диференцировки после экстирпации сохранились, а тонкие оказались нарушенными. "Различение порядка следования звуков, места паузы в музыкальной фразе, а также реакция на кличку для поврежденного звукового анализатора были недоступны" (стр. 286) (13). Таким образом, по общему заключению автора то состояние, которое Мунк называл "душевной глухотой", в свете объективного исследования сложно-нервных явлений представлялось лишь понижением тонкости работы поврежденного звукового анализатора.
Изучение условных рефлексов у собак без передних половин обоих полушарий было предметом работы д-ра Н. М. Сатурнова (15). Фактически это составляло продолжение исследований Демидова (см. главу VII), но Сатурнов оставлял на месте обонятельные доли. Результаты Демидова были подтверждены и кроме того, добавлен новый факт - возможность получения у таких собак условного рефлекса на запах. В итоге опытов на 5 животных Сатурнов пришел к следующим выводам: 1) передние половины больших полушарий не заключают какого-либо общего самостоятельного механизма, управляющего всей сложно-нервной деятельностью животного, а наблюдаемое нарушение в деятельности скелетной мускулатуры после удаления передних половин есть результат частичного выпадения функций известных отделов коры; 2) удаление передних половин больших полушарий ведет к нарушению образования условных рефлексов на звук (вероятно, в зависимости от прохождения в передних половинах проводников ушного анализатора); 3) резкое нарушение кожного анализатора, наряду с незначительным поражением двигательного аппарата, дает право на выделение "особого самостоятельного анализатора двигательного (внутреннего)" (что совпадает с упомянутым заключением диссертации Красногорского).
Д-р Г. П. Зеленый (16) повторил опыты Гольца. У одной собаки в два приема были удалены большие полушария. Попытки образовать у такой собаки условные рефлексы (с полости рта и с уха) оказались безрезультатными. У этого животного наблюдалось слюноотделение от вливания воды в полость рта, на основании чего д-р Зеленый считал, что так называемый "условный водяной рефлекс" (описанный Демидовым) есть не условный, а безусловный. Может ли вода сама по себе быть возбудителем слюноотделения? Этот вопрос теперь в результате исследований Д. С. Фурсикова и др. надо считать разрешенным положительно.
Спор Гольца и Мунка (1894 г.) по поводу поведения бесполушарных собак разрешался на основании данных Зеленого в пользу Гольца, т. е. рефлексы у таких собак считались зависящими от нормальных специфических раздражений органов чувств (Гольц), а не являющимися результатом чрезмерно сильных раздражений, вызывающих действие даже посторонних нервных путей (Мунк). Исследованная Зеленым (16а) собака прожила 11 месяцев и 4 дня. Морфологическое исследование показало, что оба больших полушария действительно были удалены полностью. Thalami optici и cotpores striati оказались уменьшенными и до 1/3 своей нормальной величины. Передние бугры четверохолмия подверглись перерождению*.
* (В течение последних лет опытами на бесполушарных животных (собаках, обезьянах, кошках и кроликах) занимался проф. Н. Ф. Попов (Москва).)
В работах павловской школы с удалением частей коры имело место противоречие двух групп фактов: с одной стороны (Эльяссон, Бабкин и др.), было установлено несомненное ослабление тормозных функций коры после удаления той или иной ее части, с другой стороны (по данным того же Бабкина и Красногорского), получалось, что при удалении лобных долей условные рефлексы с кожи пропадают, а условные тормоза с пораженных кожных участков остаются? Что касается первого факта, то он подтвержден многочисленными исследованиями. Второй же факт не может считаться твердо установленным и нуждается в проверке. По поводу последнего надо сделать два замечания: 1) непонятно, как можно судить о сохранении тормозов, если их положительные пары дают нули?, 2) необходимо в этом случае отличать возможность безусловного торможения от условного.
На основании опытов Красногорского и Рожанского И. П. Павлов делал в Обществе русских врачей доклад об основных правилах работы больших полушарий (17). Здесь им впервые была поставлена развернутая проблема коркового торможения и сформулированы основные законы движения нервных процессов в коре больших полушарий. Закон иррадиации и концентрации был распространен и на тормозной процесс. "Как раздражение сначала иррадиирует, а затем концентрируется, так и задерживание сперва разливается, а затем сосредоточивается" (стр. 108) (17). Иван Петрович полагал, что концентрирование нервных процессов направлено "в известное русло, в определенные линии или точки мозга" (стр. 101). Он говорил, что "эти факты вместе с тем представляют существеннейший довод за то, что раздражение и задерживание это лишь разные проявления одного и того же процесса" (стр. 108)*.
* (Курсив мой - Ф. М.)
Сопоставление физиологического и психологического мышления было почти постоянной темой всех выступлений Ивана Петровича. И в этом докладе он говорил: "Почему же мы до этого обращались трусливо к прежнему субъективному методу? Секрет простой: потому, что это - метод беспричинного мышления, потому что психологическое рассуждение - есть адетерминическое рассуждение, т. е. я признаю явление, происходящее ни оттуда, ни отсюда. Я говорю: собака подумала, собака желает - и удовлетворяюсь. И это есть фикция. А причины для явления так и нет. Стало быть, и удовлетворение при психологическом толковании тоже только фиктивное, без основания. Наше объективное объяснение есть истинно-научное, т. е. всегда обращающееся к причине, всегда ищущее причину" (стр. 108). Разве не ясно из приведенной цитаты, что Павлов по сути дела выступал здесь против идеализма.
В своем докладе "собака с разрушенным в больших полушариях кожным анализатором" (18) Павлов (на основании опытов д-ра Сатурнова) подчеркнул, что в данном случае мы имели "пример изолированного дефекта кожного анализатора без нарушения двигательного анализатора" (стр. 112). Был сделан вывод, что к пяти внешним анализаторам надо прибавить шестой - внутренний, двигательный анализатор.
Последний доклад Ивана Петровича касался опытов Белякова, изучавшего процесс диференцирования раздражителей в больших полушариях (19). Темой доклада был корковый механизм анализа. Иван Петрович привел разбор экспериментальных доказательств того, что в основе диференцирования лежит торможение: факты последовательного торможения, растормаживания и суммирования. Он тогда думал, что диференцировочное торможение локализировано в корковом анализаторе соответствующего раздражителя. Был подчеркнут факт "чрезвычайной замещаемости мозговой массы". Это вытекало из всех опытов с экстирпациями. Здесь же мы находим зародыш павловского учения о корковой локализации функций; исходным пунктом рассуждений об этом Павлова был факт обобщенного условного рефлекса.
Возвращаясь к теме о физиологии и психологии, Иван Петрович тогда сказал: "Это - большая задача, к которой я себя готовлю - перебрать все психологические понятия и показать в сопоставлении с нашим объективным материалом, до какой степени они фантастичны и носят грубый эмпирический характер, который представляет непреодолимую помеху при анализе тонких явлений высшей нервной деятельности" (стр. 122).
Литература
1. Н. И. Лепорский. Материалы к физиологии условного торможения. Диссертация. 1911.
2. П. Н. Николаев. К анализу сложных условных рефлексов. Архив биологических наук, 1911, т. XVI, вып. 5.
3. В. В. Беляков. Материалы к физиологии диференцирования внешних раздражений. Диссертация. 1911.
4. С. И. Потехин. Взаимное отношение различных видов внутреннего торможения условных рефлексов. Доклад в Обществе русских врачей 5 мая 1911 г. Труды Об-ва русских врачей, т. 78.
5. С. И. Потехин. К физиологии внутреннего торможения условных рефлексов. Диссертация. 1911.
6. В. М. Добровольский. О пищевых следовых рефлексах. Диссертация. 1911.
7. Ю. В. Снегирев. Материалы к учению Павлова об условных рефлексах. Клинические монографии, изд. Практическая медицина, январь, 1911.
8. Я. Е. Егоров. Влияние пищевых условных рефлексов друг на друга. Диссертация. 1911.
9. И. С. Цитович. Происхождение и образование натуральных условных рефлексов. Диссертация. 1911.
10. С. И. Потехин. К фармакологии условных рефлексов. Доклад в Обществ русских врачей 5 мая 1911. Труды Об-ва русских врачей, т. 78, стр. 231.
11. М. Н. Ерофеева. Раздражение кожи фарадическим током, как условный возбудитель слюнных желез. Доклад в Обществе русских врачей 27 октября 1911. Труды Об-ва русских врачей, т. 79, стр. 62.
12. Н. И. Красногорский. О процессе задерживания и о локализации кожного и двигательного анализаторов в коре больших полушарий у собак. Диссертация. 1911.
13. Б. П. Бабкин. Дальнейшие исследования нормального и поврежденного звукового анализатора собаки. Доклад в Обществе русских врачей 5 мая 1911. Труды Об-ва русских врачей, т. 78, стр. 249.
14. Б. П. Бабкин. Так называемая "душевная глухота" перед объективным анализом сложно-нервных явлений. Русский врач, 1911, № 51, стр. 1934.
15. Н. М. Сатурнов. Дальнейшие исследования условных (слюнных) рефлексов у собаки без передних половин обоих полушарий. Диссертация. 1911.
16. Г. П. Зеленый. Собака без полушарий головного мозга. Доклад в Обществе русских врачей 27 октября 1911. Труды Об-ва русских врачей, т. 79, стр. 50.
16-а. 2-е сообщение 9 февраля 1912. (Труды Об-ва русских врачей, т. 79, стр. 147).
17. И. П. Павлов. Основные правила работы больших полушарий. (На основании опытов д-ров Н. И. Красногорского и Н. А. Рожанского. Доклад в Обществе русских врачей 24 марта 1911. Двадцатилетний опыт (цитировано по изд. 1923 г.).
18. И. П. Павлов. Собака с разрушенным в больших полушариях кожным анализатором. (На основании опытов д-ра Н. М. Сатурнова). Доклад в Обществе русских врачей 28 апреля 1911 г. Двадцатилетний опыт.
19. И. П. Павлов. Процесс диференцирования раздражителей в больших полушариях. (На основании опытов д-ра В. В. Белякова). Доклад в Обществе русских врачей 1911 г. Двадцатилетний опыт.
При использовании материалов сайта активная ссылка обязательна:
http://anfiz.ru/ 'Анатомия и физиология человека'