§ 37. Филогенетические преобразования нервной системы
Ранние стадии филогенеза. Эволюция рецепторов у животных шла параллельно с усложнением отношений, существующих между организмом и средой. Эта эволюция началась с выделения на поверхности тела животного специальных чувствительных клеток. Нервная система на этой стадии развития (кишечнополостные) сетевидная. Она представлена расположенными под эктодермальным покровом тела нервными клетками, многообразно соединенными своими отростками. Они связаны с рецепторными клетками эктодермы и мышечными клетками. Всякое раздражение, воспринятое рецепторными клетками, нервные клетки передают мышечным. Следовательно, уже на самых ранних стадиях развития нервная система закладывается как связующее звено между рецепторами и рабочими органами. Реакция организма на этой стадии эволюции возможна лишь при непосредственном соприкосновении раздражителя с рецепторной клеткой и выражается в примитивных, главным образом обобщенных, движениях всего тела.
В дальнейшем разбросанные по всей поверхности тела рецепторные клетки концентрируются в определенных местах: в области ротового отверстия, на щупальцах и т. д. (черви и др.). Одновременно происходит функциональная дифференциация рецепторных клеток: они начинают отвечать возбуждением только на определенные - адекватные - раздражения, например одни - на световые, другие - на механические и т. д. На этой стадии рецепторы возбуждаются и на расстоянии. В строении нервной системы наступает централизация: ее клетки скапливаются в узлы, ганглии, соединенные продольными и поперечными тяжами. Так возникает ганглиозная нервная система. Нервные узлы становятся рефлекторными центрами для тех частей тела, в которых они расположены. Поэтому двигательные реакции оказываются дифференцированными.
Позднее рецепторы, воспринимающие раздражение на расстоянии, концентрируются на переднем конце тела, который в первую очередь входит в соприкосновение с внешней средой. В дальнейшем здесь образуются органы зрения, обоняния и других чувств. Расположенный спереди нервный ганглий увеличивается и начинает осуществлять координацию движений всех частей тела. Так происходит процесс цефализации (образования головы).
У хордовых происходит развитие качественно новой формы нервной системы - трубчатой. Образование у них головного мозга, как и возникновение головного ганглия у беспозвоночных, вызывается скоплением на переднем конце тела важнейших органов чувств: равновесия (вестибулярного), обоняния, вкуса, зрения и слуха. У переходной формы от беспозвоночных к позвоночным - ланцетника, не имеющего развитых дистантных рецепторов и воспринимающего лишь свето-тень, головной мозг находится в зачаточном состоянии. В дальнейшем в головном мозге позвоночных прогрессивно развиваются большие полушария. Одновременно происходит специализация анализаторов и координация их деятельности с двигательными реакциями.
Эволюция спинного и головного мозга позвоночных. Спинной мозг круглоротых занимает все протяжение позвоночного канала. Белое вещество построено примитивно и состоит из безмякотных волокон. Серое вещество концентрируется вокруг центрального канала (как у ланцетника), но рогов еще не образует. В связи с развитием жаберного аппарата самым обширным отделом головного мозга является продолговатый мозг. Мозжечок развит очень слабо. В среднем мозге намечается разделение на два бугра - зрительные доли (переднее двухолмие млекопитающих). В межуточном мозге уже имеются его три отдела; хорошо развиты надбугорная и особенно подбугорная области, связанные с обонянием и интерорецепцией. Небольшой конечный мозг состоит из серого вещества, окружающего полость желудочка и покрытого эпителиальным плащом.
В спинном мозге селяхий серое вещество разделяется на передние и слабовыраженные задние рога; волокна миелинизированы. В продолговатом мозге, который связан с иннервацией ротовой полости и внутренних органов, ядра нервов велики и выдаются на поверхности. Мозжечок большой, что связано с быстротой движений в воде (рис. 136, I). В его со став входят тело и парные ушковидные доли (гомологичные клочкам полушарий мозжечка млекопитающих). В среднем мозге хорошо выражено двухолмие, функционирующее как высший регуляторный центр, к которому подходят проводники всех видов чувствительности. В межуточном мозге особенно велика подбугорная область и все еще мал зрительный бугор. Конечный мозг имеет непарный желудочек, и спереди переходит в парные обонятельные доли. На нижней поверхности плаща появляются нервные клетки, образующие древнюю кору. Она слабо отделена от базальных ганглиев и получает импульсы от обонятельных долей.
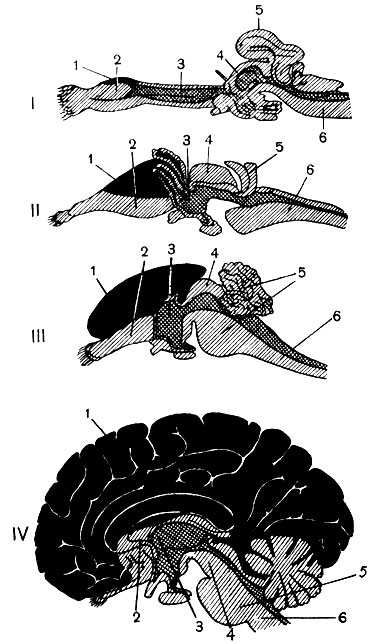
Рис. 136. Строение головного мозга: I - акулы; II - ящерицы; III - грызуна (кролика); IV - человека; 1 - плащ и кора я 2 - базальные, или подкорковые, ганглии; 3 - межуточный мозг; 4 - средний мозг; 5 - задний мозг; 6 - продолговатый мозг
С выходом позвоночных животных на сушу в их нервной системе произошли большие изменения, обеспечившие приспособления к новым условиям существования. Особое значение в преобразовании мозга сыграли два обстоятельства: 1) переход ведущей роли от органов обоняния и вкуса к органам зрения и слуха и 2) замена характерного для живущих в воде рыб передвижения при помощи всего тела более совершенным - при помощи конечностей. Перестройка мозга коснулась главным образом больших полушарий, в плаще которых около 250 млн. лет назад началось усиленное развитие коры.
В спинном мозге рептилий хорошо выражены задние и особенно передние рога серого вещества. Они делят лежащее снаружи белое вещество на канатики: передние, боковые и задние. В продолговатом мозге развиваются тонкий и клиновидный бугорки, а также верхнее сенсорное ядро тройничного нерва. В мозжечке (рис. 136, II) хорошо развиты тело и ушковидные доли. В среднем мозге у некоторых ящериц и змей позади зрительных долей выдаются еще два бугра (заднее двухолмие млекопитающих). В покрышке ножек впервые появляется красное ядро. В межуточном мозге зрительный бугор относительно велик и уже делится на ряд ядер. Одновременно развиваются и волокнистые связи их с корой.
Конечный мозг большинства рептилий состоит главным образом из базальных или подкорковых ганглиев. Кроме древней, развивается старая кора, которая хорошо отделена уже от базальных ганглиев и представляет собой переход от древней к более высокоразвитой новой коре. Начав свое развитие у амфибий на медиальной поверхности полушария, старая кора распространяется у рептилий и на дорсальную поверхность. Однако вся эта филогенетически ранняя кора продолжает оставаться связанной главным образом с обонятельными и интерорецептивными импульсами. В то же время у рептилий на боковой поверхности полушария появляется зачаток еще слабо развитой новой коры в виде так называемой боковой коры. С ее появлением конечный мозг начинает принимать импульсы от всех органов чувств, приобретает общие корреляционные функции.
Спинной мозг млекопитающих укорочен и оканчивается тонкой концевой нитью. В местах отхождения корешков спинномозговых нервов конечностейt он утолщен. В нем особенно развиты передние канатики, где прогрессивно увеличивается количество волокон передних корково-спинномозговых путей; становится немного больше и перекрещенных волокон и в корково-снинномозговых путях боковых канатиков. У собаки волокна этих пирамидных путей составляют около 7% всего белого вещества спинного мозга, у обезьян - 20% и у человека - 30%. Эти волокна, соединяя кору больших полушарий с двигательными нейронами передних рогов, все более подчиняют ее влиянию работу мышц.
Продолговатый мозг невелик (рис. 136, III), в его передних канатиках развиваются пирамиды, в боковых - большие оливы и нижние ножки мозжечка. В вентральной части моста появляется новая система клеток и волокон, увеличивающая основание моста, впервые появляются и средние ножки мозжечка. В мозжечке прогрессивно развиваются полушария, связанные с корой больших полушарий через основание моста. В среднем мозге вместо двухолмия развивается четверохолмие, причем наиболее сложный анализ зрительных раздражений перемещается из переднего двухолмия в латеральное коленчатое тело межуточного мозга и затылочную область коры. Так же и в слуховом анализаторе наиболее сложные функции передаются из ядер улиткового нерва в ромбовидной ямке и из заднего двухолмия в медиальное коленчатое тело и височную область коры. Прогрессивное развитие претерпевают зрительные бугры межуточного мозга, становясь коллектором всех видов чувствительности, направляющейся в кору больших полушарий. В конечном мозге огромного развития достигает кора, разрастающаяся не только назад (затылочные доли), но и вперед (лобные доли) и вниз (височные доли). У однопроходных, сумчатых, насекомоядных и грызунов полушария еще относительно невелики и имеют гладкую поверхность. У хищных и копытных возникают борозды, сильно увеличивающие поверхность коры. В палеогене с появлением приматов (60 млн. лет назад) полушария постепенно покрываются извилинами и бороздами, характерными для обезьян и человека (рис. 136, IV). Ведущая роль в регуляции функций переходит к новой коре, которая связана со всеми видами чувствительности и в ассоциативных областях которой (теменные, лобная) осуществляются высшие интегративные функции. Возросшая главным образом за счет увеличения ассоциативных областей, новая кора сдвигает древнюю и старую кору на нижнюю и медиальную поверхности полушария. В ряду млекопитающих новая кора занимает:
у ежа - 32% всей поверхности полушария у кролика - 56% " " " у собаки - 84% " " " у человека - 96% " " "
Длительность развития гоминид принимается в 1 млн. лет. Необычайное усложнение структуры и функций мозга за это время объясняется действием социальных факторов.
При использовании материалов сайта активная ссылка обязательна:
http://anfiz.ru/ 'Анатомия и физиология человека'