§ 35. Передний мозг
Межуточный мозг. Межуточный мозг (Атл., 77) в основном скрыт полушариями. В нем различают следующие образования: парные зрительные бугры, латеральные и медиальные коленчатые тела, подбугорную и надбугорную области. Внутри межуточного мозга находится третий желудочек.
Зрительный бугор, или таламус (thalamus opticus), - крупное эллипсоидное тело (рис. 125). Снизу он сливается с подбугорной областью, латерально сращивается с большим полушарием, а его медиальная поверхность образует боковую стенку III желудочка.
Основная пластинка мозговой трубки эмбриона в среднем мозге заканчивается, и зрительный бугор (вместе со всем передним мозгом) оказывается производным крыльной пластинки, и здесь нет двигательных ядер.
Зрительный бугор состоит из серого вещества, группирующегося в ядра (рис. 128). В нижней его части лежит вентральное ядро. В последнем оканчиваются спинно-бугорный путь, медиальная петля, тройничная петля, верхние ножки мозжечка и другие пути, проводящие импульсы от кожи и проприорецепторов. В переднем ядре оканчивается сосково-бугорный путь, приносящий многокомпонентные импульсы. На каудальном полюсе бугра, в так называемой подушке, оканчивается часть волокон зрительного пути. В настоящее время ядра таламуса, переключающие импульсы на центральные поля анализаторов в коре больших полушарий, относят к проекционным или релейным. Другие ядра, как, например, латеральное и большая часть подушки, передают возбуждение на межанализаторные или ассоциативные области коры; эти ядра относят к ассоциативным.
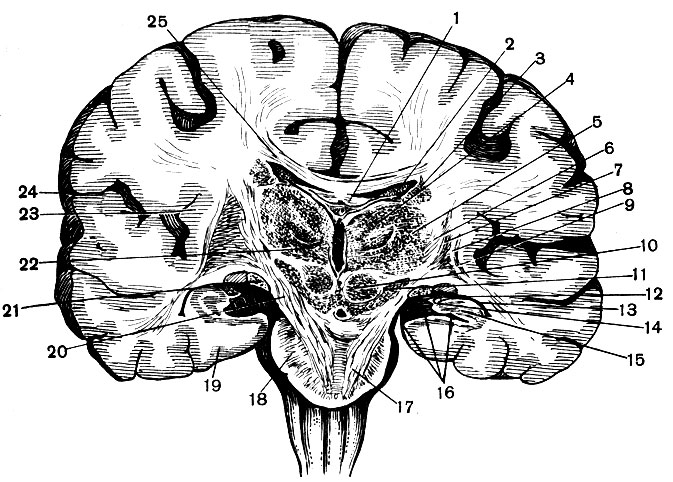
Рис. 128. Фронтальный разрез головного мозга в области ножек мозга: 1 - свод; 2 - сосудистое сплетение в средней части бокового желудочка; 3 - хвост хвостатого ядра; 4 - медиальное и 5 - латеральное ядра зрительного бугра; 6 - внутренняя капсула; 7 - чечевицеобразное ядро; 8 - наружная капсула; 9 - ограда; 10 - красное ядро; 11 - черная субстанция; 12 - сосудистое сплетение нижнего рога бокового желудочка (13); 14 - бахромка свода; 15 - гиппокамп; 16 - гиппокампова борозда; 17 - корково-спинномозговой путь; 18 - мост; 19 - парагиппокампова извилина; 20 - основание ножки мозга; 21 - зрительный путь; 22 - вентральное ядро; 23 - островок; 24 - боковая борозда; 25 - мозолистое тело
Как и в стволовой части мозга, в зрительном бугре есть образования ретикулярной формации. Они расположены между его ядрами и снаружи от латерального и вентрального ядер таламуса и получили название его неспецифических ядер. Полагают, что эти ядра оказывают на кору и подкорковые узлы более диффузное влияние, чем релейные таламические ядра; вызывают не только возбуждающий, но и отчетливый тормозной эффект. Подобно ретикулярной формации ствола, неспецифические ядра таламуса не несут каких-либо особых высших интегративных функций, но в известной мере участвуют в регуляции различных афферентных влияний, идущих по анализаторным системам.
Отростки нервных клеток зрительного бугра образуют таламическую лучистость (лучистый венец). Волокна ее направляются к большим полушариям, где они оканчиваются главным образом в коре, а также на клетках подкорковых узлов. Вследствие этого зрительный бугор входит в состав экстрапирамидной системы (Атл., 88).
Латеральное коленчатое тело (corpus genie ulatum laterale) (рис. 126) расположено вблизи подушки и латерального корешка зрительного пути, волокна которого в нем оканчиваются.
Медиальное коленчатое тело (corpus geniculatum mediale) лежит на уровне поперечной бороздки четверохолмия. В этом теле оканчиваются волокна латеральной петли, приходящие в составе ручки нижнего двухолмия. Волокна нервных клеток коленчатых тел в составе зрительной лучистости (вместе с волокнами подушки) и слуховой лучистости направляются к коре больших полушарий.
Подбугорная область, или гипоталамус (hypothalamus), - это та часть межуточного мозга, которая заметна на основании головного мозга. Заднюю часть области образуют два сосковидных тела (рис. 127). В них оканчиваются колонки свода - пути старой коры конечного мозга. Волокна сосковидных тел образуют сосково-бугорный путь, по которому импульсы достигают передних ядер зрительного бугра. В последнее время сосковидные тела, как и передние ядра зрительных бугров, связываются с лимбической системой и организацией поведенческих реакций. Спереди от сосковидных тел лежит серый бугор. Суживаясь, он переходит в воронку, проникающую в ямку турецкого седла сквозь его диафрагму. На воронке как бы подвешен гипофиз. Серый бугор - центр автономной нервной системы, влияющий на сохранение динамического постоянства внутренней среды организма и на ее приспособление к условиям внешней среды. Он посылает реципрокные импульсы к соответствующим ядрам среднего, продолговатого и спинного мозга, регулируя их деятельность. Впереди серого бугра зрительные нервы (II) образуют перекрест (хиазму), после которого получают название зрительных путей.
Над перекрестом лежит супраоптическое ядро. Его нервные клетки, как и клетки паравентрикулярного ядра, расположенного в стенке III желудочка, вырабатывают нейросекрет, проникающий в заднюю долю гипофиза. Зрительные пути, обогнув ножки мозга, делятся на медиальный и латеральный корешки. Медиальный направляется к верхнему двухолмию и зрительному бугру, латеральный (больший) идет к латеральному коленчатому телу.
Надбугорная область (epithalamus), связанная с обонятельными и рядом других импульсов, мала. Она состоит из мозговых полосок, сзади расширяющихся в поводковые треугольники (рис. 125). От последних отходят назад белые тяжики - поводки, соединяющие их с шишковидным телом, или эпифизом. Нижней стороной эпифиз прилежит к загибающейся назад задней спайке. Нисходящие волокна поводковых ядер, лежащих в одноименных треугольниках, идут к ядрам среднего мозга.
Третий желудочек имеет вид вертикальной - щели (рис. 125, 127). Его боковые стенки образованы медиальными поверхностями зрительных бугров и подбугорной области. Последняя ограничивает желудочек и снизу. В состав его задней стенки входит задняя спайка, под которой расположено отверстие водопровода. Переднюю стенку желудочка образуют колонки свода и проходящая спереди от них передняя спайка, относящиеся к конечному мозгу. Между колонками свода и зрительными буграми расположены межжелудочковые отверстия, соединяющие полость третьего желудочка с боковыми желудочками больших полушарий. Верхняя стенка третьего желудочка состоит из эпителиальной пластинки (остаток стенки мозгового пузыря) и сосудистой оболочки, образующих сосудистое сплетение третьего желудочка.
Поверхность полушарий. Межуточный мозг вместе со стволовой частью мозга прикрыт сверху и с боков большими полушариями - конечным мозгом. Полушария, как уже указывалось, состоят из подкорковых узлов (ганглиев) плаща, белого вещества и имеют полости - боковые желудочки.
Плащ у человека представлен корой, т. е. пластинкой серого вещества, отделенной от полости желудочков белым веществом. Это в основном новая кора, которая почти полностью вытесняет кору низших позвоночных; последняя остается у человека лишь в виде небольших участков древней и старой коры. Прогрессивно развиваясь у млекопитающих, новая кора достигает у человека огромных размеров, сложного строения и подчиняет себе функции всех нижерасположенных отделов мозга.
На всем своем протяжении кора углубляется в многочисленные борозды, которые делят всю поверхность полушария на выпуклые извилины и доли (Атл., 86 и 87). Различаются шесть долей: лобная, теменная, височная, затылочная, лимбическая и островок; их разделяют боковая, центральная, теменно-затылочная, поясная и коллатеральная борозды.
Боковая борозда (sulcus lateralis) начинается у основания полушария значительным углублением, дно которого образует покрытый бороздами и извилинами островок (рис. 129). Боковая борозда выходит на латеральную поверхность полушария, по которой идет полого назад и вверх, отделяя височную долю от вышерасположенных долей: лобной - спереди и теменной - сзади. От начала боковой борозды отходят две ветви: вверх - восходящая, и вперед - передняя (Атл., 87, А).
Центральная борозда (sulcus centralis) обычно прорезает медиальный край полушария приблизительно на середине его протяжения и тянется по верхнелатеральной поверхности вниз и вперед, не доходя обычно до боковой борозды. Центральная борозда отделяет лобную долю от теменной (Атл., 86, А).
Теменно-затылочная борозда (sulcus parieto-occipitalis) проходит вертикально по медиальной поверхности полушария, отделяя теменную долю (предклинье) от затылочной. На верхнелатеральной поверхности полушария границей между этими долями считается воображаемая линия, соединяющая верхний конец теменно-затылочной борозды с предзатылочной вырезкой (след давления на нижний край полушария пирамиды височной кости).
Поясная борозда (sulcus cingnli) проходит по медиальной поверхности полушария параллельно мозолистому телу, отделяя лобную и теменную доли от лимбической. Борозда оканчивается вертикальной ветвью (Атл., 87, Б).
Коллатеральная борозда (sulcus collateralis) отделяет на нижней поверхности полушария височную долю от лимбической и затылочной (Атл., 86, Б).
На нижней поверхности полушария, в его передней части, расположено овальное белое тело - обонятельная луковица, которая продолжается в обонятельный путь. Сзади он раздваивается на латеральную и медиальную обонятельные полоски, позади которых лежит переднее продырявленное вещество.
Доли полушария. Лобная доля в своей задней части, между центральной бороздой и расположенными впереди и параллельно ей верхней и нижней переднецентральными бороздами, имеет переднюю центральную извилину. Последняя на медиальной поверхности полушария переходит в переднюю часть парацентральной дольки. От обеих переднецентральных борозд отходят вперед почти под прямым углом две параллельные борозды - верхняя лобная и нижняя лобная, которые обособляют три лобные извилины. Верхняя лобная извилина заходит на медиальную поверхность полушария, где снизу ограничивается поясной бороздой. Средняя лобная извилина расположена между верхней и нижней лобными бороздами. Нижняя лобная извилина делится на три части: покрышковую часть (между нижней переднецентральной бороздой сзади, нижней лобной бороздой сверху и восходящей ветвью боковой борозды спереди), треугольную часть (между восходящей и передней ветвями боковой борозды) и глазничную часть (книзу от передней ветви боковой борозды), которая распространяется на нижнюю поверхность лобной доли. Наиболее выдающуюся вперед часть лобной доли называют лобным полюсом.
Теменная доля в своей передней части, между центральной и параллельной ей задней центральной бороздой, содержит заднюю центральную извилину, которая на медиальной поверхности полушария переходит в заднюю часть парацентральной дольки. Перпендикулярно задней центральной борозде, назад и параллельно медиальному краю полушария идет межтеменная борозда, разделяющая заднюю часть теменной доли на верхнюю и нижнюю теменные дольки. Верхняя долька заходит на медиальную поверхность полушария - это предклинье. В нижнюю теменную дольку врезаются задние концы боковой борозды и лежащей под ней верхней височной борозды. Часть этой дольки, окружающая конец боковой борозды, называется надкраевой извилиной, а часть, окружающая конец верхней височной борозды, - угловой извилиной.
Височная доля прорезается тремя продольными бороздами, параллельными боковой. Верхняя височная борозда лежит непосредственно под боковой, ограничивая вместе с ней верхнюю височную извилину. Средняя височная борозда, состоящая обычно из отдельных отрезков, ограничивает снизу среднюю височную извилину. Нижняя височная борозда, расположенная уже на нижней поверхности полушария, ограничивает нижнюю височную извилину с медиальной стороны. Между нижней височной бороздой и проходящей параллельно ей коллатеральной бороздой располагается латеральная височно-затылочная извилина. Спереди височная доля закругляется в височный полюс.
Затылочная доля кзади суживается в затылочный полюс. На верхнелатеральной поверхности она не имеет постоянных борозд, ее основная, шпорная борозда расположена горизонтально на медиальной поверхности и идет от затылочного полюса до теменно-затылочной борозды, в которой сливается в один ствол. Между этими бороздами лежит треугольной формы извилина - клин. Шпорная борозда и задняя часть коллатеральной ограничивают медиальную височно-затылочную извилину.
Лимбическая доля располагается на медиальной и нижней поверхностях полушария. Она состоит из поясной и парагиппокамповой извилин. Поясная извилина ограничивается снизу бороздой мозолистого тела, а сверху - поясной бороздой, отделяющей ее от лобной и теменной долей. Парагиппокампова извилина ограничивается сверху гиппокамповой бороздой, служащей продолжением вниз и вперед заднего конца борозды мозолистого тела. Снизу извилина отделена коллатеральной бороздой от височной доли.
Передний конец парагиппокамповой извилины охватывает передний конец гиппокамповой борозды, образуя крючок.
Базальные ганглии, или подкорковые узлы, - образования филогенетически более ранние, чем кора. К ним относятся хвостатое и чечевицеобразное ядра, объединяемые в полосатое тело, ограда и миндалевидное тело.
Хвостатое ядро (nucleus caudatus) вытянуто в сагиттальной плоскости и сильно изогнуто (рис. 125 и 130). Его передняя, сильно утолщенная часть - голова - помещается впереди зрительного бугра, в боковой стенке переднего рога бокового желудочка, сзади оно постепенно суживается и переходит в хвост.
Хвостатое ядро охватывает зрительный бугор спереди, сверху и с боков.
Чечевицеобразное ядро (nucleus lentiformis) располагается кнаружи от зрительного бугра, на уровне островка. Форма ядра близка к трехгранной пирамиде, обращенной своим основанием кнаружи. Ядро отчетливо делится прослойками белого вещества на более темноокрашенную латеральную часть - скорлупу и медиальную - бледный шар, состоящий из двух члеников: внутреннего и наружного (рис. 128 и 129).
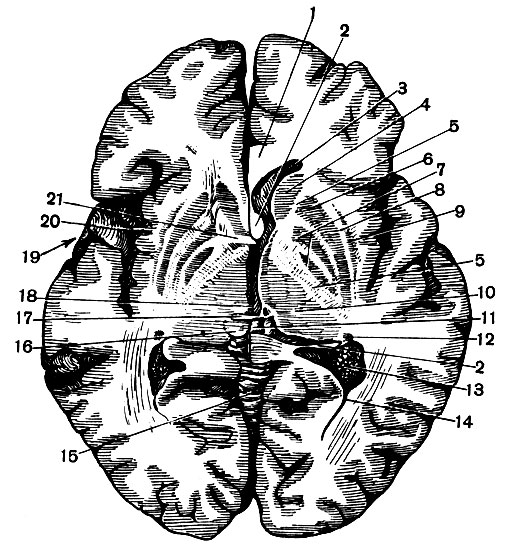
Рис. 129. Горизонтальный разрез через полушарие на уровне чечевицеобразных ядер (справа ниже, чем слева): 1 - мозолистое тело; 2 - свод; 3 - передний рог бокового желудочка; 4 - голова хвостатого ядра; 5 - внутренняя капсула; 6 - скорлупа; 7 - бледный шар; 8 - наружная капсула; 9 - ограда; 10 - зрительный бугор; 11 - эпифиз; 12 - хвост хвостатого ядра; 13 - сосудистое сплетение бокового желудочка; 14 - задний рог бокового желудочка; 15 - червь мозжечка; 16 - четверохолмие; 17 - задняя спайка; 18 - полость третьего желудочка; 19 - яма боковой борозды; 20 - островок; 21 - передняя спайка
Скорлупа (putamen) по генетическим, структурным и функциональным признакам близка к хвостатому ядру. Оба эти образования имеют более сложное строение, чем бледный шар. К ним подходят волокна главным образом от коры больших полушарий и зрительного бугра (Атл., 88).
Бледный шар (globus pallidus) в основном связан с проведением импульсов по многочисленным нисходящим путям в нижерасположенные структуры мозга - красное ядро, черную субстанцию и др.
Полосатое тело (corpus striatum), объединяющее хвостатое и чечевицеобразное ядра, относится к эфферентной экстрапирамидной системе.
В процессе филогенеза эти ядра надстроились над ядрами среднего мозга. Получая импульсы от зрительного бугра (Атл., 88), полосатое тело принимает участие в осуществлении таких сложных автоматических движений, как ходьба, лазанье, бег. В ядрах полосатого тела замыкаются дуги сложнейших безусловных, т. е. прирожденных, рефлексов. Экстрапирамидная система филогенетически более древняя, чем пирамидная. У новорожденного последняя еще недостаточно развита и импульсы к мышцам доставляются от подкорковых узлов по экстрапирамидной системе. Вследствие этого движения ребенка в первые месяцы жизни характеризуются обобщенностью, недифференцированностью. По мере развития коры больших полушарий волокна их клеток подрастают к базальным ганглиям и деятельность последних начинает регулироваться корой. Подкорковые узлы связаны не только с двигательными реакциями, но и с вегетативными функциями - это высшие подкорковые центры автономной нервной системы.
Хвостатое ядро, чечевицеобразное ядро и зрительный бугор отделены друг от друга белым веществом - внутренней капсулой.
Ограда (claustrum) - тонкая пластинка серого вещества, прилегающая снаружи к скорлупе, от которой отделяется тонким слоем белого вещества - наружной капсулой (рис. 128 и 129). Таким же тонким слоем белого вещества ограда отделена и от коры островка.
Миндалевидное тело (corpus amygdaloideum) - скопление клеток в белом веществе височной доли. При помощи передней спайки оно соединяется с одноименным телом другой стороны. Миндалевидное тело принимает импульсы из разнообразных афферентных систем, в том числе обонятельной, имеет отношение к эмоциональным реакциям.
Белое вещество располагается под корой больших полушарий, образуя выше мозолистого тела сплошную массу. Ниже белое вещество прерывается скоплениями серого (подкорковыми узлами) и располагается между ними в виде прослоек, или капсул (рис. 129). Самая мощная из них - внутренняя капсула - является продолжением основания ножек мозга (рис. 128) и состоит из проекционных путей, как восходящих, так и нисходящих (Атл., 88, 93).
Наиболее крупным нисходящим проекционным путем является пирамидный, состоящий из корково-ядерного и корково-спинномозгового путей.
Между внутренней капсулой и корой проекционные пути располагаются веерообразно, образуя лучистый венец.
В составе белого вещества различают ассоциационные, комиссуральные и проекционные волокна (рис. 131).
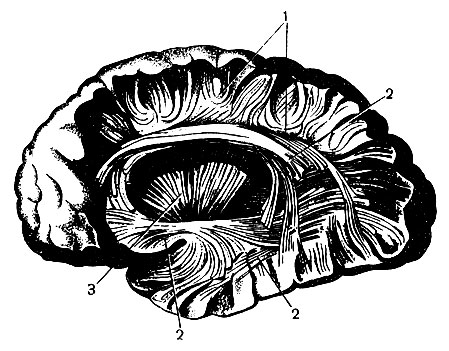
Рис. 131. Системы волокон в большом полушарии: 1 - коротких ассоциационных, или дугообразных; 2 - длинных ассоциационных; 3 - проекционных. Препарат приготовлен методом расщипывания
Ассоциационные волокна связывают различные участки коры одного и того же полушария. Короткие волокна проходят на дне борозд и соединяют кору соседних извилин, а длинные - извилины различных долей.
Комиссуральные волокна связывают кору симметричных частей обоих полушарий (Атл., 88). Мозолистое тело - самая крупная комиссуральная система, соединяющая одноименные участки новой коры. Для филогенетически ранней коры такой системой служат передняя и задняя спайки и др.
Проекционные волокна выходят за пределы полушарий в составе проекционных путей. По ним осуществляется двусторонняя связь коры с нижележащими отделами центральной нервной системы, вплоть до спинного мозга (Атл., 88 и 93).
Боковые желудочки. Боковой желудочек полушария состоит из средней части и трех отходящих от нее рогов (рис. 130).
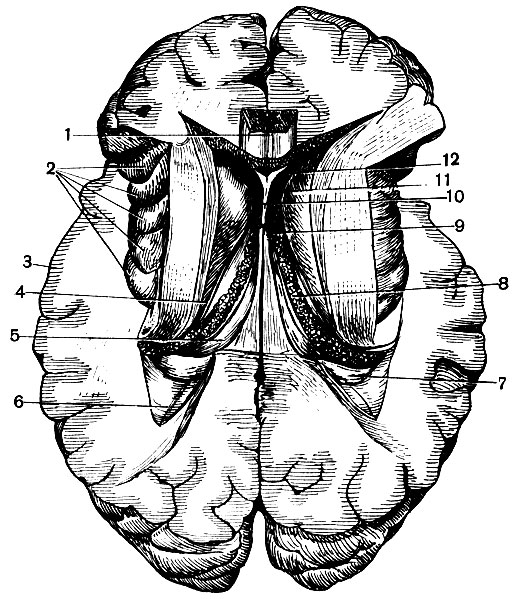
Рис. 130. Боковые желудочки (верхняя часть полушарий удалена): 1 - мозолистое тело; 2 - островок; 3 - кора; 4 - хвост хвостатого ядра; 5 - свод; 6 - задний рог бокового желудочка; 7 - гиппокамп; 8 - сосудистое сплетение; 9 - межжелудочковое отверстие; 10 - прозрачная перегородка; 11 - голова хвостатого ядра; 12 - передний рог бокового желудочка
Средняя часть в виде узкой горизонтальной щели расположена на уровне теменной доли, над зрительным бугром, и содержит сосудистое сплетение бокового желудочка.
Передний рог, имеющий треугольное сечение, помещается в лобной доле. От переднего рога другого полушария он отделен прозрачной перегородкой, расположенной между мозолистым телом и колонкой свода. Сзади средняя часть бокового желудочка значительно расширяется и переходит в задний и нижний рога.
Задний рог, углубляющийся в затылочную долю, мал; на его внутренней стенке помещается выступ - птичья шпора, - образованный вдавливанием глубокой шпорной борозды; нижняя стенка также несколько приподнята коллатеральной бороздой.
Нижний рог проходит в толще височной доли вперед и вниз. Кто дно приподнято коллатеральной бороздой; медиальная стенка сильно впячена вглубь гиппокамповой бороздой и образует здесь гиппокамп. Последний состоит из сместившейся в желудочек старой коры низших позвоночных. Получая импульсы от многих анализаторов, он связан с регуляцией обобщенных движений всего тела и эмоциями.
Межжелудочковые отверстия открываются в боковые желудочки между их средней частью и передним рогом. Через эти отверстия переходят друг в друга сосудистые сплетения третьего и обоих боковых желудочков.
Архитектоника коры. Под архитектоникой коры больших полушарий понимают особенности ее микроскопического строения. Различают цитоархитектонику (совокупность особенностей клеточного строения) (рис. 132) и миелоархитектонику (совокупность особенностей волокнистого строения коры). Начало изучения архитектоники относится к концу позапрошлого века, когда в 1782 г. Дженнари впервые обнаружил неоднородность строения коры в затылочных долях полушарий. В 1868 г. Мейнерт разделил поперечник коры полушарий на слои. В России первым исследователем коры был В. А. Бец (1874). Наиболее широко изучение архитектоники стало проводиться в XX в. и достигло своего расцвета в трудах советских нейроморфологов.
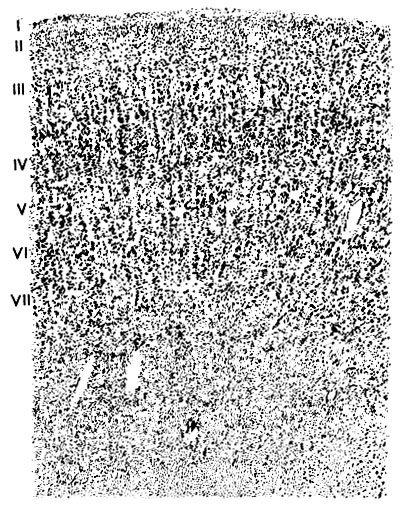
Рис. 132. Цитоархитектоника поля I больших полушарий человека (микрофотография с гистологического греза): I-VII - слои коры; ниже - белое вещество
Серое вещество, расположенное на поверхности больших полушарий, в основном новая кора, занимающая у человека 96%. Филогенетически более ранние корковые структуры - древняя, старая и межуточная кора - у человека ограниченны.
Древняя кора представлена главным образом прозрачной перегородкой боковых желудочков, передним продырявленным веществом и латеральной обонятельной полоской.
К старой коре относятся медиальная обонятельная полоска, гиппокамп, крючок парагиппокамповой извилины и полоска серого вещества в глубине борозды мозолистого тела.
За исключением крючка поверхность парагиппокамповой извилины имеет строение, переходное от старой коры к новой, и выделяется в качестве межуточной коры. Подобная кора в очень незначительном количестве сохранилась у человека и между древней корой и новой.
Толщина новой коры равна в среднем 2-3 мм. Образующие ее нервные клетки и волокна расположены в семь слоев (рис. 133). Последние отличаются по ширине, густоте расположения, форме и величине клеток, направлению, густоте и толщине волокон.
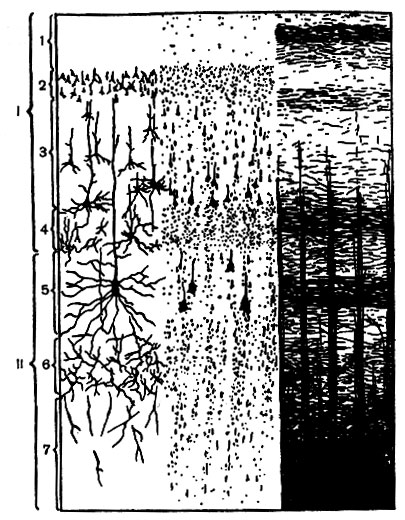
Рис. 133. Клеточное (слева) и волокнистое (справа) строение коры больших полушарий на поперечном срезе (схема). Этажи: I - верхний, II - нижний. Слои: 1 - молекулярный; 2 - наружный зернистый; 3 - пирамидный; 4 - внутренний зернистый; 5 - ганглиозный; 6 - полиморфный и 7 - веретенообразных клеток
Слой I - молекулярный, содержит немногочисленные, очень мелкие клетки и расположенные параллельно поверхности (тангенциально) волокна. Является общим для новой, старой, древней и межуточной коры.
Слой II - наружный зернистый, хотя и содержит преимущественно мелкие, неправильной формы пирамиды. Беден миелиновыми волокнами.
Слой III - пирамидный, состоит из пирамидных клеток, размеры которых увеличиваются по направлению вглубь.
Обычно они располагаются колонками, между которыми проходят проекционные волокна.
Слой IV - внутренний зернистый, образован часто расположенными звездчатыми клетками и густым скоплением горизонтально направленных миелиновых волокон.
Слой V - ганглиозный, заключает в себе пирамидные клетки (среди которых встречаются очень крупные) и лежащие радиально и горизонтально волокна.
Слой VI - полиморфный, с большим количеством веретенообразных клеток; отличается изменчивостью в распределении и густоте клеток и волокон.
Рис. 133. Клеточное (слева) и волокнистое (с прав а) строение коры больших полушарий на поперечном срезе (схема).
Этажи: I - верхний, II - нижний. Слои: 1 - молекулярный; 2 - наружный зернистый; 3 - пирамидный; 4 - внутренний зернистый; 5 - ганглиозный; 6 - полиморфный и 7 - веретенообразных клеток.
Слой VII имеет тот же клеточный состав, что и VI слой, но, являясь переходным к белому веществу, значительно беднее клетками и богаче волокнами.
Все клетки коры - вставочные нейроны рефлекторных дуг. По строению и характеру участия в деятельности коры среди них различают клетки с длинным и клетки с коротким нейритом.
Клетки с длинным нейритом представлены пирамидными и веретенообразными клетками. Это основные клеточные элементы V-VII слоев. Длинный нисходящий нейрит этих клеток отдает многочисленные коллатерали на всем своем пути и, выходя из коры, продолжается в белое вещество как нисходящее проекционное волокно. Последнее оканчивается в подкорковых ганглиях, двигательных ядрах ствола или на мотонейронах спинного мозга. Восходящий дендрит пирамидных клеток поднимается до первого слоя коры, образуя здесь густое конечное ветвление и отдавая, как и другие дендриты пирамидных нейронов, ветки в нижележащие слои. Таким образом, пирамидные клетки собирают импульсы со всех слоев коры.
В верхних слоях длинные нейриты имеют пирамидные клетки III слоя. Нейриты этих клеток входят в состав белого вещества преимущественно в качестве ассоциационных волокон, по которым осуществляется связь между различными участками коры, а также в виде комиссуральных волокон, связывающих кору двух полушарий.
Клетки с коротким нейритом отличаются тем, что последний не выходит за пределы коры. Эти клетки имеют главным образом звездчатую, а также паукообразную и кустовидную форму. У человека их больше, чем у животных, и они разнообразнее по форме. Клетки эти встречаются во всех слоях коры. В IV слое это главные элементы. Их роль заключается в восприятии афферентных импульсов и распределении их на пирамидные клетки III и V слоев. Звездчатыми клетками осуществляется, кроме того, круговая циркуляция импульсов в коре. Передавая импульс от одной звездчатой клетки к другой, эти нейроны объединяются в рефлекторные кольца. Восприняв нервный импульс, они могут длительно пребывать в состоянии скрытой, не выявляющейся во внешних реакциях активности и после того, как прекратилось действие раздражителя. Это и есть одна из форм материального субстрата памяти, анатомо-функциональные предпосылки для динамической фиксации следов возбуждений, удержания и эффективного использования информации, запасаемой человеком на протяжении всей его жизни.
Таким образом, по характеру составляющих слои нейронов кора может быть разделена на верхний и нижний этажи. Нижний этаж, представленный V-VII слоями, несет проекционную функцию, отдавая нисходящие волокна к двигательным ядрам головного и спинного мозга. Верхний этаж, состоящий из II-IV слоев, распространяет по коре импульсы, поступающие по восходящим волокнам от подкорковых структур, и посылает ассоциационные и комиссуральные волокна ко всем областям коры, т. е. имеет отношение к более сложным функциям. Верхний этаж коры онто- и филогенетически развивается позже нижнего; у человека он выражен лучше, чем у животных.
Следует считать, что сложность и совершенство строения коры больших полушарий человека по сравнению с ее организацией у животных зависят главным образом от увеличения количества клеток с коротким нейритом, а значительное увеличение площади коры человека связано с ростом ассоциационных волокон.
По ширине слоев, форме, величине и густоте расположения клеток кору делят на области и поля (рис. 134). Области появляются в течение развития раньше и характеризуются более общими признаками, чем поля, выделяющиеся в результате позднейшей, более дробной структурной дифференцировки.
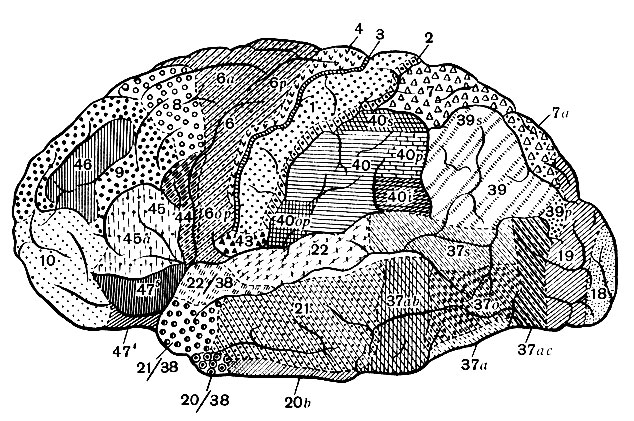
Рис. 134. Карта цитоархитектонических Полей коры наружной поверхности больших полушарий (по данным Института мозга). Цифры обозначают ноля: цифры с буквами или цифровыми указателями подполя; цифры, разделенные косой линией, отмечают пони, заходящие на височный полюс
Затылочная, височная и островковая области совпадают с соответствующими долями полушария. Теменные, верхняя и нижняя, и заднецентральная области входят в состав теменной доли. Первые покрывают одноименные дольки и отделены от заднецентральной области заднецентральной бороздой. Переднецентральная и лобная области занимают лобную долю; граница между ними более или менее соответствует верхней и нижней переднецентральным бороздам. С поясной извилиной совпадает лимбическая область, входящая в состав лимбической системы.
Возникновение цитоархитектонических областей предшествует образованию борозд и извилин на поверхности полушарий. Причина образования борозд и извилин заключается в неравномерном росте отдельных частей коры, что влечет за собой смещение некоторых ее участков и возникновение на поверхности полушарий западений и выпячиваний.
Расположение борозд и извилин на поверхности полушария, распространение архитектонических структур, их относительная величина, форма и качественные особенности неодинаковы у людей. На этом основании некоторые буржуазные ученые строили выводы о превосходстве одной расы над другой. Исследование советскими учеными большого материала показало, что индивидуальные различия в соотношении борозд и извилин, в распределении, строении и величине архитектонических формаций коры настолько велики в пределах каждой расы, что ни один из этих признаков не может считаться типичным для той или иной расы.
Локализация функций. Кора больших полушарий является наиболее высокоорганизованной материей, с которой связаны вмещая нервная деятельность и регуляция функций всех органов. И. П. Павлов считал, что даже самые малые детали строения коры рано или поздно найдут свое объяснение в свете рефлекторной теории.
При изучении деятельности коры как моста сложнейшего анализа и синтеза разнообразных раздражений необходимо учитывать локализацию в ней функций. Уже Гален знал, что функции органов связаны с мозгом. Более ста лет назад Галль указал на зависимость психической деятельности от коры больших полушарий. Он говорил о связях между характером развития определенных участков мозга и внешней формой черепа. Его наивные умозрительные представления были первыми попытками приурочить функции организма к структуре мозга.
Взгляды названных ученых не помешали в дальнейшем еще в течение долгого времени считать кору однородной в структурном и функциональном отношении. Изучение архитектоники коры широко развернулось лишь в начале XX столетия. В первых работах, принадлежащих западноевропейским ученым (Бродман, Экономо, Фогт и др.), в противоположность прежним взглядам начали проводиться узколокализационистские воззрения и делались попытки отнести локализацию функций даже к отдельным архитектоническим полям.
И только трудами И. П. Павлова и его школы создано на основании экспериментов материалистическое учение о динамической локализации функций. По этому учению кора больших полушарий представляет собой совокупность мозговых концов анализаторов. Каждому периферическому рецепторному аппарату соответствует в коре область, которая названа И. П. Павловым ядерной зоной анализатора.
Ядерная зона двигательного анализатора, воспринимающего раздражения проприорецепторов суставов, скелетных мышц и сухожилий, расположена в переднецентральной и заднецентральной областях, главным образом в полях 3 и 4, где оканчиваются восходящие проекционные волокна вентрального ядра зрительного бугра. В поле 4 от гигантских пирамидных клеток V слоя, впервые описанных русским исследователем В. А. Бецом, начинается большинство волокон самых мощных нисходящих путей коры - корково-спинномозгового и корково-ядерного (Атл., 88). Оканчиваются волокна этих путей на клетках передних рогов спинного мозга и двигательных ядер черепных нервов.
Ядерная зона кожного анализатора, связанного с температурной, болевой и тактильной чувствительностью, занимает главным образом заднецентральную область (поля 3, 1, 2). Основная масса волокон, приходящих сюда из вентрального ядра зрительного бугра, оканчивается в поле 3.
Внутри корковых концов двигательного и кожного анализаторов происходит очень закономерное проецирование различных частей тела. Так, в участки обеих центральных извилин и парацентральной дольки, расположенных вблизи медиального края полушария, поступают импульсы от нижней конечности; в нижерасположенные участки извилин - импульсы от туловища; в еще более низкие - от верхней конечности, и, наконец, в самые нижние части центральных извилин проецируются язык, гортань, глотка, лицо.
Ядерная зона зрительного анализатора находится в затылочной области (ноля 17, 18, 19). Основная масса волокон зрительной лучистости оканчивается в поле 17, на стенках и дне шпорной борозды.
Ядерная зона слухового анализатора располагается в височной области (поля 41, 42, 20, 21, 22). В полях 41 и 42 верхней височной извилины оканчивается большинство волокон слуховой лучистости.
Ядерная зона обонятельного анализатора связана с древней и старой корой обонятельного треугольника, прозрачной перегородки, крючка парагиппокамповой извилины, гиппокампа и др.
Помимо ядерных зон, или "проекционных" областей коры, воспринимающих импульсы главным образом от одной сенсорной или чувствительной системы, в коре больших полушарий обособляются другие, межанализаторные, так называемые ассоциативные области, принимающие импульсы от многих сенсорных систем. В них, например в теменных и лобной областях, происходит перекрытие ядерных зон анализаторов или проекций различных сенсорных систем и осуществляются высшие интегративные функции.
Участки проекционных областей коры, в которых оканчивается главная масса восходящих волокон анализаторов (рис. 135), выделяются как центральные, или первичные, поля. Они характеризуются специфическим строением. Так, кора полей 3, 17, 41 отличается исключительной многоклеточностью, обилием мелких зернистых клеток, хорошо развитым IV слоем. Исключение представляет поле 4, занятое двигательным анализатором. Здесь зернистые клетки у взрослого человека рассеяны по всему поперечнику коры. Это связано с тем, что поле одновременно служит и началом мощной системы нисходящих волокон пирамидных корково-спинномозгового и корково-ядерного путей.
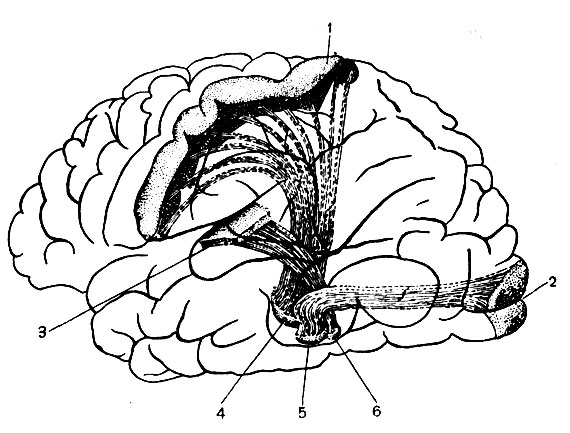
Рис. 135. Восходящие проекционные системы волокон к центральным нолям анализаторов или проекционным областям: 1 - задняя центральная извилина (кожный анализатор) приподнята чтобы показать входящие в нее нервные волокна; 2 - шпорная борозда (зрительный анализатор); 3 - верхняя височная извилина (слуховой анализатор); 4 - вентральное ядро зрительного бугра; 5-6 - латеральное и медиальное коленчатые тела
Если главная масса восходящих к проекционной области волокон оканчивается в ее центральном поле, то к ее периферическим, или вторичным, полям 1 и 2, 18 и 19, 22 и др. подходит меньше афферентных волокон, чем к центральному, но, помимо афферентных, подходят и волокна от центрального поля. В периферических полях зернистых клеток значительно меньше и преобладают пирамиды различных размеров.
Описанные структурно-функциональные особенности коры И. П. Павлов относит к первой сигнальной системе действительности, общей у человека с животными.
Помимо полей, хорошо выраженных и в коре мозга животных, у человека достигает мощного развития ряд других, филогенетически молодых полей. Примером может быть поле 40 нижнетеменной области, занимающее надкраевую извилину. С ним связано регулирование выработанных в течение жизни и направленных к определенной цели движений. Поражен не его ведет к утрате способности выполнять сложнокоординированные двигательные акты.
Локализация речевых функций. В процессе исторического развития человеческого общества в коре больших полушарий мозга усовершенствовались структурно-функциональные особенности, стоящие в зависимости от трудовой деятельности и связанной с ней речи.
Такие структуры коры мозга человека, которые воспринимают возбуждение, приходящее от речевых органов, относятся уже ко второй сигнальной системе действительности. Это поля 44 и 45, занимающие покрышковую и треугольную части нижней лобной извилины. В своей деятельности они тесно связаны с нижней частью переднецентральной извилины, в которую поступают проприорецептивные импульсы от мышц языка, губ, щек и гортани. Переключившись здесь на новые нейроны, импульсы по их отросткам поступают в поля 44 и 45. Последние описываются как речедвигательный анализатор. Этот участок коры, связанный с устной речью, расположен, следовательно, впереди того отдела двигательного анализатора (поле 4), в котором оканчиваются пути, приносящие импульсы от головы. Расположенная в заднем отделе средней лобной извилины часть ноля 6 связана с письменной речью и лежит впереди того отдела поля 4, который принимает проприорецептивные импульсы от руки. Совместная деятельность этих ядерных зон связана со сложными двигательными актами, необходимыми в письменной речи. Повреждение этой части поля 6 приводит к нарушению тонких движений руки, производимых при начертании букв. Другие участки коры, особенно тесно связанные с речью, сформировались недалеко от анализаторов зрения и слуха. С полем 39, занимающим угловую извилину нижнетеменной области и тесно примыкающим к ядерной зоне зрительного анализатора, связано зрительное восприятие письменных знаков. При поражении поля 39 утрачивается способность слагать из букв слова и фразы. В поле 22, расположенном в задней части верхней височной извилины, при участии полей 41 и 42 (ядерная зона слухового анализатора), происходит слуховое восприятие речи. При разрушении этого участка поля 22 теряется способность понимать слова.
Филогенетически новые участки коры, особенно тесно связанные с речью, асимметричны и у правшей представлены в левом, у левшей - в правом полушарии.
В настоящее время показано, что и второе полушарие небезразлично к речевым функциям (воспринимает интонации голоса и придает интонационное окрашивание речи). Специализация полушарий проявляется, кроме того, в характере организации памяти и в регуляции эмоциональных состояний.
Наличие у человека полей, разрушение которых ведет к выпадению речевых функций, не значит, что последние связаны только с определенными участками коры. Здесь, как и при локализации в первой сигнальной системе, некоторые поля имеют лишь преобладающее значение. Речь наиболее сложно локализована и осуществляется при участии всей коры. В соответствии с выработкой нового опыта речевые функции могут перемещаться в другие области коры (чтение слепых, письмо йогой у безруких и т. п.).
Возрастные изменения коры. В течение развития от пяти месяцев зародышевой жизни до взрослого поверхность коры увеличивается примерно в 30 раз. Отдельные области коры до рождения и после него растут неодинаково, что связано с их филогенетическим происхождением и функциональными особенностями.
Помимо обонятельного анализатора, связанного в основном с древней корой, в новой коре раньше других приближаются к строению мозга взрослого корковые концы двигательного и кожного анализаторов, а также лимбическая система. Лимбическая система включает лимбическую область коры, гиппокамп, сосцевидные тела, передние таламические ядра и др. Она имеет большое значение в обеспечении сложного комплекса разнообразных эмоциональных реакций. Лимбическая система связана с интерорецепторами и регуляцией секреции эндокринных желез и поддерживает гомеостаз в организме. Затем дифференцируются корковые концы зрительного и слухового анализаторов и ассоциативная верхнетеменная область, имеющая отношение к тонкой кожной чувствительности - узнаванию предметов на ощупь. При этом на протяжении всего внеутробного развития относительная величина одной из этих более старых областей - затылочной - сохраняется постоянной (12%). Значительно позднее приближаются к строению мозга взрослого такие эволюционно новые, ассоциативные области, как лобная и нижнетеменная, связанные с несколькими анализаторами. При этом, в то время как у новорожденного лобная область составляет 20,6-21,5% поверхности всего полушария, у взрослого она занимает 23,5%. Нижнетеменная область занимает у новорожденного 6,5% поверхности всего полушария, а у взрослого - 7,7%. Филогенетически наиболее новые ассоциативные поля 44 и 45, "специфически человеческие", имеющие преимущественное отношение к речедвигательному анализатору, дифференцируются на более поздних этапах развития, что продолжается и после семи лет.
В процессе развития увеличивается ширина коры в 2,5-3 раза. Прогрессивно растут и отдельные ее слои, особенно слой III, и наиболее интенсивно в ассоциативных полях коры. В течение развития наблюдается уменьшение числа клеток на единицу площади, т. е. их более разреженное расположение. Это связано со значительным ростом и усложнением отростков нервных клеток, особенно дендритов, рост которых ведет к раздвиганию тел нейронов.
Различные области коры миелинизируются в онтогенезе неодновременно. Первыми в последние месяцы зародышевой жизни получают миелиновую обкладку волокна проекционных областей, в которых оканчиваются восходящие или берут начало нисходящие корковые пути. Ряд путей миелинизируется в течение первого месяца после рождения. И наконец, на втором - четвертом месяцах жизни этот процесс охватывает наиболее филогенетически новые области, развитие которых особенно характерно для полушарий конечного мозга человека. Тем не менее кора полушарий ребенка в отношении миелинизации еще значительно отличается от коры взрослого.
Большой скачок в степени зрелости коры мозга ребенка по сравнению с корой мозга новорожденного наблюдается через 14 дней после рождения.
Особенно интенсивно увеличивается поверхность полушарий и их отдельных областей в первые два года жизни. Это связано с формированием сложных, целенаправленных действий, быстрым развитием речи и первыми признаками становления абстрактного мышления. Дальнейшее качественное совершенствование коры больших полушарий и изменение количественных показателей особенно резко выявляются в 4 года и 7 лет, когда процессы психической деятельности становятся богаче, разнообразнее и сложнее. Возраст 7 лет можно считать критическим в развитии ребенка как по морфологическим данным, так и физиологическим показателям.
При использовании материалов сайта активная ссылка обязательна:
http://anfiz.ru/ 'Анатомия и физиология человека'