Активирующие и тормозные условные реакции нейронов
Наиболее существенной особенностью условного рефлекса на клеточном уровне следует считать способность нейронов вырабатывать на положительный сигнальный стимул как активирующие условные реакции с повышением частоты разрядов нейрона, так и тормозные условные реакции с угнетением активности клетки.
На эту особенность было обращено внимание уже в первых микроэлектродных исследованиях условнорефлекторных связей. Так Jasper с сотрудниками (1960) в процессе выработки у обезьян оборонительных условных рефлексов на свет зарегистрировали в двигательной и соматосенсорной коре 77% клеток с активирующими и 23% клеток с тормозными условными реакциями. В лобной коре соотношение было противоположным: активирующие условные реакции проявило 38% клеток, а 62% нейронов выработали на положительный сигнальный стимул тормозные условные реакции. Примерно такое же относительное количество нейронов с активирующими и тормозными условными реакциями (35 и 65% соответственно) зарегистрировано в теменной коре.
Иное соотношение количества нейронов с тормозными и активирующими условными реакциями в префронтальной коре (дорсо-медиальиый отдел) обезьян обнаружили Kubota и Niki (1971) Kubota с соавторами (1972), Niki с соавторами (1972) после предварительной выработки отсроченных (на 1-5 с) пищедобывательных условных рефлексов с поочередным нажатием на действующую (правую или левую) педаль подачи пищевого подкрепления. В такой экспериментальной ситуации 67% клеток префронтальной коры проявили активирующие и 33% клеток - тормозные условные реакции. Тормозные условные реакции у таких нейронов появлялись в период очередного пищедобывательного движения (нажатие на педаль); в период "задержки" эти клетки проявляли активацию, что послужило поводом предположить о связи их активности с механизмами отсроченного действия. В хвостатом ядре, дорсо-медиальном и вентро-латеральном ядрах таламуса нейронов с тормозными условными реакциями эти авторы не нашли; все зарегистрированные клетки проявляли только активирующие условные связи.
Olds с соавторами (1969) тренировали крыс нажимать на педаль подачи подкрепляющего стимула (пищевая таблетка или вода) и затем на протяжении 1,75 с задерживать движение до подачи кормушки. Даже небольшое движение животного в этот временной интервал автоматически отключало подачу подкрепления. В таком эксперименте, который представляет собой не что иное, как выработку условного рефлекса "на состояние" по П. С. Купалову (1963), 21% нейронов гиппокампа, гипоталамуса и зоны инцерта проявили стабильное снижение частоты разрядов в последнюю секунду периода неподвижного "ожидания" пищи или воды, т. е., вырабатывали тормозные условные реакции. Нейроны других исследованных образований (таламическая группа ядер, тегментум, ретикулярная формация среднего мозга) проявляли только активирующие условные реакции.
При выработке у крыс обычных пищедобывательных условных рефлексов на звук, по данным Olds и Hirano (1969), снижение частоты разрядов на сигнальный стимул проявили только нейроны дорсального отдела ретикулярной формации среднего мозга в первые 300 мс односекундного изолированного действия звука. В последующие 700 мс эти клетки повышали частоту разрядов и вся реакция в целом оценивалась авторами как активационная. Другие исследованные образования (гиппоками, гипоталамус, преоптическая область, таламическая группа ядер) реагировали на сигнальный стимул изначальной активацией.
Неожиданное отсутствие у большого ряда важнейших подкорковых образований мозга клеток с тормозными условными реакциями Olds объясняет тем обстоятельством, что в использованных экспериментальных: ситуациях активация нейронов связана не с двигательными актами, невозможными без участия процессов торможения, а с мотивационными факторами (потребность в пище или воде) и памятью о них.
В литературе приводятся следующие крайние значения количества нейронов с тормозными условными реакциями: 9% клеток в ретикулярных структурах ствола мозга при выработке у обездвиженных кошек условных: реакций сочетаниями звуковых стимулов с электрокожным раздражением конечности (Yoshii, Ognra, 1960) и: 62% нейронов в лобной коре, 65% клеток в теменной коре при выработке у бодрствующих обезьян условных оборонительных рефлексов на вспышки света (Jasper e. a., 1960); 63% клеток в дорсальном отделе гиппокампа при выработке у бодрствующих кроликов двигательных условных рефлексов на время (Ф. В. Копытова, Л. К. Куликова, 1970).
Надо полагать, что соотношения нейронов с активирующими и тормозными условными реакциями в каждом образовании мозга также определяются совокупностью таких факторов, как структурно-функциональная организация образования, организация синаптических входов для сочетаемых стимулов и механизмы их взаимодействия на клеточном уровне, природа подкрепляющего стимула и форма эксперимента.
По нашим данным, в процессе выработки у бодрствующих кроликов двигательных условных рефлексов сочетаниями звуковых стимулов с электрокожным раздражением конечности в моторной коре активирующие условные реакции выработали около 58% нейронов, а 42% клеток проявили тормозные условные реакции; в слуховой коре преобладали нейроны с тормозными условными реакциями - 55%, активирующие условные реакции выработало 45% клеток; во внутреннем коленчатом теле только 30% нейронов проявили тормозные условные реакции, 70% - активирующие (Е. Л. Полонская, М. Я. Рабинович, 1971, 1972; Rabinovicn, Polonskaya, 1973). Примеры нейронов с активирующей и тормозной условными реакциями на положительный сигнальный стимул представлены на рис. 15.
Таким образом, по данным большинства авторов, значительная часть условнореагирующих нейронов в различных структурах мозга проявляет тормозные условные реакции на положительный сигнальный стимул. Поскольку такого рода активность вырабатывается в процессе сочетаний, ее следует рассматривать как одну из форм внутреннего торможения наряду с дифференцировками и угашением.
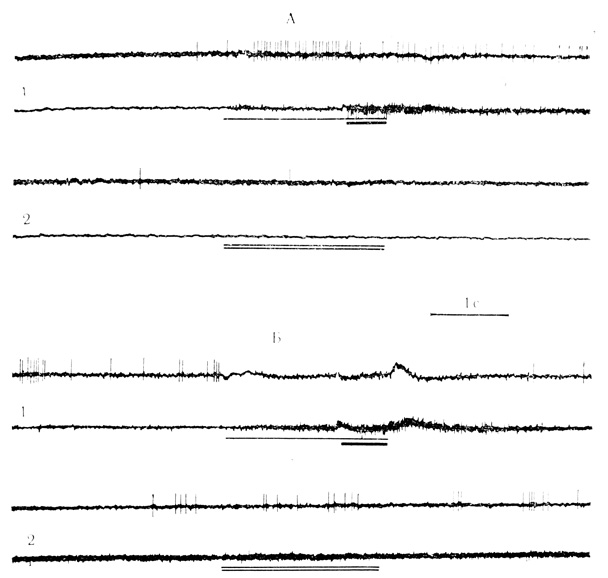
Рис. 15. Активирующая (А) и тормозная (Б) условные реакции нейронов двигательной (коры на положительный сигнальный стимул. А - клетка на глубине 1070 мкм; 501-е сочетание (1) и 30-е предъявление дифференцировочного стимула (2); Б - клетка на глубине 1400 мкм; '450-е сочетание (1) и 125-е предъявление дифференцировочного стимула (2). Условный стимул (щелчки 20 в 1 с) отмечен тонкой линией на каждой оспиллограмме, подкрепление (электрокожное раздражение конечности) - толстой линией, дифференцировочный стимул (щелчки 10 в 1 с) - двумя тонкими линиями
Можно полагать, что при выработке локального поведенческого акта нейроны с тормозными условными реакциями на положительные стимулы осуществляют координационное торможение, ограничивая с первых же сочетаний возбудительные процессы в пределах соответствующей популяции клеток. Такое изначальное развитие процессов внутреннего торможения в зонах проекции сигнального и подкрепляющего стимулов способствует формированию механизмов афферентной и эфферентной специализации приспособительного поведения животного. В этом, по-видимому, и заключается функциональное значение торможения активности значительной части нейронов при воздействии положительных условных сигналов.
Нашу точку зрения (М. Я. Рабинович, 1969, 1970) на тормозные условные реакции нейронов как составную часть клеточных механизмов формирования внутреннего торможения разделяет Э. А. Асратян (19726).
При использовании материалов сайта активная ссылка обязательна:
http://anfiz.ru/ 'Анатомия и физиология человека'