IV. Аппарат Гольджи
Обсуждение вопроса о реальности и значении "внутреннего сетчатого аппарата" Гольджи (ап. Гольджи) началось со времени его открытия в 1898 г. и продолжалось до последнего времени. Как известно, Гольджи, используя метод серебрения, описал в нейронах котенка и в клетках Пуркинье мозжечка совы окружающую ядро тончайшую внутриклеточную сеть из узловатых и анастомозирующих друг с другом нитей. Затем многочисленными исследователями было подтверждено наличие ап. Гольджи в разных клетках у различных животных. Последнему посвящено около 5000 работ, многие из которых сохранили свое значение до наших дней, например обзоры Хибберда и Б. П. Токина (Hibbard, 1945; Токин, 1963).
В последние годы в эту проблему существенный вклад внесли электронная микроскопия и биохимия; современные представления базируются на классических работах Дальтона и Феликса (Dalton, Felix, 1953-1954), изучавших эпителиальные клетки эпидермиса крысы, а также Шостранда и Ханзона (Sjostrand a. Hanzon, 1954), исследовавших экзокринные клетки панкреатической железы мыши.
Субмикроскопическая структура ап. Гольджи в различных клетках одно- и многоклеточных животных, а также растений обнаруживает принципиальное единство. Ап. Гольджи состоит из ограниченных мембраной пузырьков и цистерн, которые отличаются от таких же элементов эндоплазматической сети в основном тем, что имеют меньшие размеры, совершенно свободны от гранул и более тесно связаны между собой. По этой электронномикроскопической картине ап. Гольджи является легко распознаваемым органоидом клетки. На электроннограммах он выявляется в виде трех ультрамикроскопических структур (рис. 16). 1. Пакеты уплощенных мешков или цистерн, представляющихся на разрезе как совокупность параллельных пар мембран с гладкими стенками толщиной 200-250 Å. В центральных частях ширина просвета цистерн достигает 150 Å, концевые же их отделы часто образуют расширения. 2. Крупные вакуоли или довольно большие лакуны величиной от 0.05 до 2-3 мк, неправильной формы, находящиеся на периферии зоны Гольджи. Они ограничены гладкой мембраной, толщиной 40-90 Å. 3. Мелкие довольно плотные пузырьки от 300 до 600 Å, часто интимно связанные с мембранами цистерн, при слабых разрешениях микроскопа представляются мелкими гранулами. Во всех трех ультраструктурах ап. Гольджи отсутствуют гранулы Палада (макромолекулы РНК, рибосомы). В ап. Гольджи различных клеток всегда присутствуют два образования - пакеты двойных пластинок (уплощенных мешочков) и мелкие плотные пузырьки. Большие светлые вакуоли не всегда обнаруживаются. Толщина мембран, ограничивающих все ультраструктуры ап. Гольджи, в пределах одной клетки одинакова. Большинство авторов считает, что мембраны ап. Гольджи являются одинарными. Однако Ямамото (Yamamoto, 1963) в нервных клетках продемонстрировал при очень большом разрешении трехслойный комплекс, который по толщине соответствует поверхностной мембране клетки.
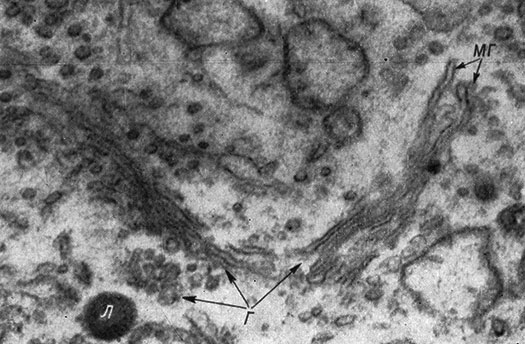
Рис. 16. Электронная микрофотография аппарата Гольджи нервной клетки. (Ориг.)
В ряде последних работ отмечаются интимные взаимоотношения ультраструктур ап. Гольджи с другими компонентами клетки, образующими прямые анастомозы с эндоплазматической сетью (Palade, 1959a, 1959b; Manton, 1960; Freeman, 1962; Novikoff, 1962).
Новиков и Эсснер (Novikoff, Essner, 1962) считают мелкие пузырьки ап. Гольджи "транспортными" структурами, обеспечивающими динамическую связь его с эндоплазматической сетью.
Методом дифференциального центрифугирования удается выделить чистую фракцию ап. Гольджи, которая отличается от других внутриклеточных органоидов клетки (Shneider a. Ruff, 1954, De Duve etc., 1955). Биохимическое и рентгеноструктурное изучение ап. Гольджи показало, что все мембранные системы этого органоида построены как из фосфолипидов, так и из нелипидных компонентов (Kuff, Dalton, 1957, 1959). На ультратонких срезах мембраны ап. Гольджи в нейронах всегда отчетливо выявляются (рис. 12, 16). Наиболее тщательные и обширные работы по функциональным свойствам ап. Гольджи выполнены Д. Н. Насоновым и его сотрудниками (Nassonov, 1923, 1930; Насонов, 1924). Представление Д. Н. Насонова об этом важном клеточном органоиде, как обеспечивающем сепарацию и конденсацию веществ в самых различных клетках, не потеряло значения и в настоящее время, получив дальнейшее развитие в работах, выполненных на ультраструктурном уровне. Рядом исследователей было установлено, что в области цитоплазмы, где находится ап. Гольджи, непосредственно происходит накопление липидов во время и после их всасывания (Weiner, 1926; Palay, Karlin, 1959).
Чоу и Мик (Chou a. Meek, 1958) электронномикроскопически обнаружили, что фосфолипидные капельки в нейронах улитки Helix aspersa превращаются в ап. Гольджи в процессе фиксации. Однако аналогичные исследования Дальтона (Dalton, 1961) не подтвердили этих данных. Другие электронномикроскопические исследования (Sjostrand, Hanzon, 1954; Burgos a. Fawcett, 1955; Ferreira, 1959) представили доказательства участия an. Гольджи в секреторной активности клетки. Так, секреторные гранулы в эмбриональных клетках островков Лангерганса и клеток аденогипофиза возникают внутри ап. Гольджи. Тесную связь между последним и образованием липидов в клетках сальной железы продемонстрировал Пелей (Palay, 1958). Имеется немало указаний на участие ап. Гольджи в формировании нейросекреторных гранул клеток ганглия таракана и гипоталамуса лягушки (Bern et al., 1961; Murakami, 1962). Однако проблема функционального значения этого органоида полностью еще не выяснена.
В настоящее время имеются многочисленные работы, посвященные выяснению функции ап. Гольджи, которые освещены в обзорах Дальтона (Dalton, 1961) Каффа и Дальтона (Kuff a. Dalton, 1957), И. Токина (1963), В. Ф. Машанского (1965) и др.
Таким образом, можно заключить, что ап. Гольджи является важным компонентом клетки, участвующим в накоплении и конденсации продуктов, особенно липидов, воспринимаемых и вырабатываемых в других участках цитоплазмы, а затем поступающих в его элементы.
При использовании материалов сайта активная ссылка обязательна:
http://anfiz.ru/ 'Анатомия и физиология человека'