Глава XVI. Внутриорганные межтканевые взаимоотношения и влияние организма как целого на ткани
Теснейшие взаимоотношения между тканями в органе складываются уже с самого начала его образования. На протяжении всего органогенеза происходят согласованные изменения в формирующихся строительных материалах - тканях, из которых постепенно складывается орган. Морфогенетический процесс, осуществляющийся на протяжении эмбрионального развития, протекает на основе координированного взаимоиндуцирующего влияния одной тканевой закладки на другую. Лишь после такого индукционно-формативного воздействия эмбриональных тканевых закладок друг на друга возникают органы и дифференцируются ткани. Эмбриогенез, собственно, и состоит из цепей такого рода взаимодействий органотканевых закладок, что было подробно изучено Д. П. Филатовым и его учениками. Вопрос о специфическом взаимодействии эмбриональных зачатков, вызывающем формообразовательные процессы в развивающемся зародыше, был широко разработан немецкими учеными Ру и Шпеманом, положившими начало особой области науки - механике, или динамике развития. Большое значение корреляциям, то есть взаимно регулирующим влияниям в индивидуальном развитии организмов, придавал академик И. И. Шмальгаузен.
Рассмотрим для примера развитие кожи как органа в онтогенезе (рис. 73). Она возникает из двух эмбриональных зачатков - кожной эктодермы и кожной мезенхимы - в результате их взаимного формативно-индукционного воздействия. Кожная эктодерма представлена у зародыша позвоночного слоем плоских клеток, покрытых с поверхности еще одним слоем очень уплощенных клеток перидермы. Такую структуру кожная эктодерма сохраняет и после того, как оказывается подостланной более или менее густым скоплением неспецифической мезенхимы, возникающей путем выселения отдельных клеток из париетального листка спланхнотома, их размножения и расселения между кожной и другими эмбриональными закладками. С момента, когда дерматом начинает рассыпаться на клетки кожной мезенхимы, кожная эктодерма над ними перестраивается в многослойную структуру эмбрионального эпидермиса специфического строения с резко выраженным слоем базофильных палисадно расположенных базальных клеток и вышележащими слоями пузыревидных клеток. Кожная мезенхима усиленно пролиферирует (размножается) и густым потоком, состоящим из тесно прилегающих друг к другу короткоотростчатых клеток, распространяется из области бывшего дерматома в дорзальном, и особенно в вентральном направлении. При этом кожная мезенхима вклинивается между кожной эктодермой и мезенхимой париетального происхождения. Клетки последней лежат рыхло, они длинноотростчаты и ориентированы параллельно поверхности тела. Там, где теперь кожная эктодерма оказывается подостланной кожной мезенхимой, она перестраивается в кожный эпителий - эпидермис.
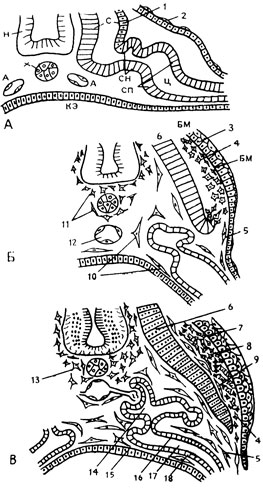
Рис. 73. Три стадии (А - Б - В) развития кожного покрова (по А. А. Брауну): Н - нервная трубка, Х - хорда, С - сомит, СН - сегментная ножка (нефротом), СП - спланхнотом, Ц - целомическая полость (вторичная полость тела). А - парная закладка аорты, КЭ - кишечная энтодерма, БМ - базальная мембрана на границе эпидермиса и дермы. 1 - дерматом; 2 - кожная эктодерма; 3 - эпидермис; 4 - кожная мезенхима; 5 - мезенхима, возникшая из висцерального листка спланхнотома; 6 - миотом; 7 - базальная мембрана; 8 - развивающаяся волокнистая дерма; 9 - мезенхима не дерматомного происхождения; 10 - мезенхима, возникшая из висцерального листка спланхнотома; 11 - скелетогенная мезенхима склеротомного происхождения; 12 - аорта; 13 - пролиферирующая скелетогенная мезенхима; 14 - формирующийся мочевой каналец; 15 - эпителий кишки; 16 - наружный листок целомического эпителия; 17 - целомическая полость; 18 - внутренний листок целомического эпителия
Под влиянием индуцирующего воздействия со стороны специально детерминированной кожной мезенхимы происходит превращение эмбриональной закладки в дифференцированную ткань. Процесс этот обоюдосторонний - при взаимодействии кожной мезенхимы с кожной эктодермой она дифференцируется в дермальную ткань со специфическим строением и расположением ее клеточных и волокнистых элементов. В париетальной мезенхиме, лежащей под кожной эктодермой, можно обнаружить лишь отдельные серебрящиеся преколлагеновые волокна. Формирующаяся соединительная ткань, согласно классификации Г. В. Ясвоина, находится еще на мезенхимальной стадии фибриллогенеза. Перестраиваясь в дерму, кожная мезенхима образует уже настоящие коллагеновые волокна, то есть переходит в следующую десмальную стадию развития и становится дифференцированной соединительной тканью. На границе между сложившимися тканями - эпидермисом и его физиологической подстилкой, дермой, как выражение установившихся между ними коррелятивных отношений, возникает аргирофильная базальная мембрана (рис. 73). Описанный процесс распространяется по обеим сторонам тела до тех пор, пока не произойдет смыкание строящихся из эмбриональных закладок дефинитивных тканей, закономерно объединяемых в орган - кожу.
Установившиеся внутри органов в эмбриональном периоде межтканевые корреляции сохраняются на протяжении всего онтогенеза. Органы являются не просто конгломератами тканей, а строгими их системами, в которых тканевые компоненты не только морфологически, по и функционально соподчинены друг другу, в целом работая на орган.
Основополагающая роль в разработке теории межтканевых корреляций во взрослом организме принадлежит школе академика А. А. Заварзина, а также В. Г. Гаршину. Предметом их исследований были взаимоотношения, главным образом, между пограничными тканями и тканями внутренней среды. Дальнейшее развитие теория межтканевых корреляций получила в работах А. Н. Студитского, развивающего идею о "тканевых регуляциях" на основании собственного большого фактического материала.
В своих ранних работах А. Н. Студитский выдвинул гипотезу о гистогенетических корреляциях, основанную на изучении взаимодействия тканевых компонентов развивающихся костей. Затем он расширил круг своих исследований в данном направлении, включив в них процессы постэмбрионального развития скелетных мышц за счет трансплантатов измельченной или предварительно денервированной мышечной ткани, а также процессы опухолеобразования, индуцированного введением целлофановых и других инертных прокладок между взаимодействующими тканями.
В. Г. Гаршин высказал идею об элиминативной функции эпителиев. Согласно его представлениям одним из гистиотипических свойств эпителиальных тканей всех типов является способность врастать в находящуюся под ними подстилку. В силу коррелятивных связей они не врастают лишь в свою физиологическую подстилку - подлежащую соединительную ткань, выполняющую по отношению к эпителиальному пласту трофическую функцию. Достаточно наличия хотя бы совсем небольшого слоя нормальной соединительной ткани с фибробластами, то есть сохранения нормальных корреляций между соединительной тканью и эпителием, чтобы воспрепятствовать образованию врастаний эпителия.
При попадании инородного тела во внутреннюю среду организма, в частности, в физиологическую подстилку эпителия, вокруг него, как вредности, создается очаг воспаления. Если зона воспаленной соединительной ткани, инфильтрированной лейкоцитами и другими блуждающими клетками, достигнет эпителия, подстилка его перестанет быть физиологической, в эпителиальном пласте начнутся процессы дедифференцировки и он проявит инфильтрирующий рост - образуются врастания, которые проникнут сквозь воспаленную соединительную ткань и остановятся, достигнув нормальной соединительной ткани. Инфильтрирующий рост тяжами перейдет в гистиотипический для эпителия рост пластом по здоровой соединительнотканой подстилке. По В. Г. Гаршину, это будет уже регенераторный рост эпителия.
В итоге языки регенераторно растущего эпителия смыкаются друг с другом и спаиваются с покровным его пластом, а над очагом воспаления в результате дифференцировки эпителия образуется прорыв. Инородное тело вместе с воспаленной соединительной тканью будет отграничено пластом эпителия от внутренней среды организма, то есть элиминировано из нее. Поскольку подобные разрастания эпителия связаны с устранением вредности из организма, В. Г. Гаршин назвал их воспалительными и в этом видел их биологическое значение.
А. А. Браун показал, что элиминативная функция может осуществляться эпителием как активно, так и пассивно. Активная элиминативная функция присуща лишь защитному, кожному эпителию и является его гистиотипическим свойством. Кишечный эпителий тоже может элиминировать инородное тело из внутренней среды, но лишь попутно, а не специально. Так, если в тело моллюска беззубки ввести целлоидиновую занозу и при этом рассечь стенку кишки, кишечный эпителий начнет регенерировать. Он растет пластом по соединительной ткани с обоих пораненных концов вплоть до смыкания. Инородное тело в итоге окажется лежащим в просвете кармана кишки и таким путем изолированным из внутренней среды организма. Однако элиминация инородного тела здесь только результат регенераторного роста и происходит пассивно. При более осторожном введении занозы, когда стенка кишки остается интактной, кишечный эпителий на инородное тело не реагирует и его элиминации из внутренней среды организма не происходит.
Иное дело когда, как установил Ф. М. Лазаренко, целлоидиновая заноза находится неподалеку от дыхательной трахеи в теле насекомого. Трахеи - это разветвленные трубочки кожного эпителия. В ответ на воспалительную реакцию вокруг занозы эпителий трахеи начинает активно расти к очагу воспаления, изолируя, элиминируя вредность из внутренней среды организма. Аналогичное поведение кожного эпителия было обнаружено Е. С. Данини у речного рака, а А. А. Заварзиным - у моллюска беззубки.
Очень наглядно удалось наблюдать активный, направленный рост кожного эпителия к очагу воспаления, возникающему (вокруг целлоидиновой занозы у лягушки. Если нанести лягушке сквозную рану кожи, дефект заполняется сгустком фибрина, проникающего сюда из подкожного лимфатического мешка. Из клеточных элементов в нем встречаются в первую неделю нейтрофилы, а затем лимфоидные клетки - те и другие в небольшом количестве. Эта фибриновая пробка очень скоро перекрывается пластом эпидермального регенерата и дальнейшие восстановительные процессы в ране протекают под его покровом. Затем фибриновый сгусток начинает заселяться фибробластами и постепенно перестраивается в плотную соединительную ткань, а эпидермальный покров успокаивается, утончается и уже мало отличается от нормального. Обычно эпителиальный регенерат не образует врастаний в подлежащий фибриновый сгусток или они выражены незначительно.
Однако между эпителиальным покровом формирующегося регенерата и его подстилкой уже на данном этапе устанавливаются определенные функциональные отношения и морфологическим выражением их служит провизорная аргирофобная (не проявляющая склонности к импрегнации серебром) базальная мембрана. Там, где она образовалась, эпидермис, точнее его базальный слой, принимает палисадный характер. Все же это не те межтканевые связи, которые имеют место в нормальной коже между эпидермисом и его физиологической подстилкой - соединительной тканью, ибо если создать под регенератом введением в подкожный лимфатический мешок целлоидиновой пленки очаг асептического воспаления, эпителиальный регенерат начинает образовывать врастания в фибриновый сгусток, которые доходят до инородного тела и элиминируют его вместе с клеточным инфильтратом из внутренней среды организма. Если же целлоидиновая заноза слишком велика и не может пройти через образующийся над ней пролежень в эпидермальном выросте, она все равно, оставаясь на месте, оказывается изолированной от внутренней среды лягушки защитным кожным эпителием.
А. А. Брауну удалось показать, что движение эпидермальных выростов к очагу воспаления является активным и направленным. Помещая под большим регенератом кожного эпителия малый кусочек целлоидиновой пленки в каком-нибудь краевом участке, всегда наблюдают врастания именно над ним. Эти врастания доходят до подкожного слоя, где залегает инородное тело, и элиминируют его. Можно убедиться специальными экспериментами, что дело здесь вовсе не в механическом воздействии целлоидиновой пленки на регенерат.
Если ставить опыт в обратном порядке - сначала ввести в лимфатический мешок занозу, подождать недели три, пока воспаление вокруг нее успокоится и целлоидиновая пленка оденется капсулой из плотной фиброзной ткани, фиксирующей ее на месте, а затем над бывшим очагом воспаления удалить кусочек кожи во всю ее толщину, никаких врастаний эпителия в фибриновый сгусток мы не обнаружим. Все как будто не изменилось, но наступила другая фаза воспаления и результат получился иной. По-видимому, когда вначале инородное тело оказывается окруженным лишь слоем лимфоидных клеток, от очага воспаления распространяются какие-то химические сигналы, которые действуют на молодой эпителий, лежащий на фибриновой пробке, и вызывают его активный и направленный рост к инородному телу, завершающийся элиминацией последнего из внутренней среды организма.
Наоборот, если внести целлоидиновую занозу под более старый регенерат, когда под эпителием уже залегают в фибрине ряды фибробластов и когда между эпителием и его физиологической подстилкой образовалась типичная аргирофильная базальная мембрана, никаких врастаний в фибрине не произойдет. Однако эпителий не остается безразличным к сигналам, идущим от очага воспаления, и начинает усиленно пролиферировать, утолщаться. Происходит, по терминологии В. Г. Гаршина, рост эпителия на месте.
С другой стороны, А. Н. Студитским была установлена необходимость постоянных взаимосвязей тканей в органе для нормального течения в нем формообразовательных процессов и жизнедеятельности. Нарушение же межтканевых корреляций может приводить к извращению морфогенеза и появлению злокачественных новообразований. Так, в лаборатории А. Н. Студитского вводили крысам между диафизом кости и эпифизом в области хрящевой пластинки роста целлофановую пленку. Через 14 - 16 месяцев в ряде случаев возникла остеосаркома, то есть происходило опухолевое перерождение остеогенной ткани.
В других опытах на крысах обертывали целлофановой пленкой (нарушающей межтканевые взаимодействия) икроножные мышцы, при сохранении нормальных сухожильных связей, иннервации и кровоснабжения. В 2 случаях из 10 через 14 и 16 месяцев возникли опухоли типа рабдомиобластомы, отличавшиеся чрезвычайной злокачественностью. Контрольные опыты с введением между мышцами равного количества измельченной целлофановой пленки показали, что в этих случаях наблюдается лишь воспалительная реакция, а злокачественного перерождения тканей не происходит.
Разного рода злокачественные новообразования возникали при использовании для нарушения взаимосвязи между тканями таких инертных материалов, как золотая или платиновая фольга.
Приведенные примеры достаточно убедительно показывают, как велико значение внутриорганных межтканевых корреляций. Они необходимы для нормального развития органов и для поддержания их сложившейся структуры.
Выше мы писали о том, что ткани, входя в состав органа, как бы подчиняются его "интересам", то есть подчиняются запросам со стороны системы высшего порядка, структурным компонентом которой они являются. Наивысшая целостная система - организм; его запросам отвечает и работа, а следовательно, и строение самих органов. Поэтому ткани, как строительные материалы, находятся в двойном подчинении - как со стороны органа, в конструкцию которого они вписаны, так и со стороны целого организма, который всегда остается единым при всей его расчлененности.
В сложившемся организме уже дифференцированные ткани детерминированы как представители определенных типов и вариации в их строении дальше внутритиповых модификаций, ограниченных природой этих тканей, не идут. Однако они могут претерпевать большую или меньшую гипертрофию либо, наоборот, атрофию, замещать, оттеснять в органе другие ткани или же, напротив, уступать "натиску" других тканевых компонентов органа. Наконец, может меняться их функциональное состояние, их общая жизнедеятельность либо специфическая рабочая производительность. Все это находится под контролем организма, как целого, и регулируется им нейрогуморальным путем. Нервная регуляция осуществляется из разных отделов центральной нервной системы или из ее периферических подстанций - нервных вегетативных узлов, управляющих работой внутренних органов.
Гуморальные влияния - это прежде всего гормональные воздействия. В организме, как мы указывали, имеется целая система желез внутренней секреции, вырабатывающих гормоны. Они характеризуются рядом общих свойств: во-первых, это вещества исключительно активного биологического действия, во-вторых, оказываемое ими влияние на различные органы и ткани весьма специфично и, в-третьих, их эффект реализуется на расстоянии от железы, в которой они вырабатываются. Особенностью гормонов является также их видовая неспецифичность. Это позволяет при недостаточности их у человека использовать гормоны, получаемые от животных разных видов. Химическая структура многих гормонов хорошо изучена, а некоторые из них уже научились синтезировать искусственно.
Наряду с эндокринными органами - гипофизом, надпочечниками, щитовидной железой и околощитовидными имеются еще железы со смешанной функцией - эндокринной и зкзокринной. Помимо упомянутой нами поджелудочной железы, сюда должны быть отнесены и половые железы - гонады (семенники и яичники), выполняющие, кроме своей прямой функции гаметообразования, и эндокринную. Регулируя обменные и морфологические процессы в организме, железы внутренней секреции сами находятся под влиянием нервной системы. В свою очередь, гормоны разных эндокринных органов оказывают действие на нервную систему. Например, при гиперфункции щитовидной железы человек становится чрезмерно подвижным и очень раздражительным, а при гипофункции ее - крайне медлительным и безразличным. Взаимовлияния нервной и эндокринной систем так тесны и многообразны, что обычно предпочитают говорить об единой нейроэндокринной регуляции жизненных процессов.
Филогенетически более древней является гуморальная координация, то есть взаимодействие между частями организма путем химической передачи сигналов через жидкости тела. Исторически позднее сформировалась нервная система, позволяющая быстро и строго направленно регулировать работу разных органов. Эндокринные железы явились еще более поздним филогенетическим приобретением, облегчающим деятельность нервной системы - импульсы, поступающие в органы внутренней секреции из нервной системы, генерализуются благодаря выбросу гормонов, которые попадают в кровь и разносятся до каждой клетки тела. Сигналы из нервной системы идут по нервным проводникам строго к определенным адресатам, сигналы из эндокринных органов распространяются по принципу: "Всем, всем, всем!". Эти сигналы воспринимают и отвечают соответствующими реакциями ткани и органы, на них настроенные.
Наиболее многочисленны гормоны, вырабатываемые гипофизом. Причем некоторые из них регулируют работу других эндокринных желез - щитовидной, надпочечников, половых. Вот почему гипофиз называют центральным в системе эндокринных органов, а другие железы внутренней секреции - периферическими. Большинство же гормонов гипофиза участвует в регуляции обменных процессов в организме, в регуляции его роста и развития. При увеличенной или уменьшенной выработке гормонов роста развиваются "гипофизарные" великаны и карлики, резко отличающиеся по росту от нормальных людей, но сохраняющие пропорциональность в строении тела.
Гормоны щитовидной железы, как мы уже указывали, являются важнейшими регуляторами основного обмена в организме. При гиперфункции этой железы он резко усиливается, человек быстро теряет в весе, температура тела его заметно повышается. Наоборот, при гипофункции щитовидной железы обмен веществ в организме более или менее сильно снижается, появляются отеки в тканях, а температура тела падает. Щитовидная железа оказывает влияние на формообразовательные процессы. Если ее убрать в период роста организма, он становится дефективным. У человека это приводит к кретинизму, а карлики, появляющиеся при отсутствии в развивающемся организме гормонов щитовидной железы, отличаются от "гипофизарных" лилипутов нарушениями пропорций тела.
Щитовидная железа вырабатывает еще один гормон - антагонист гормону околощитовидных желез. Вместе эти гормоны регулируют жизненно необходимый кальциевый обмен.
Надпочечные железы продуцируют ряд гормонов, также регулирующих разные формы обмена веществ. Кроме того, они сразу же включаются своими гормонами при всяком состоянии напряжения в организме, называемом стрессом, повышая общую его устойчивость; гормоны коры надпочечников играют ведущую роль в защитных приспособительных реакциях.
Гипофиз, регулирующий деятельность других желез внутренней секреции, находится под влиянием веществ - секретов, поступающих в него из близлежащего гипоталамуса - базальной части промежуточного мозга. Здесь расположены скопления особых невросекреторных клеток, которые являются одновременно и невронами, и железистыми элементами. Вырабатываемый ими невросекрет поступает по аксону в задний отдел гипофиза.
Наконец, следует иметь в виду, что гипоталамус связан со всеми отделами головного мозга, в том числе с наивысшим его центром - корой больших полушарий, а также с подкорковыми центрами и стволовой частью. Поэтому он является основной структурой, через которую происходит переключение импульсов из высших отделов центральной нервной системы ко всем эндокринным органам.
Все органы и ткани нашего тела находятся под регулирующим воздействием нервной системы. В опытах, проведенных нами на коже, было установлено, что под влиянием возбуждения центральной нервной системы тканевые реакции усиливаются, а торможение приводит к снижению реактивности тканей. Обратный эффект оказывают воздействия на периферическую иннервацию. Нарушения ее повышают чувствительность, повреждаемость тканей по отношению к механическим, физическим и химическим факторам. Усиление же эфферентных нервных импульсов, идущих к тканям, ослабляет чувствительность последних к повреждающим воздействиям.
Касаясь механизма тканевых реакций на внешние воздействия, следует отметить, что ткани, являясь строительными материалами органов, представляют собой в то же время живые системы и, как таковые, сами по себе способны воспринимать внешние раздражения и реагировать на них. Однако в целом организме и процесс восприятия тканями раздражений извне, и тканевые ответы на них находятся под контролем прежде всего нервной системы.
Чувствительность к внешним воздействиям нервных рецепторов - специализированных воспринимающих аппаратов нервной системы, конечно, несравненно выше, чем любых других тканевых элементов. Восприятие внешнего воздействия рецептором передается в центральную нервную систему гораздо быстрее, чем это воздействие будет воспринято самой тканью. В ответ на сигнал с периферии из центральной нервной системы по эфферентным (центробежным) путям идут в соответствующие ткани импульсы, которые регулируют уровень чувствительности данных тканей к этому внешнему воздействию, и, таким образом, определяют степень их реагирования. Импульсы тормозного характера сдерживают чрезмерное и анархическое развертывание тканевых реакций. Следовательно, несмотря на способность тканей как живых частей организма к осуществлению чисто местных реакций, последние в условиях нормальной иннервации не реализуются.
На жизнедеятельность организма оказывают влияние самые разнообразные факторы внешней среды и состояние его собственной внутренней среды. Возьмем, к примеру, суточный ритм жизни организма. В ходе исторического развития человек и животные приспособились именно к земным условиям, определяемым Землей как планетой с ее суточным вращением вокруг своей оси. В связи с этим у всех живых существ изменяются в течение суток их коренные свойства ассимиляции и диссимиляции. Периодическая смена различных процессов в организме определяется в течение суток чередованием периодов света и темноты, а отсюда бодрствования, активности или же покоя, сна. Причем у одних видов животных, что обусловлено их филогенезом, период бодрствования приходится на день, у других - на ночь. Как выяснилось, это находит свое отражение и в жизнедеятельности тканей.
Очень удобным показателем сдвигов, испытываемых в течение суток тканью, является ее митотическая активность. Было установлено, что у человека и животных с дневным образом жизни максимум митозов приходится на ночные часы, а у животных, бодрствующих ночью, митозов, наоборот, больше в дневные часы. Состояние возбуждения нервной системы организма приводит к торможению митотической активности в его органах и тканях. Поскольку при этом кривые суточного ритма митозов в разных органах оказываются не тождественными, а значительно отличаются друг от друга, нужно придти к заключению о наличии не только организменной, но и органной регуляции митотической активности тканей.
Если изменить достаточно резко условия существования организма, то это скажется и на состоянии его тканей, что может быть установлено во многих случаях тем же способом - определением их митотической активности. Очень демонстративными в этом отношении оказались данные Ф. Х. Шарипова, поднявшего крыс из долины в высокогорье (на Анзобский перевал Памира, где высота над уровнем моря 3600 м). В новых условиях, прежде всего недостаточности кислорода во внешней среде, сразу же произошло изменение митотической активности почечного эпителия - в первые два дня после подъема животных в горы она снизилась втрое. На 4 - 5-й день пребывания крыс в условиях высокогорья, когда они несколько приспособились к новой внешней среде, митотическая активность эпителиальной ткани почечных канальцев удвоилась. Однако даже спустя один месяц, то есть в состоянии относительной адаптации животных к высокогорным условиям, митотическая активность ткани была статистически достоверно ниже, чем в долине. Через 4 - 5 дней, после того как крысы были доставлены с гор в долину, митотическая активность эпителия почечных канальцев у них фактически возвратилась к исходной.
Л. Я. Бляхер и С. Л. Бляхер с помощью определения митотической активности эпителия тонкого кишечника у мышей установили, что состояние тканей зависит и от условно-рефлекторной деятельности организма. Вспомним знаменитые опыты академика И. П. Павлова по выработке условных рефлексов. Если собаке ввести в пасть каплю кислоты или кусочек мяса, у нее сразу же потечет слюна. Этот ответ на непосредственное раздражение рецепторов И. П. Павлов обозначил как врожденный, безусловный рефлекс. Но можно наблюдать процесс слюноотделения у собаки на любой сигнал из внешней или внутренней среды, если сочетать его с дачей животному пищи и повторить это какое-то количество раз. Например, зажечь перед глазами собаки электрическую лампочку и тут же, потушив ее, подкормить животное кусочком мяса. После нескольких таких экспериментов достаточно будет только электрического сигнала, чтобы у собаки потекла слюна. Подобного рода приобретенные рефлексы, по И. П. Павлову, относятся к условным. Л. Я. и С. Л. Бляхеры установили, что при непосредственном кормлении у мышей в эпителиальной выстилке тонкого кишечника митотическая активность резко усиливается. Затем они проделали следующий опыт: посадили мышей в стеклянный сосуд, закрытый сеткой таким образом, что животные могли обонять и видеть лакомый кусочек пищи, положенный на верх сетки, но для них он оставался недоступным. При этом у мышей было отмечено заметное учащение митозов в эпителиальной выстилке кишечника. Это было условно-рефлекторным ответом на раздражитель.
Таким образом, ткани, входя как структурные компоненты органов в систему целого организма, находятся под его постоянным регулирующим влиянием.
Мы рассмотрели различные вопросы, касающиеся строения, жизнедеятельности, а также развития в фило- и онтогенезе клеток и тканей - материалов, из которых построено тело животных и человека. Структура и функция любых составных элементов тела животного или человеческого организма, в том числе его клеток, тканей и органов, взаимно обусловливают друг друга и каждой структуре строго соответствует ее функциональный эквивалент. Вот почему, даже изучая клетки и ткани чисто морфологическими статистическими методами, мы не только ставим перед собой задачу дать функциональную трактовку обнаруживаемым структурам, но и вправе экстраполировать полученные результаты на живой организм.
Тело человека и родственных ему организмов - позвоночных животных представляет собой чрезвычайно расчлененную и в то же время целостную систему. Части этой сложной системы являются, в свою очередь, системами все более низкого порядка, такими как разнообразные органы тела, выполняющие в нем различные функции, ткани, строящие данные органы, и клетки, образующие эти ткани. Каждая из этих систем разных уровней организации обладает способностью к саморегуляции, обеспечивающей единство протекающих в ней процессов развития и деятельности. И в то же время работа такой системы подчинена регулирующему воздействию системы высшего порядка, а в конечном счете координируется со стороны организма как целого.
В процессе эволюции возникли клетки-организмы, потомки которых в изобилии населяют нашу Землю. Их становлению в филогенезе предшествовали события, длительность которых измерялась миллиардами лет. Жизнь на земле возникла около 3 миллиардов лет тому назад и качественной особенностью живых существ является с того времени их белково-нуклеотидная природа, обуславливающая непрерывно текущий обмен веществ с окружающей внешней средой. Именно ими определяется специфический характер всех других черт, свойственных живым организмам: движение, чувствительность, развитие, рост и размножение. Субстратом - почвой жизненных процессов являются белки, а специальный аппарат их воспроизведения, как показала современная молекулярная биология, представлен нуклеиновыми кислотами.
За миллиарды лет существования жизни на Земле взаимоотношения живых существ с внешней средой становились все более сложными и разнообразными, а вместе с этим становились все более сложными и совершенными сами живые организмы. В результате длительного исторического процесса из первоначально бесформенных мельчайших комочков живого вещества развились, под действием открытого Ч. Дарвиным естественного отбора, одноклеточные организмы. Некоторые из них объединились во все более и более усложнявшиеся колонии, и в результате дифференцировки первоначально однородных клеточных элементов колоний, разделения труда между ними, появились многоклеточные организмы с телом, расчлененным на органы и ткани. Вершиной эволюции явилось возникновение позвоночных животных, а в ряду беспозвоночных -насекомых и головоногих моллюсков.
Первыми позвоночными животными были очень примитивные бесчелюстные рыбы, одетые панцирем из костных пластинок. Появились панцирные рыбы около полумиллиарда лет тому назад. И прошло еще более 100 миллионов лет, пока позвоночные стали осваивать сушу и создались первые земноводные - стегоцефалы. Первоптицы и примитивные млекопитающие возникли на нашей планете лишь немногим более полутораста миллионов лет, а человек (первоначально в виде обезьяно-человека) всего около 3 миллионов лет.
Но позвоночные это только подтип одного из многих типов животных, обитающих на нашей Земле, а именно: губок, кишечнополостных, плоских, круглых и кольчатых червей, членистоногих, моллюсков, иглокожих и хордовых. Число видов животных и анатомическое разнообразие их необозримо. И при всем том поражает сходство материалов, из которых построены их тела - тканей. Как подчеркивал академик А. А. Заварзин, основоположник советской эволюционной гистологии, структурное сходство однозначных тканей у неродственных животных и ограниченность их типов объясняются параллелизмом выполняемых ими в любом многоклеточном организме элементарных биологических функций (пограничности, внутреннего обмена, сократимости, раздражимости).
В каждой же клетке любого многоклеточного организма можно обнаружить общие черты, присущие ей как представителю определенного уровня организации живой материи. А, с другой стороны, каждая клетка несет в своей структуре и функции признаки ее принадлежности к определенной ткани. На это накладываются и видовые особенности, свойственные данному организму.
При использовании материалов сайта активная ссылка обязательна:
http://anfiz.ru/ 'Анатомия и физиология человека'