Кора мозга
Архипалий и неопалий (старый плащ и новый плащ). Мозговая кора, взятая в своей совокупности, обычно носит название pallium - плаща. Та часть pallii, которая связана с обонятельным аппаратом, развилась в эволюции позвоночных гораздо раньше, чем необонятельная часть pallii, и поэтому носит название - archipallium (старый плащ). Вся же необонятельная область мозговой коры носит название neopallium (новый плащ).
Archipallium достигает максимального развития у низших млекопитающих, особенно у сумчатых (кенгуру, опоссум), и состоит из hippocampus и lobus pyriformis (грушевидная доля). У человека на поверхности мозговой коры архипалий занимает очень незначительную область. Большая часть его скрыта в глубине вследствие образования вдавления (складки) вдоль заднего края неопалия. Самый передний отдел извилины морского коня и слабо выраженная stria lateralis обонятельной ножки - вот все, что осталось в человеческом мозгу от сравнительно обширной lobus pyriformis многих низших млекопитающих. Сама извилина морского коня (gyrus hippocampi), которая лежит на наружной стороне lissurae dentitae, хотя и продолжается непосредственно в грушевидную область, не относится к обонятельному мозгу (rhinencephalon), - она принадлежит к неопалию.
Rhinencephalon - наиболее архаическая часть мозговых полушарий - в мозгу низших позвоночных (рыбы, амфибии, рептилии) составляет наибольшую часть их полушарий. В мозгу человека rhinencephalon представлен: bulbus olfactorius, tractus olfactorius и его корешки, переднее продырявленное пространство, uncus, gyrus subcallosus, septum lucidum, hippocampus, fornix, gyrus dentatus и gyrus supracallosus.
Слабая бороздка, которая отделяет uncus от полюса височной доли incisura temporalis (височная вырезка), является одной из наиболее древних борозд. Она представляет собой пограничную борозду между архипалием и неопалием (sulcus ecto-rhinalis).
За исключением архипалия (обонятельного мозга - rhinencephalon) и полосатого тела (corpus striatum), вся остальная часть полушарий представляет собой неопалий. Он охватывает не только область извилин, но и связанное с ними белое вещество. Наибольшего развития неопалий достигает в мозгу человека. В своей филогенетической эволюции архипалий и неопалий развиваются более или менее независимо друг от друга. В одних случаях архипалий атрофируется, а неопалий достигает высокой степени развития (человек, обезьяны), в других случаях происходит обратная картина: архипалий образует значительную часть полушарий, а неопалий сравнительно малую часть (рис. 239, 240, 241, 242, 243).
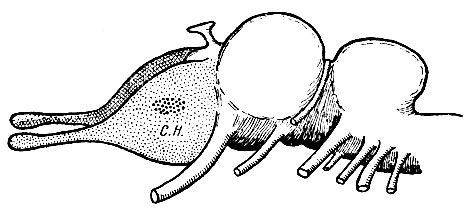
Рис. 239. Схема примитивного мозга позвоночного животного. Область коры, связанная с не обонятельными впечатлениями (обозначена более грубыми точками); область коры, связанная с обонятельными впечатлениями (обозначена мелкими точками)
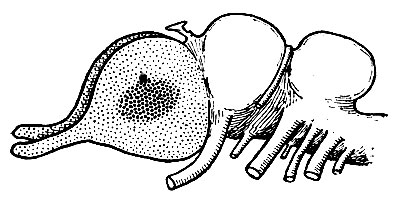
Рис. 240. Схема мозга в дальнейшей стадии эволюции. Необонятельная кора (более грубые точки) занимает на поверхности полушария более обширную область, чем в предыдущей стадии
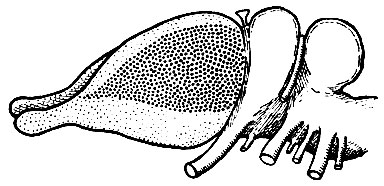
Рис. 241. Схема примитивного мозга млекопитающего, показывающая дальнейшее развитие неопалия (область, заштрихованная крупными точками)
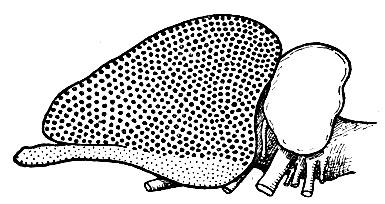
Рис. 242. Схема типичного мозга млекопитающего, показывающая, что неопалий (область, заштрихованная крупными точками) распространился за счет архипалия (область, заштрихованная мелкими точками) и занимает теперь большую часть полушария
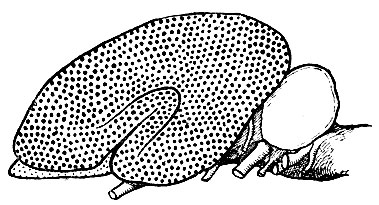
Рис. 243. Схема законченного мозга млекопитающего. Неопалий увеличился; на нем стали появляться борозды. Архипалий (область, заштрихованная мелкими точками, занимает лишь незначительный, краевой отдел полушария)
Надо думать, говорит Wood Jones, что мозг первичного млекопитающего состоял из ряда нервных узлов, причем каждый узел служил или для восприятия определенного чувствования, или для выполнения определенного действия. В таком мозгу впечатления от определенных органов чувств шли по определенным путям к своим определенным анатомический станциям, а эти узловые станции находились во взаимной связи друг с другом. В придачу к вышеописанному аппарату здесь существовал вырост с каждой стороны переднего мозга - небольшое мозговое полушарие, - предшественник обширной мозговой коры высших типов. В этом первичном мозговом полушарии впечатления, пришедшие по различным путям, соединялись между собой и накоплялись. Здесь возник процесс сознания, суждения и памяти. Связи отдельные стволовых) центров с корой полушарий устанавливались постепенно и в определенном порядке. Первым органом, установившим связь с корой, был орган обоняния. У предшественников млекопитающих мозговые полушария по функции были по преимуществу обонятельными, и даже когда возникло истинное млекопитающее и все другие чувствования получили должное представительство в мозговой коре (неопалии), поведение животного все еще находилось в гораздо большей степени под влиянием обонятельных впечатлений, чем каких-либо других чувств. Это зависело, говорит Wood Jones, не только от того факта, что чувство обоняния уже имело в своем ведении выработанный аппарат в мозговых полушариях задолго до вступления в полушария соответствующего представительства других органов чувств, но также и от того, что для маленьких, вечно тычащих в землю своим носом животных, какими были первичные млекопитающие, руководство обонятельными впечатлениями как для отыскивания пищи, так и для распознавания друзей и врагов, оказывало гораздо больше услуг, чем все другие чувствования.
Таким образом, духовная жизнь первобытного существа протекала по преимуществу в атмосфере запахов, и о каждом объекте внешнего мира составлялось суждение прежде всего и главным образом на основании его запаха. Чувства прикосновения, зрения, слуха являлись лишь вспомогательными при господствующем влиянии обоняния.
Раз такое животное покидало твердую почву и переходило к древесному образу жизни, все вышеприведенное должно было измениться, ибо вдали от земли руководство чувством обоняния imoro теряет в своей полезности. Обонятельные части мозга и самый периферический чувствующий обонятельный аппарат у живущих на деревьях животных заметно атрофированы; обонятельные впечатления играют лишь малую роль в жизни приматов. У человека обонятельные впечатления играют еще меньшую роль.
Wood Jones в своей работе "Arboreal man" (древесный человек) очень картинно описывает последовательное возникновение у млекопитающих чувствительно-двигательных центров коры, степени их последующего развития и причин их топографо-анатомических взаимоотношений. Из работы Wood Jones я и заимствую нижеприведенные соображения.
Примитивное млекопитающее прежде всего познавало мир, знакомясь с запахом предметов, затем осязанием, - прикасаясь к предметам концом своего рыла, тыча всюду свое рыло. Впечатления, получаемые от органа обоняния, дополняются, таким образом, впечатлениями, получаемыми при помощи органа осязания. Эти осязательные впечатления, получаемые рылом, скоро находят путь к коре мозга. Длинное вытянутое рыло является типичным для всех первичных млекопитающих: однопроходных, сумчатых и даже плацентарных. С освобождением передней конечности, с развитием захватывающей способности ручной кисти замечается постепенное отступление удлиненного рыла кзади. Захватывающая кисть заступает место захватывающих челюстей; мало того, она становится органом осязательным, органом испытующим; она не только постепенно берет на себя значительную долю работы челюстей, но и осязательную функцию рыла. Главный осязательный орган животного переносится таким образом с одно и части тела на другую. Свободная чувствующая рука идет теперь впереди животного на его жизненном пути вместо рыла. В своей древесной жизни животное раньше исследует предмет осязанием своих рук, а уже затем, если пожелает, может обследовать его при помощи обоняния. Свободная рука открывает новые пути для обследования мира в придачу к старым путям - обонятельному и осязательному при помощи рыла. Увеличивается число объектов, доступных исследованию, увеличивается полнота исследования, - рукой мы можем определить очертание предмета, строение его поверхности, объем, температуру, твердость, мягкости; вес. Сорванный и поднесенный к глазам предмет повышает нашу зрительную оценку. Осязательное чувство руки помогает нам проверять, и связывать впечатления, получаемые при помощи других органов чувств.
Как было указано выше, первые ощущения, достигшие коры, были обонятельные; вслед за ними достигли коры осязательные ощущения, идущие от рыла, и поместились в коре в непосредственном соседстве с обонятельными. Весьма вероятно, что и вкусовые ощущения из полости рта также поместились по соседству с обонятельными. Так как в стволовой части мозга двигательные центры расположены по соседству с соответствующими чувствующими, то весьма вероятно, что двигательная область коры ассоциированная с движениями рыла, будет развиваться по соседству с соответствующими чувствующими областями. С появлением руки как чувствующего органа к вышеописанным чувствительно-двигательным областям коры должна была присоединиться новая область - чувствительно-двигательная область руки, расположенная по соседству с чувствительно-двигательной областью рыла. Для древесного животного с уменьшением роли чувства обоняния слуховые и световые ощущения приобретают большое значение (рис. 244-248).
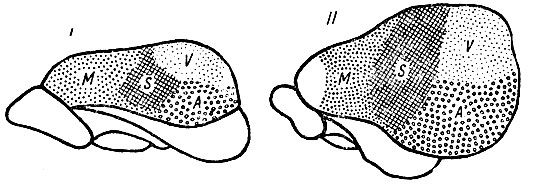
Рис. 244. Слева мозговое полушарие прыгунчика (macroscelides), справа - мозговое полушарие тупаий (tupaia). И то, и другое животное принадлежат к насекомоядным (insectivora). Корковые области показаны, как они были определены проф. Elliot Smith. M - двигательная область; S - общая чувствительность; V - зрительная область; A - слуховая область. Белое поле, расположенное на обоих рисунках в нижнем отделе полушария, обозначает обонятельную область. Белое поле на рисунке II, расположенное впереди двигательной коры, обозначает префронтальную область
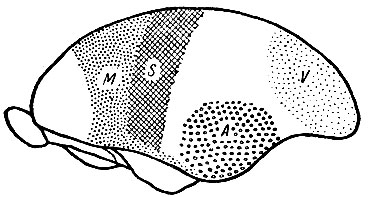
Рис. 245. Мозговое полушарие долгопята (Tarsius). Корковые области показаны, как они были определены проф. Elliot Smith. Сравнивая этот рисунок с предыдущим, мы отмечаем расширение префронтальной области и развитие ассоциационных областей между полями - зрительным, слуховым и полем общей чувствительности. Обозначения те же, что и на предыдущем рисунке
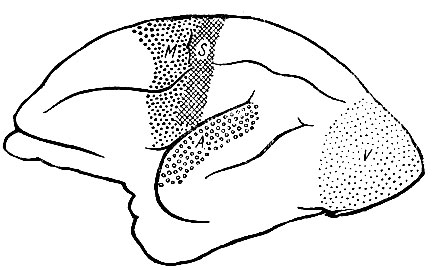
Рис. 246. Мозговое полушарие лемура. Корковые области показаны, как они были определены Brodmann. Имеется значительное расширение ассоциационных областей по сравнению с тем, что мы отмечали у долгопята
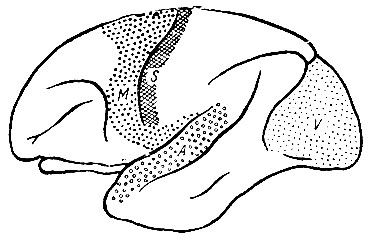
Рис. 247. Мозговое полушарие обезьяны макака (macacus). Корковые области показаны, как они были определены Brodmann. Дальнейшее развитие стадии, представленной лемуром. Особенно заметно увеличение префронтальной области
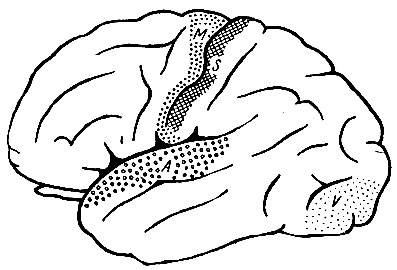
Рис. 248. Корковые области мозгового полушария человека. Имеется усиленное развитие ассоциационных областей. (Wood Jones. Сделаны небольшие изменения)
Слуховая область впервые появляется в задне-нижнем отделе мозговых полушарий; зрительная область лежит непосредственно над нею, а осязательно-двигательная кора - кпереди от обоих. Когда мозговые полушария в связи с увеличением области представлений увеличиваются, расширение вышепоименованных областей не идет равномерно. Часть коры, прилежащая сбоку к серым узлам corporis striati, не участвует в этом разрастании и остается более или менее фиксированной (insula Reilii), тогда как участки, расположенные вокруг нее, разрастаются. При этом росте слуховая область коры, отталкиваемая сверху расширяющейся зрительной областью, будет распространяться по направлению книзу и кпереди. В конце концов, она становится отделенной от зрительной области так называемой промежуточной областью коры. Разрастающийся промежуточный корковый слой отталкивает к задней границе полушарий зрительную область и отделяет ее целым участком от осязательно-двигательной области. На переднем конце полушарий развивается новая префронтальная область. Во вновь образованных при разрастании коры промежуточных областях поместились области ассоциаций: здесь сливаются и координируются впечатления, находящиеся в пограничных с ними чувствующих областях. Эти ассоциационные участки между слуховым и зрительным полями, между зрительным и осязательно-двигательным, расширившись у древесных приматов, становятся отличительной чертой человеческого мозга.
Животное без корковой чувствительно-двигательной области совершает все его действия при отсутствии какого-либо картинного сознания об этих действиях. Животное с чувствительно-двигательной корковой областью, наоборот, совершает действия, о которых оно имеет определенное картинное представлении. Оно знает, что оно делает; помнит, что оно делало и может представить, что оно будет делать.
Brodmann, Economo и др. делят толщу коры неопалия у человека на шесть слоев (пластин) (рис. 249): 1) зональная пластина, состоящая из тангенциальных волокон; 2) пластина наружных зерен или малых пирамидальных клеток; 3) пирамидальная пластина или слой средних и больших пирамидальных клеток; 4) пластина внутренних зерен, содержащая покрытые миэлином волокна наружной линии Baillarger; 5) ганглионарная пластина, содержащая в двигательной области гигантские пирамидальные клетки Betz, а в большинстве других областей - покрытые миэлином волокна внутренней линии Baillarger; 6) мультиформная пластина или слой полиморфных клеток. Bolton соединил 2-й и 3-й слои Brodmami в один слой и получил таким образом у человека пять слоев коры неопалия: 1) наружная пластина волокон (нервные фибриллы); 2) наружная клеточная пластина (пирамидальные клетки); 3) средняя клеточная пластина (зерновидные клетки); 4) внутренняя пластина волокон (нервные аксоны); 5) внутренняя клеточная пластина (полиморфные клетки). Watson объединяет вместе все слои, лежащие над слоем зерен, и называет эти объединенные слои супрагранулярной (надзерновидной) корой; слой зерен называет гранулярной (зерновидной) корой, а слои, расположенные ниже слоя зерен, называет инфрагранулярной (подзерновидной) корой.
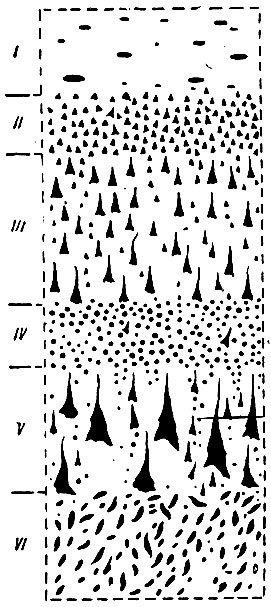
Рис. 249. (Объяснение в тексте)
И онто- и филогенетически различные слои мозговой неопалиальной коры появляются в следующем порядке: первой в процессе развития появляется инфрагранулярная кора; очень быстро она достигает своей зрелости: представляет очень малые различия в своей толщине у высших и низших млекопитающих. Гранулярный, слой появляется вскоре после инфрагранулярного. Последним появляется супрагранулярный слой. Своей полной зрелости он достигает позднее других слоев. Он слабо выражен в неопалиальной коре низших млекопитающих; его толщина значительно увеличивается у высших млекопитающих и достигает своего наибольшего развития у человека.
Таким образом, неопалиальную кору низших млекопитающих, где над зерновидный слой еще недостаточно развит, можно рассматривать в общем как бы состоящей из трех слоев: 1) из слоя зерен, 2) из расположенного под ним слоя покрытых миэлином рецепторных волокон и 3) еще глубже лежащего слоя полиморфных клеток. В такой коре рецепторные импульсы, приносимые рецепторными аксонами, будут накопляться в зерновидных клетках. Позднее эти клетки под влиянием стимуляции могут освобождать накопившуюся нервную энергию и давать таким образом отсроченный эффекторный ответ вместо немедленного. В мозгах такого рода (например, у кенгуру) приносимые импульсы, а по-видимому, и зерновидные клетки, связаны со всеми частями коры того же полушария при помощи ассоциационных нейронов полиморфного слоя. У более высокостоящих млекопитающих к перечисленным слоям присоединяется еще более расположенный кнаружи слой пирамидальных клеток и корковые нейронные пути таким образом значительно увеличиваются. Элементы контроля над действиями и память предыдущих стимулов и действий начинают заметно вступать в поведение животного. В конце концов, с присоединением ассоциационных областей и огромным размножением в этих областях нервных элементов как инфрагранулярного, так и супрагранулярного типов мы достигаем мозга человека с его проявлениями - памяти, разума, речи. Здесь мы имеем дело с теми же структурными элементами, что и у других млекопитающих, но с сильно развившейся системой промежуточных нейронов.
Функции инфрагранулярной мозговой коры, состоящей из внутренней пластины волокон и внутренней клеточной пластины (полиморфные клетки), как показали исследования Bolton на мозгах человека и исследования Watson на мозгах млекопитающих, сводятся к обслуживанию низшей волевой и инстинктивной деятельностей животного. Главной функцией инфрагранулярной коры человека, говорит проф. Berry, является управление теми действиями нашего тела, которые образуют базу многочисленных сложных рефлекс сов, необходимых для сохранения индивидуума и вида, как, например, добывание пищи, отыскивание соответствующего убежища, отыскивание и обладание субъектом другого пола и т. п. Ширина (глубина) инфрагранулярного слоя коры у взрослых нормальных людей более или менее постоянна. По Bolton, при тяжелых формах аменции и деменции глубина этого слоя заметно уменьшается. Словом "аменция" Bolton обозначает умственное состояние больных, страдающих от недостаточного развития корковых нейронов (субэволюция мозга), а словом "деменция" - умственное состояние больных, страдающих от перерождения корковых нейронов.
При тяжелых формах аменции и деменции больные не в состоянии сами надлежащим образом обслуживать свои естественные нужды.
Гранулярный слой, который всегда хорошо развит на воспринимающих участках коры, в первую очередь обслуживает восприятие рецепторных импульсов, независимо от того, проходят ли эти рецепторные импульсы прямо от низших рецепторных нейронов или не прямо - от других областей коры. Как известно, первоначальным связующим типом нервной ткани являются ассоциационные и коммиссуральные клетки, наблюдаемые в спинном мозгу почти всех позвоночных животных, и лишь позднее в надсегментальных прибавлениях мозга появляются зерновидные клетки. Ясно, говорит проф. Berry, что они служат для восприятия и накопления рецепторных импульсов и для ассоциирования впечатлений данного момента с ранее накопленными впечатлениями, а следовательно, и в коре человеческого мозга зерновидные нейроны должны играть роль в процессе запоминания.
Слой надгранулярных пирамид является новейшим добавлением к неопалиальной мозговой коре млекопитающих. По Watson, у насекомоядных надгранулярный слой пирамидальных клеток находится в зачаточном (рудиментарном) состоянии, но становится все более и более выраженным по мере восхождения по лестнице млекопитающих и лучше всего выражен у человека. Слой надгранулярных пирамид является единственным, глубина которого заметно вариирует на нормальных мозгах. Так как этот слой развился последним, то он раньше других слоев поддается регрессивным изменениям.
По Berry, супрагранулярный слой в общем можно рассматривать как удвоение инфрагранулярного слоя, однако и форма клеток, и функция супрагранулярного слоя иная, чем инфрагранулярного. Многочисленные пирамидальные клетки супрагранулярного слоя перемешаны с клетками 2-го типа Golgi, с горизонтальными клетками Cajal и с клетками Martinotti. Рецепторные аксоны, приходящие главным образом от зрительного бугра (а также и от других частей мозга) и образующие наружную линию Baillarger, обслуживают по преимуществу супрагранулярную кору.
Хотя все чисто двигательные клетки мозговой коры имеют пирамидальную форму, все же, говорит Berry, нельзя рассматривать все пирамидальные клетки коры как двигательные. Исходя из того положения, что чем длиннее аксон, тем обширнее клетка, Berry высказывает предположение, что малые пирамидальные клетки образуют короткие ассоциационные связи с прилежащими извилинами; пирамидальные клетки среднего размера могут давать ассоциационные связи с более отдаленными извилинами, и, наконец, большие пирамидальные клетки, за исключением клеток Betz, могут быть рассматриваемы как клеточные тела по крайней мере некоторых коммиссуральных волокон, идущих через мозолистое тело к тем или другим участкам коры полушарий противоположной стороны. Супрагранулярный слой коры обслуживает "психические", ассоциативные функции нашего мозга в противоположность органическим, инстинктивным функциям инфрагранулярного слоя, на которые супрагранулярный слой может действовать тормозящим образом. Функции супрагранулярного слоя связаны с личным опытом животного и подлежат значительным изменениям на протяжении жизни индивидуума.
Неопалиальная кора млекопитающих не является однородной по своей структуре на всем своем протяжении, но может быть подразделена на ряд участков, которые отличаются друг от друга своим микроскопическим строением. Одни из этих корковых участков являются чувствующими проекционными областями коры, куда приходят чувствующие импульсы от нижележащих уровней мозга. Так 1 -3-я области Brodmann связаны с общей чувствительностью, 22-я область - со слуховой чувствительностью, 17-я область со - зрительной чувствительностью. Зрительная область сравнительно обширна и хорошо дифференцирована у всех приматов, по-видимому, в связи с преобладанием у. них чувства зрения. На вентральной стороне лобного полюса, как это хорошо видно, например, на рисунке мозга лемура microcebus, имеется небольшая область коры (8-е поле), которую можно назвать префронтальной областью и которая получает волокна от дорзо-медиального ядра зрительного бугра. Это ядро связано с висцеральными центрами hypothalamus. У человека префронтальная кора занимает 8-12-е и 44-47-е поля Brodmann, причем 8-е поле является переходным. По-видимому, префронтальная кора, говорит Le Gros Clark, развивается как механизм, при помощи которого висцеральные реакции, которые являются единственным элементом эмоционального поведения и инстинктивных импульсов, могут быть контролируемы высшими функциональными уровнями мозга. Интересно отметить, что префронтальная кора, а вместе с ней и дорзо-медиальное ядро зрительного бугра подвергаются прогрессивному расширению в ряду приматов, достигая своего наивысшего выражения у человека. У низших млекопитающих префронтальная область выражена слабо. По наблюдениям Le Gros Clark, эта область развивается как распространение кпереди коры островка (поля 13-16-е) и не может быть рассматриваема как дифференцированный участок собственно лобной коры. Поле 4-е представляет двигательную проекционную кору, которая дает начало двигательным пирамидальным трактам, при помощи которых непосредственно вызываются и контролируются произвольные движения. Ассоциационными областями коры называют те области, которые не являются непосредственно воспринимающими импульсы, идущие от более низких мозговых уровней. Ассоциационные области связаны с корреляцией функций проекционных областей. Они представляют собой анатомический субстрат таких мозговых процессов, как ассоциация идей, память и т. п. Эти ассоциационные области лучше выражены на мозгах более высокостоящих приматов; по мере своего распространения они все более и более отдаляют друг от друга проекционные области. У лемура microcebus ассоциационные поля по сравнению с ассоциационными полями низших млекопитающих одинакового с microcebus веса довольно обширны, особенно теменное ассоциационное поле, отделяющее поле общей чувствительности от зрительного поля.
Структура мозговой коры. Мозговая кора состоит из большого количества клеточных элементов и большого количества нервных волокон как безмиэлиновых, так и покрытых миэлином. Нервные клетки принадлежат к различным морфологическим типам и собраны в более или менее определенные слои.
Большинство нервных волокон коры имеет радиальное направление, однако некоторые зоны характеризуются многочисленными волокнами, которые идут параллельно поверхности.
Большинство нейронов коры соответственно их форме делят на три группы: 1) пирамидальные клетки, 2) полиморфные клетки и 3) зерновидные клетки. Помимо указанных трех типов нервных клеток, которые наблюдаются почти во всех отделах коры, на некоторых участках коры встречаются также нейроны и других типов, например, клетки Cajal, клетки Martinotti и т. д.
Нижеприводимая более или менее детальная характеристика клеток и волокон коры взята мною у Kuntz (рис. 250).
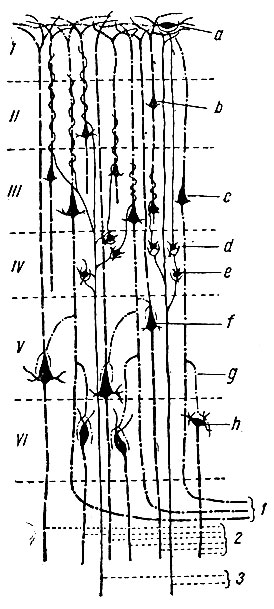
Рис. 250. Схема некоторых главных внутрикорковых синаптических связей. a - горизонтальная клетка; b - малая пирамидальная клетка; c - пирамидальная клетка среднего объема; d - клетка Martinotti; e - зерновидная клетка; f - большая пирамидальная клетка; g - коллатераль; h - полиморфная клетка. 1 - ассоциационные волокна; 2 - выносящие волокна; 3 - приносящие волокна (по Kuntz)
Пирамидальные клетки образуют большинство корковых нейронов и встречаются почти во всех отделах коры. Они сильно вариируют по своему объему и обычно классифицируются как малые, средние, большие и гигантские пирамидальные клетки. Верхушка пирамидальной клетки обращена к поверхности коры; от верхушки отходит верхушечный дендрит, который направляется к поверхности и оканчивается в виде разветвлений в поверхностном слое коры. Его длина вариирует в зависимости от расстояния клеточного тела от поверхности. Обычно он дает небольшое число ветвей. Различное число небольших дендритов отходит также от основания и от боковых сторон пирамидальной клетки. Аксон отходит от основания клетки и направляется в подкорковое белое вещество. Аксон дает коллатерали, которые разветвляются в прилежащей коре.
Полиморфные клетки также имеются почти во всех областях коры, но встречаются лишь в самом глубоком слое. Они вариируют по форме и по величине в различных областях коры, но лишь в сравнительно ограниченных пределах. Длинная ось их клеточного тела по преимуществу направлена перпендикулярно к поверхности коры. Дендрит возникает из наружного полюса. Нейрит возникает или от внутреннего полюса, или со стороны клеточного тела. Помимо двух вышеуказанных отростков, клеточное тело может дать начало небольшому числу небольших дендритов (от 3 до 6). Аксон направляется в подкорковое белое вещество.
Зерновидные клетки чрезвычайно многочисленны и присутствуют почти во всех отделах коры. Они по преимуществу небольшого объема и сильно вариируют по форме. Они дают начало различному числу коротких дендритов и короткому аксону, который отдает коллатерали и оканчивается в прилежащей коре. Они встречаются почти во всех слоях коры, но обильны по преимуществу во 2-м и в 4-м слоях.
Горизонтальные клетки Cajal в небольшом числе имеются во всех областях коры, но располагаются лишь в поверхностном слое. Эти клетки небольшого размера, имеют веретенообразную, звездчатую или неправильную форму, отдают длинные ветвящиеся дендриты, которые распространяются горизонтально в поверхностном слое. Аксоны клеток Cajal или коротки и заканчиваются вблизи их клеточного тела, или сравнительно длинны, покрыты миэлином и идут параллельно поверхности. Они образуют значительную часть тангенциальных волокон поверхностного слоя.
Клетки Martinotti встречаются по преимуществу в полиморфном слое; в небольшом количестве встречаются и в других слоях коры. Их аксоны направляются к поверхности, разветвляются в поверхностном слое и образуют часть тангенциальных волокон этого слоя.
Волокна мозговой коры. В состав волокон мозговой коры входят: 1) дендриты и аксоны корковых нейронов и 2) приносящие волокна, вступающие в кору и устанавливающие связи с ее нейронами. Эти последние включают в свой состав как миэлиновые, так и безмиэлиновые волокна. Многие из волокон внутри коры идут радиально, другие идут тангенциально, пересекая радиальные волокна под прямыми углами. Радиальные волокна коры включают в свой состав: 1) аксоны пирамидальных и полиморфных клеток, которые направляются в белое вещество мозга, 2) концевые отделы восходящих проекционных волокон, 3) коммиссуральные и 4)ассоциационные волокна, которые вступают в кору, чтобы закончиться в ней, и 5) аксоны клеток Martinotti. Радиальные волокна расположены пучками, которые направляются радиально от подкоркового белого вещества через последовательные корковые слои к верхнему корковому слою. На вертикальных разрезах коры радиально идущие волокна чередуются с радиально расположенными колоннами нервных клеток.
Тангенциальные волокна коры состоят главным образом из: 1) коллатералей и концевых ветвей аксонов пирамидальных клеток, полиморфных клеток и клеток Martinotti, из ветвлений аксонов зерновидных клеток и горизонтальных клеток Cajal, из коллатералей и концевых ветвей проекционных, коммиссуральных и ассоциационных волокон. Все эти волокна образуют несколько слоев, из которых важнейшими являются: 1) поверхностное тангенциальное сплетение, расположенное в поверхностном отделе наружного слоя; 2) полоска Бехтерева, расположенная в наружном отделе второго слоя; 3) наружная полоска Baillarger, расположенная в четвертом слое (внутренний слой зерен); 4) внутренняя полоска Baillarger, рас положенная в глубоком отделе пятого слоя, или между пятым и шестым слоями и б) глубокие тангенциальные волокна, расположенные в более глубоких отделах шестого слоя. Описанные нами слои тангенциальных волокон вариируют в сравнительно широких пределах в различных областях коры, причем в некоторых областях один или более слоев отсутствуют.
Архитектонические коpковые области. Почти все области коры, за исключением обонятельной, содержат в себе шесть вышеуказанных клеточных слоев, но и по толщине, и по структуре каждый из этих клеточных слоев отличается от соответствующего слоя не только в различных корковых областях, но и в различных отделах, принадлежащих одной и той же структурной корковой области.
1-й и 2-й слои клеток коры обычно бывают тоньше на вершине извилины и толще на дне соответствующей борозды, чем на стенках борозды. Глубокие слои (5-й и 6-й), наоборот, бывают толще на верхушке извилины и тоньше на дне борозды, чем на стенках борозды. Средние клеточные слои (3-й и 4-й) по характеру своих изменений подходят к нижним слоям, хотя эти изменения выражены в них менее резко (рис. 251).
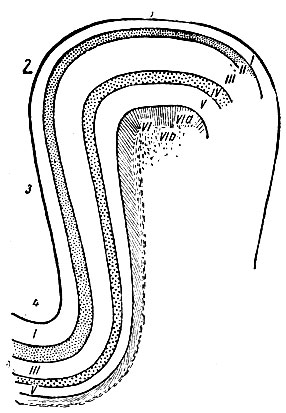
Рис. 251. Схема, представляющая изменения толщины корковых слоев на различных уровнях поперечного сечения мозговой извилины. 1 - вершина извилины; 2 - угол извилины; 3 - стенка извилины; 4 - дно борозды. Римские цифры обозначают соответствующие слои коры (по Economo и Koskinas)
Эта разница в толщине глубоких и поверхностных слоев коры в глубине борозд и на выпуклости извилин связана, по-видимому, с процессом образования складчатости коры. В общем можно сказать, что там, где кривизна слоя уменьшается, клетки его рассеиваются и толщина слоя становится меньше, там же, где кривизна слоя увеличивается, клетки его концентрируются и толщина становится больше. На выпуклых частях извилин кривизна слоев будет возрастать с поверхности в глубину, а на дне борозд уменьшаться с поверхности в глубину.
С указанными изменениями толщины слоев связаны изменения формы нейронов, особенно пирамидальных клеток. В тех местах, где протяжение данного слоя ограничивается и он становится более толстым, пирамидальные клетки становятся более длинными и более тонкими; там же, где протяжение данного слоя увеличивается и он делается более тонким, пирамидальные клетки становятся короче и шире. Самые малые нейроны не представляют заметных изменений формы, которые можно было бы связать с изменениями толщины корковых слоев. Нейроны средних слоев, особенно их пирамидальные клетки, также обладают сравнительно большим постоянством формы.
Независимо от только что описанных нами изменений, присущих каждой борозде и каждой извилине, в той или другой области коры наблюдаются не только изменения толщины коры, взятой в целом, но и изменения толщины отдельных ее слоев, которые могут становиться или расширенными, или суженными, или расслоенными.
И количество, и расположение как нервных клеток, так и нервных волокон также могут быть изменены.
На основе вышеуказанных изменений вся кора мозга была подразделена на структурно различные области. Карты структурных областей коры были составлены Campbell (1905), Elliot Smith (1907) и Brodmann (1909)*. Различаясь в деталях, эти карты в общем сходны между собой.
* ("Еще в 1881 г. проф. анатомии Киевского университета А. Ф. Бец показал, - говорит проф. Гринштейн, - что вся кора головного мозга человека может быть разделена на восемь полей, значительно отличающихся друг от друга по своей структуре. Бец описал структуру этих полей и указал границы их. Далее он указал на существование аналогичной картины и у животных. Работа эта, установившая основные факты цитоархитектоники коры, однако, не обратила на себя в то время внимания, и лишь через 25 лет после этого появились почти что одновременно работы Кемпбелла, Смита, Бродмана и Фогта, создавшие архитектонику коры как самостоятельную науку".)
По мере того как исследования коры мозга подвигались вперед, число структурных областей все увеличивалось. В 1913 г. Campbell насчитывал около 20 таких областей, позднее Brodmann довел число этих областей до 60 (рис. 252 и рис. 253), а в 1920 г. Rconomo насчитывал уже 107 таких областей. Многие из этих областей не отличаются друг от друга по своей структуре, но разделены между собой промежуточными областями иной структуры. В одних случаях борозды являются границами структурных областей, в других случаях этого не наблюдается. Иногда мы имеем ряд структурных областей на одной и той же извилине.
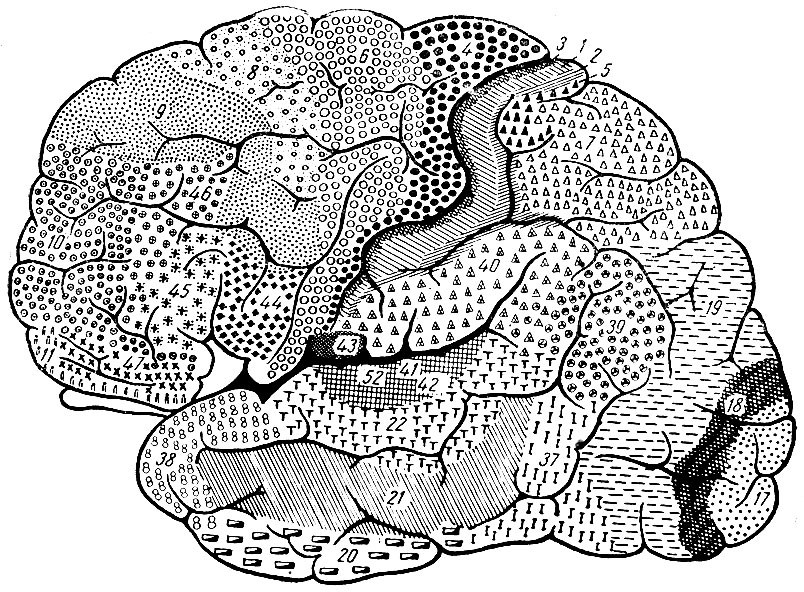
Рис. 252. Детальное подразделение коры мозга на отдельные поля, как это было определено Brodmann. Каждое поле, обозначенное соответствующей цифрой и соответствующим пунктиром, обладает определенной структурой его клеточных слоев и его нервных волокон. Наружная сторона полушария
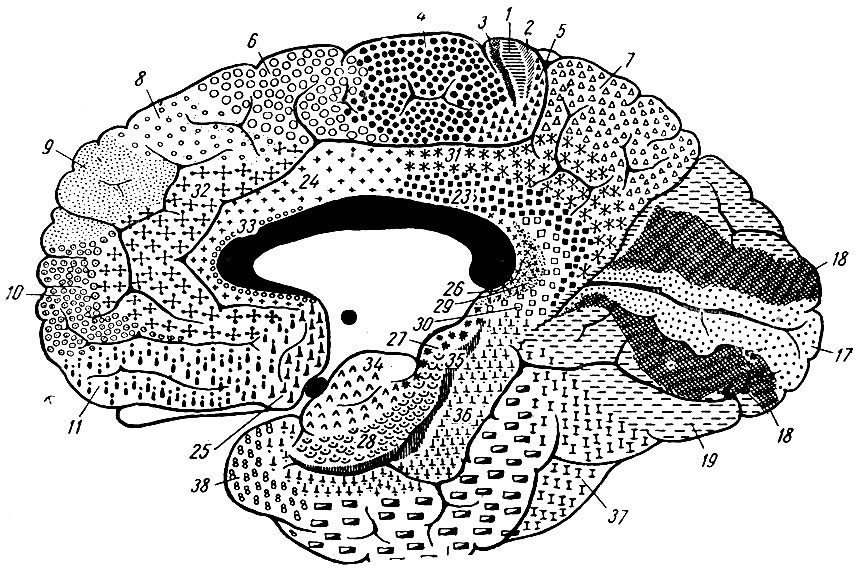
Рис. 253. То же. Внутренняя сторона полушария
Цитоархитектурные типы. Все то разнообразие гистологических структур, которое имеется на различных участках коры головного мозга, Economo свел лишь к пяти типам, из которых 2-й, 3-й и 4-й он считал основными и соответственно тем областям коры, где они имеют наибольшее распространение, назвал их: 1) фронтальный, 2) париетальный и 3) полярный типы. Тин 5-й он назвал гранулярным (зерновым), а тип 1-й - агранулярным (беззерновым) (рис. 254 и рис. 255).
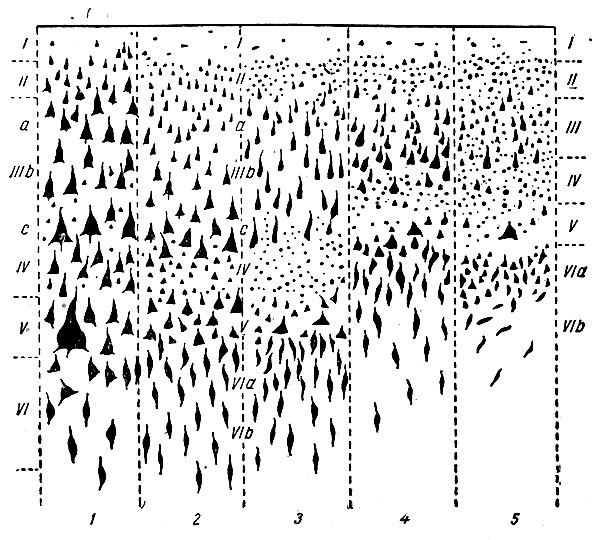
Рис. 254. Схематическое изображение 5 структуральных типов коры. Римские цифры обозначают соответствующие слои (по Economo)
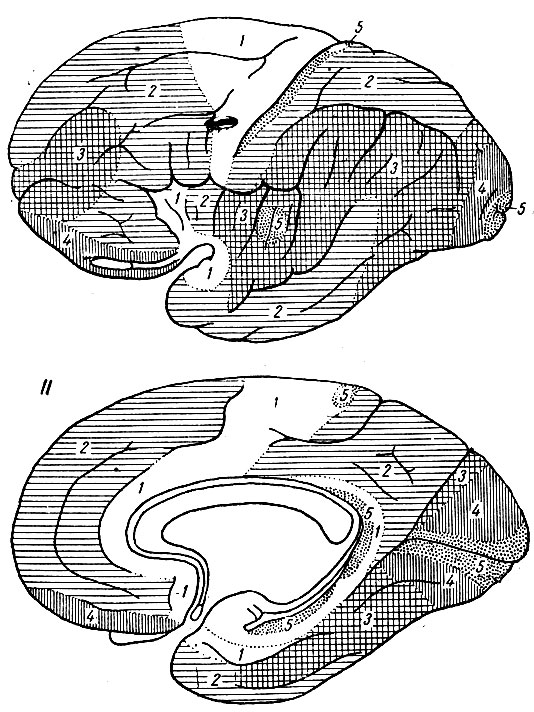
Рис. 255. Схема распределения пяти цитоархитектуральных типов коры на боковой и медиальной поверхностях мозгового полушария (по Economo)
Фронтальный тип характеризуется шестью типичными корковыми слоями. Кора этого типа сравнительно толста. Нейроны сравнительно большого объема, особенно пирамидальные клетки 3-го и 5-го слоев и полиморфные клетки 6-го слоя. Зерновидные клетки 2-го и 4-го слоев также сравнительно крупны и имеют по преимуществу треугольную форму. 2-й и 4-й слои не особенно богаты клеточными элементами.
Париетальный тип также представляет шесть слоев и имеет почти ту же толщину, как и фронтальный. Слои в париетальном типе резче отграничены друг от друга, чем во фронтальном. Это происходит от того, что наружный и внутренний слои зерен лучше развиты: они несколько толще и содержат в изобилии нервные клетки, которые представляются более округлыми, чем во фронтальном типе. Пирамидальные клетки 3-го и 5-го слоев менее крупны, более многочисленны, менее правильно расположены и менее типичны.
Полярный тип коры имеется на лобном и затылочном полюсах. Кора этих областей заметно тоньше, чем кора фронтального и париетального типов. Имеются все шесть слоев. Слоистость коры хорошо выражена. Пирамидальные клетки сравнительно невелики и по своей форме занимают промежуточное положение между соответствующими клетками фронтального и париетального типов.
Всю область коры, обладающую у взрослого или у зародыша 6 слоями клеток, Economo и Vogt называют isocortex. Небольшая часть коры, особенно обонятельный мозг, и у зародыша, и у взрослого обладает неполной слоистостью или представляет даже полное отсутствие слоистости. Такого рода кора носит название allocortex (alios - другой) и охватывает следующие участки: 1) подмозолистую извилину; 2) интралимбическую извилину вместе с indusium; 3) fasciola cinerea; 4) зубчатую извилину с аммоновым рогом 5) subiculum и presubiculum извилины морского коня; 6) крючковидную извилину; 7) извилины наружного и внутреннего обонятельных корешков и, по-видимому, переднее продырявленное пространство. У человека все перечисленные области составляют лишь 1/12 часть коры, тогда как у животных с сильно развитым обонянием (макросмических) они занимают очень большой отдел коры.
Как и isocortex, allocortex распадается на многочисленные поля с разнообразной структурой.
Шестислойную структуру isocortex не всегда бывает легко обнаружить. Тот или другой слой может утончиться почти до полного исчезновения, клетки данного слоя могут утерять свои характерные признаки, и остатки измененного слоя незаметно сольются с выше и нижележащими слоями.
Isocortex, видоизмененный вышеуказанным способом, носит название гетеротипического (heteros - по-гречески иной, другой) в отличие от isocortex, обладающего шестью ясно выраженными слоями, который носит название гомотопического (homos -по-гречески одинаковый, сходный).
Практически в коре человека различают лишь два вида гетеротипического isocortex: 1) агранулярный и 2) гранулярный.
Агранулярный тип получил свое название почти от полного отсутствия зерновидных клеток (2-й и 4-й слой). Агранулярный тип тесно связан с фронтальным типом, но величина клеток агранулярного типа, особенно пирамидальных клеток 3-го слоя, значительно превосходит величину клеток фронтального типа.
Самые крупные клетки коры, так называемые гигантские пирамиды, содержатся в 5-м слое агранулярного типа. Полиморфные клетки глубокого слоя (6-го) также имеют значительно большие размеры в агранулярном типе, чем во фронтальном. Так как зерновидные клетки в агранулярном типе сохранились лишь в самом незначительном количестве, то зерновидные слои лишь едва заметны. Соответствующие им зоны заняты главным образом пирамидальными клетками. Очевидно, говорит Economo, клетки 2-го и 4-го слоев, пережившие исчезновение зерен, изменили свой вид и приобрели форму и размеры средних и малых пирамидальных клеток.
Такого рода процесс Economo назвал пирамидализацией. В силу этого процесса получился незаметный переход 3-го слоя в 5-й. Особенно резко агранулярный тип выражен в предцентральной извилине.
Гранулярный (зерновидный) тип (5-й тип). В общем этот тип характеризуется малым объемом составляющих его клеток. Даже пирамидальные клетки 3-го и 5-го слоев сравнительно невелики по объему и имеются в небольшом количестве. Кроме пирамидальных клеток, оба эти слоя содержат также мелкие зерновидные клетки, которые имеются в таком большом количестве, что границы между отдельными слоями гранулярного типа коры становятся различимыми гораздо труднее, чем при каком-либо другом типе коры. 6-й слой содержит сравнительно небольшое количество полиморфных клеток. Общая толщина гранулярного слоя меньше, чем толщина других типов коры.
Как отличительной чертой 1-го типа Economo считает пирамидализацию, так отличительной чертой 5-го типа он считает "гранулизацию". Пятый тип коры Economo назвал "пылеобразным" (koniocortex). Классическим примером этого типа является шпорная борозда.
Проекционные центры. Некоторые области коры непосредственно связаны восходящими и нисходящими проекционными волокнами с особыми центрами промежуточного мозга. Такие области носят название проекционных центров коры. Каждый из проекционных центров коры состоит: 1) из центрального участка, где проекционные волокна особенно обильны, и 2) краевой зоны, с меньшим количеством проекционных волокон и большим числом ассоциационных волокон, при помощи которых этот корковый центр анатомически и физиологически связан не только с прилежащими областями коры, но и с более отдаленными корковыми областями.
Проекционный центр общей чувствительности (соматический) помещается на задней стенке (склоне) центральной борозды и на прилежащем участке поверхности позади центральной извилины. Этот центр получает волокна при помощи зрительнобугровой радиации от вентрального ядра зрительного бугра.
Зрительный проекционный центр помещается на стенках шпорной борозды и на прилежащих участках клина и язычной извилины. В некоторых случаях зрительный проекционный центр переходит через затылочный полюс на боковую поверхность полушария. Зрительный проекционный центр связан при помощи зрительной радиации с боковым коленчатым телом и с подушкой зрительного бугра.
Слуховой проекционный центр помещается на передней поперечной височной извилине, которая расположена на медиальной поверхности височной покрышки. Самый наружный отдел слухового проекционного центра появляется на боковой поверхности полушария близ середины верхнего края верхней височной извилины. При помощи слуховой радиации слуховой проекционный центр связан со срединным (медиальным) коленчатым телом.
Обонятельные проекционные центры помещаются: 1) в крючке; 2) в прилежащем отделе извилины морского коня и 3) в аммоновом роге. Крючок и прилежащий к нему отдел извилины морского коня представляют собой чувствующую обонятельную кору и получают свои приносящие волокна главным образом через боковой обонятельный тракт. Аммонов рог представляет собой выносящую обонятельную проекционную область. Выносящие обонятельные проекционные волокна выходят из этой области через бахромку и свод.
Двигательный проекционный центр коры помещается: 1) на передней стенке центральной борозды; 2) на прилежащей части предцентральной извилины и 3) на участке парацентральной дольки, расположенном тотчас же кпереди от верхнего конца центральной (роландовой) борозды и находящегося на медиальной поверхности полушария.
Ассоциационные центры. У человека двигательные и чувствующие проекционные центры занимают сравнительно небольшую часть мозговой коры. Остальная часть коры принадлежит так называемым ассоциационным центрам.
Агранулярный тип корковой цитоархитектоники, наилучше выраженный в двигательной проекционной области, как известно, не ограничивается пределами только этой облает, но распространяется на известном расстоянии кпереди на лобную долю. Точно так же и гранулярный корковый тип заходит за пределы чувствующих проекционных областей. Пограничные зоны проекционных областей таким образом структурно связаны с соответствующими проекционными областями и функционально зависят от этих областей. В общем, говорит проф. Kuntz, эти отношения становятся все менее и менее заметными по мере того, как расстояние от данного проекционного центра увеличивается. Ассоциационная кора, продолжает проф. Kuntz, может быть поэтому подразделена на первичные и вторичные ассоциационные зоны.
Первичные ассоциационные зоны представлены пограничными зонами, которые структурно связаны с соответствующими проекционными центрами и функционально подчинены этим центрам. Вторичные ассоциационные зоны располагаются в участках, ограниченных первичными ассоциационными зонами.
Дорзальный thalamus внутренней медуллярной пластиной делят на медиальный и латеральный отделы. Медиальный отдел филогенетически является более старым, а латеральный отдел - более новым. Однако нужно помнить, что в том и другом отделе имеются части и более старые, и более новые. Наружные и внутренние коленчатые тела, а также претектальное ядро низших позвоночных обычно присоединяют к латеральному отделу зрительного бугра. У человека претектальное ядро почти целиком втянуто в передний бугор четверохолмия. Медиальный отдел дорзального зрительного бугра4 содержит три группы ядер: 1) группу средней линии, 2) группу переднюю и 3) группу медиальную.
Боковые стенки 3-го желудочка, как известно, связываются между собой при помощи тяжа серого вещества большего или меньшего объема (massa intermedia, commissuramedia). Этот коммиссуральный отдел зрительного бугра содержит многочисленные ядра, которые носят название ядер средней линии; они лучше развиты у млекопитающих, стоящих ниже приматов, чем у приматов. При переходе от низших приматов к высшим ядра средней линии несколько уменьшаются. От чего происходит уменьшение этих ядер, которые, очевидно, ассоциируют между собой дорзальные отделы зрительных бугров, - сказать трудно. Может быть, это зависит от того, что с возрастанием преобладающей роли коры в нее переходят и высшие ассоциационные центры.
Передняя группа ядер филогенетически является старой группой, так как представители этой группы имеются у пресмыкающихся и у птиц, однако полного своего развития (дифференцировки) она достигает лишь у млекопитающих. Взятая в целом передняя группа ядер образует обонятельный корреляционный центр, получающий обонятельные и обонятельно-внутренностные импульсы при помощи мамилло-таламического тракта, а по-видимому, также и путем перивентрикулярной системы. Из передних ядер зрительного бугра импульсы направляются в поясную область коры (gyrus cinguli sive fornicatus), откуда они могут переходить на выносящие пути, а также могут достигать сознания.
Медиальная группа ядер зрительного бугра филогенетически, по-видимому, моложе, чем передняя группа ядер. Возможно, говорит A. Kappers, что классическое медиальное ядро получает небольшое количество прямых лемнисковых волокон (например, от тройничного лемниска), но его наиболее характерные приносящие связи образованы короткими межъядерными волокнами, приходящими от масс серого вещества (ядер) дорзального отдела зрительного бугра, окружающих медиальное ядро, и преимущественно от вентрального ядра и ядер средней линии, а при помощи перивентрикулярных волокон - от hypothalamus. Таким образом, медиальное ядро служит для корреляции проприоцептивных, экстероцептивных и интерцептивных импульсов. Экстероцептивные и проприоцептивные импульсы достигают медиального ядра жри посредстве вентрального ядра, обонятельно-внутренностные импульсы из гипоталамической области достигают медиального ядра при помощи перивентрикулярной системы, а обонятельно-соматические импульсы от habenula - при помощи хабекуло-таламического тракта. Коррелированные таким образом импульсы направляются: 1) или в подзрительнобугровую область для зрительнобугровых рефлексов, 2) или к полосатому телу для соответствующих рефлексов, 3), или наконец, при посредстве передней зрительнобугровой радиации внутренней капсулы к ассоциационным центрам коры лобной доли. Эта связь медиального ядра с корой лобной доли дает анатомическую основу для передачи чувствующего тона (комплекс приятного и неприятного), а отчасти и создает анатомическую основу личности (Tilney и Riley, Huber и Crosby).
Центромедиальное ядро (n. centromedianus) является по преимуществу ядром корреляции, связанным как с медиальным, так и с латеральным подразделениями зрительного бугра. Наибольшего развития это ядро достигает у высших млекопитающих.
Боковое ядро дорзального отдела зрительного бугра, незначительное у низших млекопитающих, постепенно увеличиваясь, достигает наибольшего объема и наибольшей дифференцировки у приматов (рис. 256). Это прогрессивное увеличение объема и дифференцировки тесно связано с прогрессивным увеличением ассоциационных центров коры, - сперва в теменной области, а затем в лобной. Подушка, представляющая собой дифференцированный задний отдел бокового ядра, имеет такое же отношение к зрительной ассоциационной области, какое боковое ядро имеет к теменной и лобной ассоциационным областям. Боковое ядро увязано с лобной и теменной ассоциационными областями как центробежными, так и центростремительными путями, а подушка такими же путями - с затылочной ассоциационной областью.
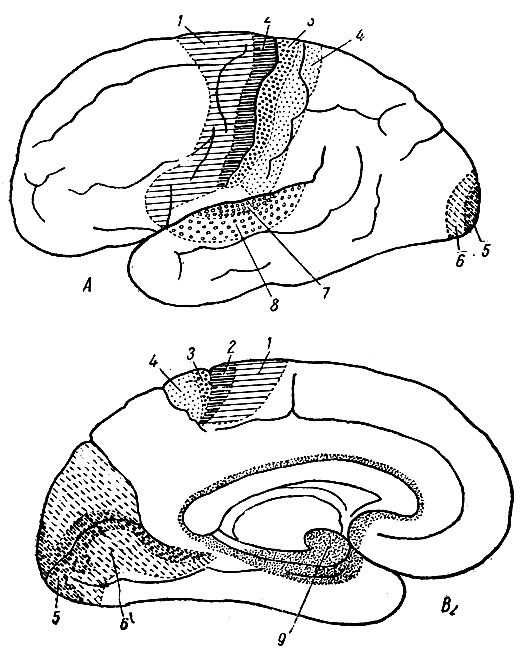
Рис. 256. Схема корковых проекционных областей на боковой и на медиальной поверхностях мозгового полушария) (по Kuntz). Обозначения на этих рисунках даны мной. 1 - прецентральная психомоторная (двигательная) область; 2 - прецентральная моторная область; 3 - постцентральная соместетическая область; 4 - постцентральная соместетико-психическая область; 5 - зрительно-сенсорная область; 6 - зрительно-психическая область (парастриальная и перистриальная области); 7 - слуховая сенсорная область; 8 - слуховая психическая область; 9 - обонятельная сенсорная кора
Вентральное ядро зрительного бугра является главным ядром, где оканчиваются проприоцептивные и экстероцептивные лемнисковые восходящие волокна и дентато-рубро-таламические пути. Получаемые вентральным ядром импульсы проецируются на предцентральную и позадицентральную извилины мозговой коры; импульсы, достигающие предцентральной извилины, по-видимому, являются проприоцептивными. Через вентральное ядро болевые, температурные, осязательные, проприоцептивные и вестибулярные импульсы доходят до сознания. Корково-таламические волокна, по-видимому, регулируют деятельность зрительного бугра. При помощи коротких нейронов вентральное ядро связано с латеральным и с медиальным ядрами.
Предцентральная (прецентральная) область. Предцентральной областью называют область, расположенную тотчас же кпереди от центральной (роландовой) борозды. Эта область по преимуществу выполняет двигательную функцию. Стимуляция определенных участков этой области вызывает определенные движения. У человека передняя граница области, отвечающей на стимуляцию, в ее вентральном отделе не распространяется дальше предцентральной борозды, но в ее дорзальном отделе может переходить за эту борозду, например, область движения глаз занимает каудальный отдел второй лобной извилины. Стимуляция этой области производит сопряженные боковые движения глаз и головы. Помимо 4-го поля Brodmann, содержащего гигантские пирамиды Беца, к предцентральной области относят также и 6-е поле Brodmann, не содержащее гигантских пирамид Веца. Campbell назвал это поле промежуточной прецентральной областью, a Millis - срединной лобной корой. В 6-м поле Brodmann, помимо области сопряженного движения глаз и головы, дающей на раздражение прямой двигательный ответ, имеются центры, в которых тонический характер выступает очень ясно, например, центры для эмоционального выражения радости, злобы (m. corrugator superciliorum, m. orbicularis oculi, мышцы крыльев носа). Возможно, что в 6-м поле имеются также и висцеральные центры, роль которых в эмоциях очевидна, например, центр слез, центр потоотделения. При повреждениях 6-го поля движения приобретают спазматический характер. Как было отмечено Hoff и другими авторами (привожу по A. Kappers), кортико-спинальные волокна, начинающиеся от 6-го поля Brodmann, у некоторых приматов спускаются до поясничной области.
Предцентральная извилина (передняя центральная) одной стороны связана при помощи волокон мозолистого тела не только с предцентральной извилиной другой стороны, но также и с позадицентральной извилиной другой стороны. При помощи длинных ассоциационных путей предцентральная область связана с теменной, с затылочной, а по-видимому, также и с височными областями коры. Многочисленные короткие связи соединяют предцентральную область с соседними прилежащими областями, в том числе и с областью лобной доли, содержащей зерновидные клетки.
На рис. 257, заимствованном мной у Ranson, изображена кортикальная локализация отдельных двигательных центров, расположенных на наружной поверхности полушария. На этот рисунок Ranson я нанес чувствующие и двигательные центры речи и двигательный центр письма.
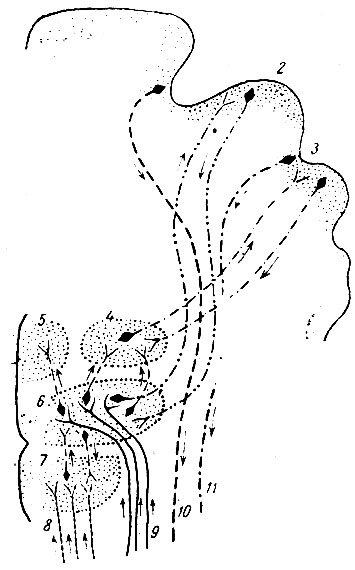
Рис. 257. Таламические, гипоталамические и корковые связи человеческого мозга (по Elliot Smith). 1 - двигательная кора; 2 - чувствующая кора; 3 - ассоциационная область; 4 - боковое ядро зрительного бугра; 5 - медиальное ядро (главный орган зрительного бугра); 6 - вентральное ядро; 7 - hypothalamus; 8 - висцеральные волокна; 9 - восходящие чувствующие волокна; 10 - путь к двигательным ядрам; 11 - путь к мозжечку
Regio frontalis granularis - область лобной доли, содержащая зерновидные клетки. В этой области возникают импульсы, связанные с высшей сознательной деятельностью. Филогенетически увеличение лобной доли у высших млекопитающих происходит главным образом за счет увеличения области, расположенной кпереди от предсильвиевой борозды, которая является эквивалентом s. arcuatus или s. praecentralis inferior. Особенно сильно эта область выражена у человека. Располагается regio frontalis кпереди от regio praecentralis. У хищников regio front, gran, выражена очень слабо. У обезьян обе указанных области приблизительно равны по объему. У шимпанзе и у человека гранулярная фронтальная область обширнее, чем прецентральная область. Sulcus praecentralis inferior служит у человека каудальной границей лобной гранулярной области. У высших млекопитающих, а особенно у человека, лобную гранулярную область подразделяют на вторичные поля, число которых у различных авторов различно. По Brodmann в состав лобной гранулярной области входят поля 8-12-е и 44-47-е. Слой зерен здесь выражен отчетливо за исключением переходного участка 8-го. Поля 44-е, 45-е и 47-е Brodmann объединяет в особую подобласть (subregio frontalis inferior), образующую 3-ю лобную извилину, верхней границей которой служит s. frontalis inferior, а задней границей - s. praecentralis inferior. Указанную подобласть левого полушария правшей считают центром артикулированной речи (кинестетические картины слов). Соответственно своему положению эта подобласть не обладает прямой двигательной функцией, которая присуща предцентральной агранулярной области. Непосредственная стимуляция центра речи третьей лобной извилины двигательного эффекта не вызывает. Разрушение этого центра показывает, что двигательная функция мышц, участвующих в произнесении артикулированных слов, не нарушена, но самая способность артикулировать слова (кинестетическая корреляция) отсутствует.
Развитие 3-й лобной извилины у человека вызывает окончательную оперкуляризацию островка, которая находит свое морфологическое выражение в прикрытии той области, которая расположена позади sulcus fronto-orbitalis.
Le Gros Clark утверждает, что гранулярный отдел лобной коры представляет собой не распространение кпереди моторной коры, а распространение кпереди области островка. Связь этого гранулярного отдела лобной доли коры с медиальной группой ядер зрительного бугра (paleo-thalamus) указывает на то, что эта связь очень древнего происхождения. В общем, как известно, медиальное подразделение дорзального отдела зрительного бугра связано с корреляцией экстероцептивных и проприоцептивных импульсов, причем обонятельно-висцеральные импульсы проходят из гипоталамических областей путем перивентрикулярной системы, а обонятельно-соматические импульсы - от habenula путем хабенуло-таламического тракта. Коррелированный таким образом материал направляется: 1) или в субталамическую область для таламических рефлексов, 2) или в полосатое тело, 3) или, наконец, в лобную долю коры. Эта связь с корой представляет собой анатомическую базу для чувствующего тона (комплекс приятного и болевого).
Термин "пирамидальный тракт" различные авторы употребляют в самом различном смысле. Первоначально этот термин указывал на то, что его спинальный отдел является продолжением пирамид продолговатого мозга. Flechsig (1877 и 1881) показал, что пирамидальный тракт связан с мозговой корой и преимущественно с прецентральной и парацентральной извилинами. Campbell (1905) на основании своих исследований утверждал, что пирамидальный тракт начинается исключительно от гигантских клеток Betz, расположенных внутри цитоархитектонически определяемой двигательной области (4-е поле Brodmann, поле FA-Economo, предцентральное поле Campbell). Однако большинство исследователей, наоборот, описывало пирамидальные волокна начинающимися от более обширной корковой области, чем цитоархитектонически определяемая двигательная область (4-е поле Brodmann). Так, Flechsig, полагает, что миэлогенетическая область, от которой начинается пирамидальный тракт, заходит вперед на премоторную область (6-е поле Brodmann). O. Vogt считает, что некоторые пирамидальные волокна начинаются от позадицентральной извилины. Основываясь на поражениях мозговой коры, Dejerine, Monakow, Minkowski описывают пирамидальные волокна, начинающиеся как от премоторной, так и от позадироландовой областей. Economo и Koskinas приходят к заключению, что клетки Betz не являются единственными клетками, которые дают начало пирамидальному тракту, и что область начала этого тракта, по-видимому, гораздо обширнее, чем цитоархитектонически определяемая двигательная область.
В 1889 г. Brown-Sequard опубликовал, свою экспериментальную работу, проделанную на кроликах и собаках. Он рассекал у этих животных или только пирамиды продолговатого мозга или всю толщу продолговатого мозга, за исключением пирамид. Он нашел, что стимуляция мозговой коры после рассечения пирамид вызывала почти такие же ответы; как стимуляция до рассечения пирамид. Когда рассекалась вся толща продолговатого мозга, за исключением пирамид, стимуляция коры также вызывала ответ, но уже значительно ослабленный. На основании своих опытов Brown-Sequard пришел к заключению, что пирамидные пути не только не являются единственными путями, при помощи которых наступают вышеописанные реакции, но они не являются при этом даже главными, и что связи между корой и спинным мозгом более многочисленны, чем это обычно допускают. При вышеописанных опытах дело шло при раздражении мозговой коры о движениях конечности противоположной стороны (контралатеральные движения). Позднее, при раздражении мозговой коры были отмечены движения, происходящие на той же стороне, на которой производилось раздражение (гомолатеральные движения). Некоторые авторы полагали, что гомолатеральные движения возникали путем передачи раздражения с двигательной области одного полушария при посредстве мозолистого тела двигательной области другого полушария, которая и вызывала вышеуказанные движения как контралатеральные. Exner, однако, показал на кроликах, что гомолатеральный ответ не изменялся не только после рассечения мозолистого тела, но даже после полного удаления противоположного мозгового полушария. На основании этих опытов Exner пришел к заключению, что иннервация гомолатеральной конечности кролика идет прямо от стимулируемого участка коры.
Hering в своих опытах на собаках показал, что, наряду с пирамидальным трактом, должен существовать "второй кортико-фугальный путь" и что этот путь должен перекрещиваться выше продолговатого мозга. Было найдено, что этот "второй путь" проходит вместе с пирамидальными волокнами в боковом столбе спинного мозга. Rothmann показал, что этим "вторым путем" является руброспинальный тракт. Probst многочисленными опытами на кошках показал, что полное повреждение внутренней капсулы уничтожает все ответы на стимуляцию двигательной коры той же самой стороны. Полное рассечение одной половины среднего мозга краниально к красному ядру вызывало полное уничтожение ответа. Если разрез ножки мозга захватывал только пирамидальный и корково-мостовый тракты, оставляя нетронутым путь к красному ядру, - реакция сохранялась, хотя для этого требовался более сильный ток. В конце концов, Probst пришел к заключению, что и пирамидальный, и руброспинальный тракты являются важными путями при вызывании у кошек контралатеральных ответов стимуляцией двигательной коры. Leyton и Sherrington (1917) описали у шимпанзе двустороннее перерождение в боковых столбах спинного мозга, развившееся после одностороннего повреждения двигательной коры. Операции были произведены на трех животных; у одного была удалена корковая область верхней конечности, а у двух - корковая область нижней конечности. При обследовании спинного мозга оперированных животных было обнаружено не только перерождение пирамидальных волокон в боковом столбе противоположной стороны, но также и перерождение волокон в переднем и боковом столбах той же самой стороны. В опытах, захватывающих нижние конечности, волокна неперекрещенного бокового пирамидального тракта были прослежены до треть его крестцового сегмента. Fulton и Keller обследовали в спинном мозгу одного шимпанзе перерождение, наступившее после удаления корковой двигательной области левой нижней конечности. Они подтвердили наблюдения Leyton и Sherrington и определили почти в 10% число перерожденных неперекрещенных пирамидальных волокон, которые проходили в заднебоковом отделе бокового столба. Kennard и Ной показали, что у обезьян кортико-спинальные тракты той же стороны начинаются как в моторной, так и в премоторной корковых областях и что моторная область подтает свои ипсилатеральные волокна как в боковой, так и в вентральный (передний) спинномозговые столбы той же самой стороны, тогда как ипсилатеральные волокна от премоторной области находят только в боковом столбе. В общем тотальная ипсилатеральная дегенерация от премоторной области коры сравнительно больше, чем от моторной области. Кроме того, Hoff показал также, что значительная часть нисходящих ипсилатеральных трактов заканчивается на той же самой стороне, а не переходит в нижележащих сегментах на противоположную сторону спинного мозга, как предполагали некоторые авторы. John Fulton и Donal Sheehan удалили у молодого самца шимпанзе двигательную корковую область левой ноги, определив при операции границы этой области стимуляцией фарадическим током. Гистологическое обследование этого случая показало, что волокна от 4-го поля Brodmann спускались по той же стороне до пирамидального перекреста в продолговатом мозгу. На этом уровне указанные волокна делились на три пучка, из которых наибольший пучок (1) переходил на противоположную сторону и спускался вниз в боковом столбе спинного мозга. Второй пучок (2), соответствующий по величине приблизительно десятой части первого пучка, спускался в дорзальном отделе бокового столба той же самой стороны. Небольшой пучок волокон (3) спускался в вентральном столбе той же самой стороны, лежа возле передней центральной борозды спинного мозга (прямой или вентральный пирамидальный тракт). Ни в одном сегменте спинного мозга в передней коммиссуре не было обнаружено перерожденных волокон. Таким образом, не удалось подтвердить гистологически, что прямой пирамидный тракт, в конце концов, переходит на противоположную сторону. Прямой вентральный пирамидальный тракт можно было проследить до нижних грудных сегментов, а прямой боковой пирамидальный тракт можно было проследить на своей стороне до крестцовой области.
При использовании материалов сайта активная ссылка обязательна:
http://anfiz.ru/ 'Анатомия и физиология человека'