Ядра и корешковые волокна головных нервов
Ядра головных нервов, начиная от III пары (n. oculomotorius) и кончая XII (n. hypoglossus), соответствующие ядрам спинных нервов, заложенных в серых столбах спинного мозга, находятся в сером веществе продолговатого мозга, варолиева моста и сильвиева водопровода. Корешковые волокна этих нервов входят как часть в состав forn ationis reticularis albae покрова. Но так как анатомия ядер и корешков головных нервов представляет важное практическое значение в невропатологии, то она и будет изложена здесь в дополнение к описанию tegmenti.
Серое вещество спинного мозга, заключающее в себе ядра движущих и чувствующих корешков спинномозговых нервов, можно представить себе в виде пластинки, обогнутой вокруг центрального канала спинного мозга. Серое вещество, покрывающее дно IV желудочка, можно рассматривать в таком случае как пластинку, разогнутую и не окружающую всей периферии канала, здесь расширенного и являющегося в качестве полости IV желудочка. Состав этой пластинки, однако, остается тот же, что и в спинном мозгу, т. е. он содержит отдельные скопления нервных клеток - ядра головных нервов. Расположение этих ядер изменилось настолько, насколько изменилась форма всей пластинки: как сказано, вместо сомкнутой в виде трубки вокруг канала серая пластинка в продолговатом мозгу представляется разостланной на дне IV желудочка. Часть ее, образующая в спинном мозгу задние серые столбы, расположилась кнаружи от средней линии, часть, образующая в спинном мозгу передние серые столбы, осталась на месте - около средней линии - впереди полости. Поэтому ядра движущих нервов (соответствующих ядрам двигательных корешков спинного мозга) не переместились: они лежат по-прежнему впереди центрального канала, по бокам средней линии IV желудочка и образуют правильный ряд, начиная от calamus scriptorius до переднего конца сильвиева водопровода. На схеме (рис. 103) представлено их относительное расположение (ядра движущих головных нервов для большей наглядности заштрихованы, а ядра чувствующих - нет).
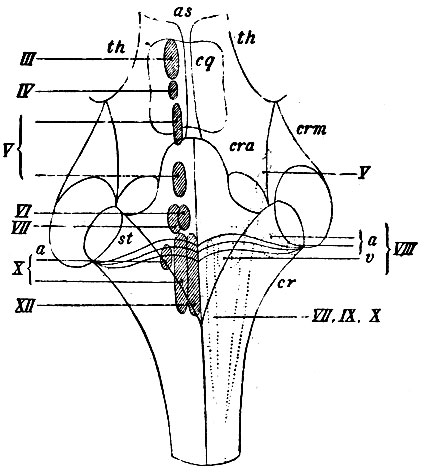
Рис. 103. Схематическое изображение расположения ядер головных нервов. as - сильвиев водопровод; th, th - зрительные бугры; cq - четверохолмие; cra - передние ножки мозжечка (crura cerebelli ad corpus quadrigeminum): crm - средние ножки мозжечка (crura cereb. ad pontem Varolii)); cr - corpora restiformia (crura cereb. ad medull. oblong.). На левой стороне рисунка - ядра двигательных нервов (заштрихованы): III - ядро n. oculomotorii; IV - ядро n. trochlearis; V - два ядра двигательного корешка n. trigemini; VI - ядро n. abducentis; VII - ядро n. facialis; X - два ядра двигатзльной части n. vagi; XII - ядро n. hypoglossi. На правой стороне рисунка - ядра чувствующих нервов (обозначены пунктиром); V - ядро чувствующего корешка n. trigemini; VIII - ядра n. acustici; a - два ядра rami cochlearis; v - два ядра rami vestibularis; VII, IX, X - ядро n. intermedii Wrisbergii, n. vagi, n. glosso-pharyngei
В этом ряду лежат последовательно, начиная сзади, ядра n. hypoglossi (XII), движущей части n. vagi (X), n. facialis (VII), n. abducentis (VI), ядро движущего корешка n. trigemiui (V), n. trochlearis (IV) и n. oculomotorii (III). Ядра чувствующих нервов по причине перемещения задних частей серой пластинки расположены кнаружи от движущих и несколько глубже их; здесь, образуя второй ряд, лежат ядра чувствующей части n. vagi (X), n. glosso-pharyngei (IX), n. facialis (VII), ядра n. acustici(VIII) и ядра чувствующего корешка n. trigemini (V).
Ядра эти и нервные корешки, находящиеся в связи с ними, однако, трудно разгруппировать попарно, как они разгруппированы в спинном мозгу. Это зависит, разумеется, от более сложного дифференцирования головной части мозга и самой головы, анатомическое устройство которой можно подвести под тип сегментов туловища только в общих чертах, но не в частностях.
Ядро XII пары n. hypoglossi.
Ядро подъязычного нерва (рис. 104, nХII), состоящее из больших мультиполярных клеток, заложено под нижним концом funiculi teretiïs дна IV желудочка, на некоторой глубине, около самой средней линии, таким образом, что нижний конец его переходит несколько предел ромбовидной ямки, т. е. верхушку писчего пера, а верхний лежит на уровне слуховых полосок (striae acusticae). Оно занимает, таким образом, половину протяжения fossae rhomboideae. Поперечник его невелик, и потому в сторону от него остается место для ядра n. vagi (так наз. ala cinerea). Корешки n. hypoglossi выходят из нижней стороны ядра в виде многочисленных пучков, которые пронизывают вещество продолговатого мозга сверху вниз и выходят (рис. 104; VII) на поверхность его, на границе между оливой и пирамидой. Так как нерв этот чисто двигательный, то волокна его происходят, как отростки, из клеток ядра.
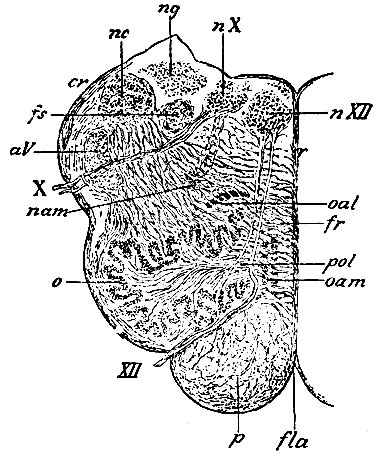
Рис. 104. Поперечный разрез продолговатого мозга приблизительно на середине оливы (правая половина). nХII - ядро n. hypoglossi; nХ - ядро n. vagi; XII - корешковые волокна и выход n. hypoglossi; X - корешковые волокна и так называемый выход n. vagi; лат - nucleus ambiguus; o - nucleusdentatus olivae; oal - наружное прибавочное ядро оливы; oam - внутреннее прибавочное ядро оливы; ng - ядро funiculi gracilis; nc - ядро funiculi cuneati; aV - нисходящий корешок n. trigemini; fs - funiculus solitarius или дыхательный пучок Krause, входящий в состав n. vagi; cr - corpus restiforme; p - пирамидный пучок, в котором близ поверхности видны так называемые fibrae arcuatae externae; fr - formatio reticularis tegmenti, пронизанное внутренними дугообразными волокнами (fibrae arcuatae internae); pol - ножка оливы (pedunculus olivae); r - raphe; fla - fiss. longitudinalis anterior
Ядро виллизиева нерва, XI пара. Описание его требует предварительной оговорки о том, что нужно понимать под именем прибавочного виллизиева нерва. Долгое время под именем XI пары (n. accessories Willisii) разумели сложный нерв, который начинается частью в сером веществе спинного мозга и выходит несколькими корешками из поверхности боковых столбов в шейном его отрезке, частью в сером веществе продолговатого мозга из ядра, общего для XI-X пар, и выходит также несколькими нитями из боковой поверхности продолговатого мозга. Первую часть называли спинной, вторую - головной частью виллизиева нерва. Сравнительно недавно Holl обратил внимание на то, что такое понимание не соответствует первоначальному описанию этого нерва, сделанному английским анатомом Willis, именем которого и назван нерв. Willis назвал прибавочным нервом только то, что впоследствии именовалось спинной частью этого нерва, т. е. часть, выходящую из серого вещества спинного мозга и представляющую чисто двигательный нерв. Так называемую головную его часть он считал принадлежащей к блуждающему нерву. Такого же взгляда держался Claude Bernard. Это разделение лучше соответствует способу периферического разветвления обоих нервов и потому в настоящее время усваивается более и более.
Что касается положения ядра спинной части n. access. Willisii, то в настоящее время всеми признаны наблюдения Роллера (Roller) и Даркшевича. Ядро это заложено в переднем роге серого вещества шейного отдела спинного мозга кнаружи и кзади от ядер передних корешков спинных нервов. Нижний конец этого ядра лежит на уровне V шейного корешка, верхний - на уровне задней трети оливы. Корешковые волокна n. Willisii образуют дуги, обращенные выпуклостью назад, и проходят сквозь массу боковых и пирамидных пучков, чтобы выйти наружу.
Ядра n. vagi (X пара), n. glosso-pharyngei (IX пара) и чувствующего к орешка n. facialis, так наз. portio intermedia Wrisbergii.
Эти ядра могут быть описаны только вместе, так как некоторые из них общи всем этим нервам.
Nn. vagus и glosso-pharyngeus в своих стволах содержат двигательные и чувствующие волокна, не разделенные в особые ветви, но соответствующие ядра обособлены.
Двигательных ядер по современному учению (Vincenti, v. Gehuchten, Алфеевский) два. Одно, большее, лежит на дне IV желудочка рядом с ядром n. hypoglossi и известно в макроскопической анатомии под именем ala cinerea (рис. 104, nХ). Другое, меньшее, лежит под главным в массе subst. reticularis продолговатого мозга. Оно известно под именем nucleus ambiguus (рис. 104, nam). Волокна, происходящие от клеток этого ядра, выходят к главному ядру и смешиваются с волокнами, происходящими от последнего. Те и другие образуют многочисленные пучки, которые пронизывают наискось массу tegmenti продолговатого мозга (рис. 104, X) и выходят на поверхность вместе с чувствующими корешками этого нерва. В числе этих пучков имеются некоторые, входящие в состав ствола n. glosso-pharyngei. С корой противоположного полушария мозга (Перекрестно) описанные ядра связаны волокнами пирамидного пучка (см. схему рис. 114).
Ядра чувствующих корешков n. vagi, glosso-pharyngei и portionis Wrisbergii, подобно ядрам чувствующих корешков спинных нервов, лежат вне мозга, в узлах, которые имеются на стволах этих нервов (описаны ниже в отделе анатомии периферических нервов).
N. vagus имеет два таких ядра. Одно, большее, лежит в массе утолщения ствола n. vairi на шее и носит название ganglion nodosum. Другое, меньшее, заложено в узле, носящем название ganglion jugulare, лежащем в соименном отверстии черепа, через которое нерв выходит.
Ядро n. glosso-pbaryngei заложено в узле, носящем название ganglion petrosum, который находится на стволе нерва в том месте, где ствол выходит из черепа (также for. jugulare).
Ядро n. Wrisbergii лежит в узле, который имеется па стволе n. facialis в том месте, где он в полости canalis Falopiae височной кости образует колено (перегибается под прямым углом). Узел этот носит название ganglion genieuli. Клетки всех этих узлов, так же как клетки спинных узлов, дают отростки, которые затем делятся па две ветви; одна из них направляется в периферию, другая в мозг в составе корешков нерва. Войдя в массу продолговатого мозга, они входят в состав круглого канатика, известного под названием funiculus solitanus (рис. 104, fs), иначе - пучок Krause, который направляется вниз в шейный мозг и, истончаясь, оканчивается на уровне VIII шейного нерва. Волокна, его составляющие, делятся на восходящие и нисходящие веточки и оканчиваются коллатералями около клеток задних рогов спинного мозга, мимо которых проходит funiculus solitarius. Второе звено цепи, соединяющее тотчас описанное ядро продолговатого мозга с головным мозгом, неизвестно, но несомненно, что волокна, которые начинаются от клеток ядра, перекрещиваются с волокнами противоположной стороны в raphe продолговатого мозга и затем в составе петли (lemniscus) оканчиваются в зрительном бугре.
Ядро слухового нерва. Восьмая пара, n. acusticus, чувствующий нерв, состоит из двух частей или ветвей различной функции. Более толстая часть ствола назначена для улитки слухового органа - это ramus cochlearis; она представляет собственно слуховой нерв. Более тонкая ветвь назначена для преддверия и полукружных каналов слухового лабиринта, который представляет собой аппарат совершенно другого значения - аппарат, входящий в состав сложного механизма самоопределения в пространстве и соблюдения равновесия тела. Ветвь нерва, его снабжающая, носит название ramus vestibularis.
Каждая из этих двух ветвей имеет в продолговатом мозгу особые вторичные ядра (по два у каждой).
Ramus cochlearis состоит из волокон, которые начинаются от клеток очень маленьких узелков, лежащих в улитке лабиринта. Как у всех чувствующих волокон, отросток клетки делится на две ветви - периферическую и центральную. Периферическая ветвь направляется к концевому аппарату, так называемому кортиеву органу лабиринта. Центральная идет в продолговатый мозг, чтобы войти в соотношение с вторичным ядром. Эти волокна rami cochlearis огибают corpus restifome по наружной стороне и оканчиваются около клеток двух ядер, из которых одно лежит под нижним краем corporis restiformis (рис. 105, na) и носит название переднего, или вентрального; другое расположено на наружной поверхности corp. restiformis (рис. 105, tu) и носит название слухового бугорка (tuberculum acusticum) или дорзального ядра. В каждом из этих ядер возникают новые волокна, составляющие второе звено цепи - собственно проводящие к мозгу пути. Пучок таких волокон, возникающий из переднего ядра, проникает в массу subst. reticularis продолговатого мозга, в raphe переходит на другую сторону и в составе петли (lemniscus) восходит кверху, чтобы окончиться в ядрах заднего возвышения четверохолмия и наружного коленчатого тела. Пучок волокон, возникающий в клетках tuberculi acustici, огибает corpus restiforme снаружи (он виден невооруженным глазом на свежем мозгу) и, достигнув дна IV желудочка, образует striae acusticae макроскопической анатомии. На средней линии strae acusticae также переходят на другую сторону (перекрещиваются) и затем идут тем же путем, как и пучок от переднего ядра (lemniscus); оканчиваются так же. Третье звено цепи, соединяющее ядра четверохолмия и коленчатого тела с корой (первой височной извилины), неизвестно, но, конечно, существует и направляется в составе внутренней капсулы.
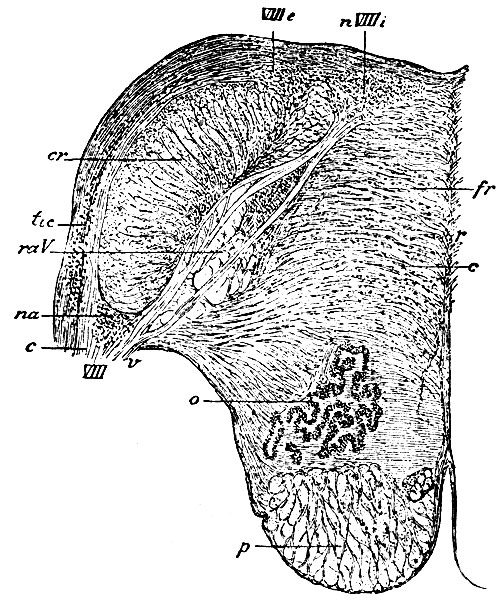
Рис. 105. Разрез продолговатого мозга на уровне striae acusticae. VIII - главное или мелкоклеточное ядро n. acustici; nVIIIe - заднее наружное ядро; na - переднее или вентральное ядро: tic - tuberculum acusticum; VIII - n. acusticus; c - его ramus cochlearis; v - его ramus vestibularis; raV - нисходящий корзшок чувствительной части n. trigemini; o - nucleus dentatus olivae; cr - corpus restiforme; fr - formatio reticularis; r - raphe; c - продольные волокна formationis reticularis и рассеянные клетки его; p - пирамиды
Ramus vestibularis состоит из волокон, которые начинаются в небольших узелках, лежащих в лабиринте. Центральные ветви их проникают в продолговатый мозг под нижним краем corp. restiiormis и, дойдя до дна IV желудочка, входят в соединение с двумя ядрами, расположенными у наружного края ромбовидной ямки. Одно из них состоит из мелких клеток, лежит ближе к середине и носит название главного ядра (рис. 105, nVIIIi); другое, крупноклеточное, помещено кнаружи от предыдущего и носит название задне-наружного (рис. 105, VIIIe). Здесь из обоих ядер возникают волокна второго звена, которые разделяются первоначально на два пучка (Cajal, v. Gehuchten), восходящий и нисходящий. Первый, не переходя на другую сторону, направляется к ядрам нервов, движущим глаз (oculomotorius, troehlearis). Второй идет сначала внутрь в составе subst. reticularis tegmenti и опять делится на две части. Большая часть, оставаясь на той же стороне, спускается вниз через весь продолговатый мозг, затем переходит в спинной мозг, где, находясь в массе передне-бокового столба, оканчивается около движущих ядер всего тела. Меньшая часть, идя поперечно к средней линии, частью переходит в другую сторону, частью остается на той же стороне и затем, в составе правого и левого (рис. 108 и 109, flp) задних продольных пучков, проникает в пирамидный пучок своей и противоположной стороны. Другая часть волокон, заложенная в задние продольные пучки, напротив, направляется кверху и оканчивается в зрительном бугре.
Нужно отметить, что большинство волокон этого второго звена цепи не переходит на другую сторону. Далее v. Gehuchten отрицает существование третьего звена этой цепи n. vestibularis, т. е. волокон, которые соединяли бы ядра n. vestibularis с корой мозга. Этим и объясняется, что весь механизм равновесия бессознателен и совершается исключительно рефлекторным путем.
Ядра и корешки nervorum facialis (VII пара) и abducentis (VI пара). Ядра движущих нервов лежат рядом под круглым канатиком (funic, teres) IV желудочка, тотчас впереди слуховых полосок. Ядро отводящего нерва лежит ближе к поверхности ромбовидной ямки (рис. 106. nVI), ядро лицевого нерва - под ним на глубине 4-5 миллиметров и несколько далее от средней линии. Корешок n. facialis слагается, по-видимому, из двух пучков волокон: 1. Главный пучок происходит от ядра с верхней стороны и направляется ко дну IV желудочка; затем он заворачивает кнутри и проходит между поверхностью fossae rhomboideae и ядром отводящего нерва. Далее он под прямым углом поворачивает вперед и, пройдя несколько в соседстве средней линии, вновь заворачивает кнаружи (genu cerebrale n. facialis, gVII), огибает снова ядро abducentis и потом направляется вниз сквозь массу продолговатого мозга к заднему краю варолиева моста. 2. Второй корешок, как думают некоторые, происходит из ядра противоположной стороны, и пройдя через raphe, присоединяется к колену главного корешка. Этот второй корешок другими авторами рассматривается как мозговой путь, соединяющий ядро facialis с корой мозга, который идет, как у всех движущих нервов, в составе пирамидного пучка.
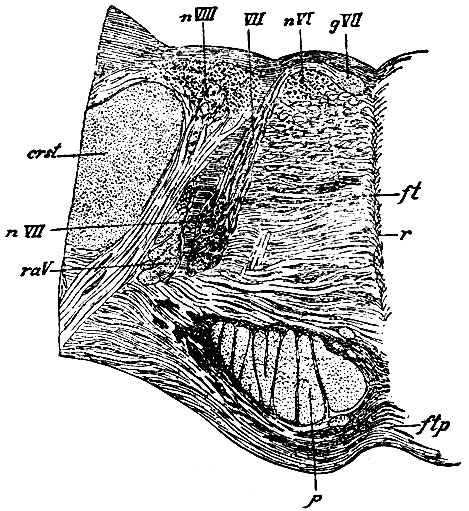
Рис. 106. Разрез через ядра лицевого и отводящего нервов. nVII - ядро n. facialis; VII - восходящая часть корешка n. facialis; gVII - колено n. facialis, перерезанное поперек в той его части, которая идет вперед; nVI - ядро n. abducentis; raV - нисходящий корешок чувствительной части n. trigemini, перерезанный поперек: p - пирамидальный пучок: nVIII - ядро n. acustici; crst - corpus restiforme
Верно только то, что на препаратах, окрашенных по способу Weigert, он хорошо виден на протяжении от raphe до колена n. facialis, но связь его с ядром противоположной стороны неясна: пройдя raphe, он рассыпается в сеть и исчезает из виду.
N. abducens происходит из описанного выше ядра (рис. 106, nVI). Корешки его появляются из внутренней стороны ядра и идут прямо вниз сквозь formatio reticularis, чтобы появиться на поверхности мозга позади края варолиева моста.
По исследованиям Duval, ядро n. abducentis служит вместе и для происхождения некоторых волокон n. oculomotoni. Такое значение ядра n. abducentis объясняет случаи совместного паралича m. recti externi oculi одной стороны и m. recti interni другой стороны при поражении одного только ядра отводящего нерва.
Ядра и корешки n. trigemini (V пара). Нерв этот смешанный: он состоит из большей - чувствующей части (задний корешок) и меньшей - двигательной (передний корешок). Волокна чувствующей части происходят из клеток гассерова узла, лежащего на стволе нерва в полости черепа. Как у всех чувствующих нервов, волокна эти еще в узле делятся на две ветки, из которых одна идет к периферии. другая в мозг. Пучок центральных ветвей, войдя в мозг, частью оканчивается коллатералями около клеток ядра, лежащего в массе tegmenti варолиева моста, над местом входа нерва в мозг (рис. 107, s), частью направляется вниз, образуя так наз. нисходящий корешок нерва (прежде он назывался восходящим) (рис. 105, 106, raV), который, уменьшаясь мало-помалу в толщине, спускается в спинной мозг и входит в соотношение с серым веществом задних рогов до уровня VI шейного нерва (?). Второе звено чувствующей цепи возникает из клеток описанного ядра, переходит в raphe варолиева моста на другую сторону и присоединяется к общему тракту чувствующих проводников, т. е. к петле (lemniscus).
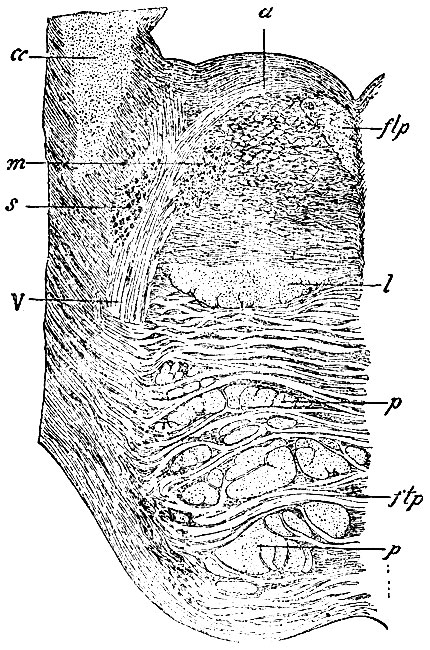
Рис. 107. Разрез, проведенный через так наз. locus coeruleus IV желудочка и через ядра trigemini, заложенные в этом месте. m - движущее ядро n. trigemini; s - чувствующее ядро n. trigemini; V - корешок n. trigemini (который попал в разрез не на всем своем протяжении); d - перекрещенный корешок trigemini; flp - задний прямой пучок покрова, который начинает выделяться из массы formationis reticularis; l - lemniscus (петля); p - пирамидный пучок, разбитый поперечными волокнами моста (ftp) на отдельные группы; cc - corpus restiforme
V. Gehuchten считает этот путь соответствующим путям голлева и бурдахова столбов спинного мозга, т. е. проводниками мышечного чувства, так как он не заворачивает в мозжечок. Но так как n. trigeminus несомненно проводит все виды кожного чувства, то должен существовать еще путь с заворотом в мозжечок.
Ядро двигательного корешка n. trigemini лежит несколько впереди от ядер лицевого и отводящего нервов, под так наз. locus coeruleus IV желудочка (также над местом появления корешка на поверхности варолиева моста) (рис. 107, m). Оно отделено от поверхности слоем темно пигментированных клеток, которые и обусловливают темный цвет этого участка дна желудочка. Корешок, выходящий из ядра, направляется вниз сквозь массу варолиева моста (рис. 107, V). Henle, Kölliker, Понятовский и в настоящее время почти все авторы признают существование еще другого ядра двигательного корешка тройничного нерва в виде рассеянных клеток, заложенных под серым веществом сильвиева водопровода (рис. 108, n). Волокна, происходящие из этих клеток, образуют нисходящий корешок нерва (рис. 108 и 109, rV), который идет вниз (или, все равно, назад) по направлению к IV желудочку. Мозговые проводники, соединяющие эти ядра с корой мозга, находятся в пирамидном пучке противоположной стороны. Перекрест их находится в raphe tegmenti.
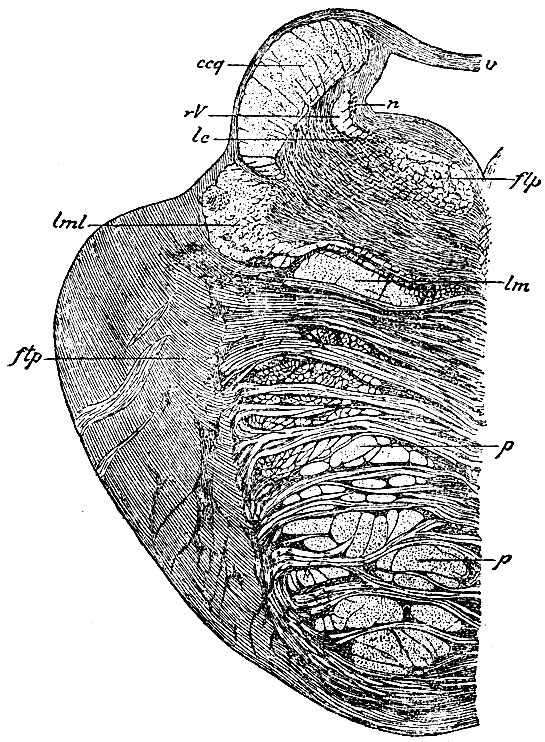
Рис. 108. Разрез, проведенный несколько впереди от предыдущего (рис. 107). rV - нисходящий корешок движущей части n. trigemini; n - клетки, образующие его ядро; lc - клетки loci coerulei; flp - задний прямой пучок покрова, который уже окончательно сложился; lml - боковой пучок петли; lm - средний пучок петли; p, p - пирамидный пучок: ccq - волокна ножки мозжечка к четверохолмию; ftp - поперечные волокна моста; v - передний мозговой парус
Ядро и корешки n. trochlearis, IV пара. Ядро этого нерва заложено в сером веществе, составляющем дно сильвиева водопровода, на уровне поперечной борозды четверохолмия. Корешок (рис. 109, rIV), выходящий из него, идет назад под слоем серого вещества водопровода и, дойдя до заднего края corporis quadrigemim, вступает в ткань переднего мозгового паруса, где образует перекрест (может быть, неполный). После этого он появляется на наружной поверхности паруса в виде уже сложившегося нерва, видимого невооруженным глазом.
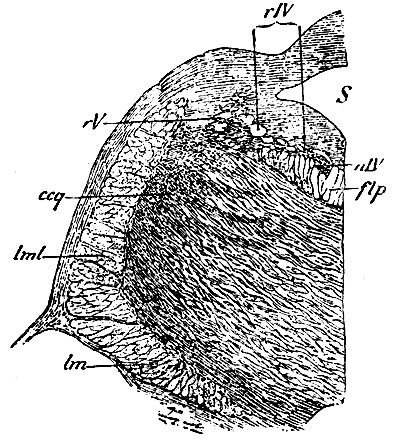
Рис. 109. Разрез, проведенный через ядро n. trochlearis. S - aquaeductus Sylvii; nIV - ядро n. throchlearis; rIV - корешок n. trochlearis; rV - двигательный корешок n. trigemini, перерезанный поперек (точки около него обозначают клетки); flp - задний прямой пучок покрова; ccq - волокна ножки мозжечка к четверохолмию; lml - боковой пучок петли (lemniscus макроскопической анатомии); lm - средний пучок петли
Ядро и корешки n. oculomotorii, III пара. Сравнительно большое ядро (5 мм длины) заложено так же, как предыдущее, в сером веществе дна сильвиева водопровода, на уровне переднего возвышения четверохолмия. Многочисленные корешки этого нерва (рис. 110, rIII), выйдя из нижней стороны ядра, направляются прямо вниз сквозь массу ножек мозга, на внутреннем крае которых они выходят наружу, слагаясь в один ствол уже после выхода. Часть волокон происходит из ядра другой стороны, так что между ядрами (в задней части их) правой и левой стороны виден перекрест волокон.
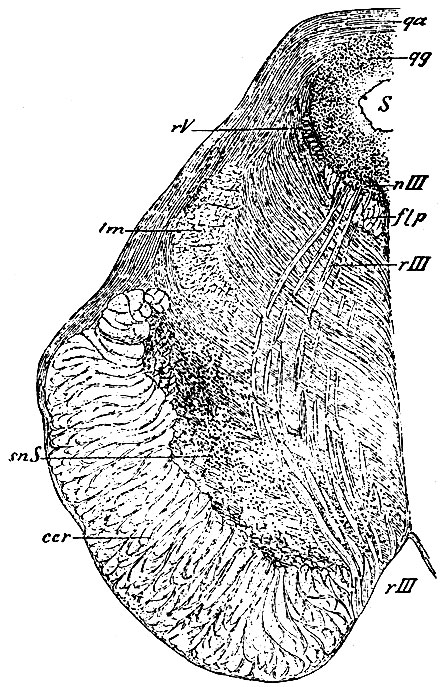
Рис. 110. Разрез проведен через ядро и корешки n. oculomotorii. nIII - ядро oculomotorii; rIII - корешки oculomotorii; rV - нисходящий корешок n. trigemini и клетки его ядра; snS - substantia nigra Sommeringii; ccr - основание (basis) ножки мозга; flp - задний прямой пучок покрова; lm - петля; qg - серое вещество переднего возвышения четверохолмия: qa - его белое вещество; S - сильвиев водопровод
Бехтерев (Arch. f. Anat. u. Physiol., 1S97) различает еще несколько прибавочных ядер, расположенных в окрестности главного.
Ядра зрительного нерва, II пара. Ядра этого нерва лежат так близко к поверхности, что о внутримозговых корешках нерва, подобных, например, корешкам n. oculomotorii, пронизывающих мозг, говорить не приходится. Волокна же, составляющие зрительный нерв и зрительный тракт, как у всех чувствующих нервов, начинаются от скопления клеток, лежащих вне мозга, именно от ганглиозных клеток сетчатой оболочки глаза. Затем, выходя через отверстие в наружных оболочках глазного яблока, образуют зрительный нерв, который вступает в перекрест зрительных нервов (chiasma nervorum opticorum). Перекрест, однако, неполный у человека и животных, глаза которых имеют одно общее поле зрения. Часть волокон каждого глаза остается на той стороне, от глаза которой она произошла; другая часть переходит в хиазме на противоположную сторону. За хиазмой начинаются те части зрительных нервов, которые носят название зрительных трактов. Каждый из этих трактов слагается из волокон, пришедших из обоих глазных яблок, таким образом, что в нем заключается пучок волокон, соответствующий наружной половине сетчатки паза своей стороны, и другой пучок, соответствующий внутренней половине сетчатки глаза противоположной стороны (эти части сетчаток идентичны, т. е. на них надают изображения одного и того же предмета). Волокна зрительных трактов направляются затем к мозговым (вторичным) ядрам. Но в составе их имеются, кроме волокон, происходящих от сетчаток, еще волокна образующие так наз. спайку Gudden. Это пучок волокон, соединяющий мозговые ядра зрительного нерва между собой. Спайка лежит частью на заднем краю, частью на поверхности хиазмы.
У животных, глаза которых помещаются на сторонах головы и потому не имеют общего поля зрения, перекрест волокон в хиазме полный.
Ядра зрительного нерва многочисленны, но общепризнаны только три. Эти последние видны на поверхности мозгового ствола невооруженным глазом и описываются в макроскопической анатомии: 1) pulvinar зрительного бугра, 2) наружное коленчатое тело и 3) переднее возвышение четверохолмия. Из них зрительный бугор и наружное коленчатое тело считаются собственно зрительными центрами, а четверохолмие рассматривается как интернодий в рефлекторном пути к мышцам глазного яблока. Даркшевич таким же рефлекторным интернодием считает еще шишковидную железу, которая соединена с зрительным трактом посредством pedunculi conarii. Вторым звеном в цепи зрительных проводников являются волокна, которые начинаются от клеток указанных ядер и направляются в составе белого вещества полушария к тому участку коры, который считается корковым, психосензорным, зрительным центром. Этот участок лежит на внутренней и частью на наружной стороне затылочной доли полушария (рис. 111 и 112, 16). Путь волокон второго звена вполне не известен; но один пучок, заложенный в centr. semi ovale и идущий от зрительного бугра к затылочной доле (так наз. пучок Вернике), признается за такой путь.
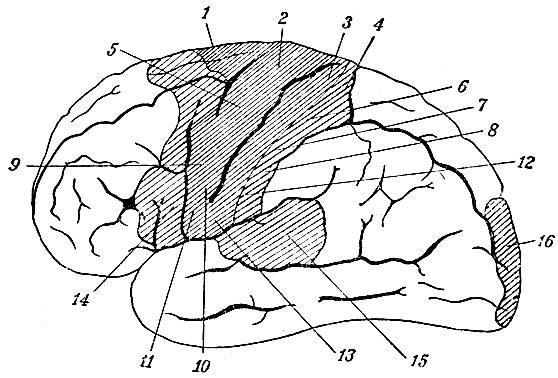
Рис. 111. Психомоторные и психосензорные центры в коре наружной поверхности полушария мозга человека. 1 - туловище; 2 - бедро; 3 - голень; 4 - стопа; 5 - плечо; 6 - локоть; 7 - кисть; 8 - пальцы руки; 9 - голова и глазное яблоко; 10 - мышцы лица; 11 - мышцы рта; 12 - подкожная мышца шеи (platysma); 13 - жевательные мышцы; 14 - язык и гортань; 15 - центр слуха; 16 - часть зрительного центра
Ядра n. olfactorii (I пара). Волокна так называемой fila olfactoria, которые соответствуют периферическим частям других нервов и соединяют обонятельную луковицу мозга (и у хищных животных обонятельную долю мозга) со слизистой оболочкой носа, начинаются как отростки от клеток, заложенных в эпителии слизистой оболочки носа, и, войдя в полость черепа, оканчиваются около клеток луковицы. Здесь возникают волокна второго звена цепи, которые образуют так называемый tractus olfactorius и направляются к корковым центрам. Сколько этих центров в коре мозга и где они находятся, в этом отношении указания авторов крайне разнообразны. На одной области, однако, все авторы согласны - это передний конец gyri uncinati (s. gyrus hyppocampi) височной доли мозга (рис. 112, 17). Кроме этих мозговых путей, tracms olfactorius содержит еще пучок волокон, который соединяет правую и левую обонятельные луковицы в виде спайки. Пучок этот переходит с одной стороны на другую через переднюю белую спайку мозга (comissura alba anterior).
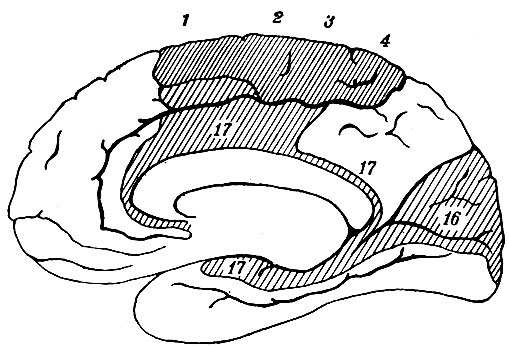
Рис. 112. Психомоторные и психосензорные центры в коре внутренней поверхности полушария. 1, 2, 3, 4, - туловище и нижняя конечность (продолжение центров наружной поверхности полушария); 16 - зрительный центр; 17, 17, 17 - обонятельный центр
Обонятельный нерв есть единственный - на пути между концевым аппаратом и мозгом - не перекрещенный.
Центры мозговой коры. Прежде всего следует установить точнее понятие о границах этих центров. Центры эти не следует представлять себе как строго отграниченные от соседних подобных центров: по выражению Оберштейнера, это только области, которые имеют наибольшее отношение к данной функции, и степень этого отношения постепенно уменьшается по мере удаления от данного центра, а края соседних центров как бы покрывают друг друга.
Представленная первоначально (Ferier) схема кортикальных центров была проверена Exner и другими на основании многочисленных собранных ими патологоанатомических наблюдений над мозгом человека. С некоторыми изменениями она подтвердилась. Так, Флексиг дает следующее разделение поверхности мозга человека.
Вся кора полушария, исключая insula Reilii, распадается на следующие области: па наружной поверхности полушария по обе стороны fiss. Rolandi помещается ряд двигательных центров (психомоторные центры), из которых верхние имеют отношение к туловищу и нижней конечности (рис. 111), средние - к верхней конечности, нижние - к лицу, языку и шее. В этой же области, но занимая более широкое место, помещаются так называемые психосензорные центры, имеющие отношение к чувству осязания. Область эта впереди захватывает начала всех трех лобных извилин, а сзади - всю ширину задней центральной извилины. Этот пояс коры (содержащий двигательные и чувствующие центры) распространяется также на внутреннюю поверхность полушария, занимая там всю парацентральную дольку и заднюю часть первой лобной извилины (рис. 112).
Кроме указанной области, отмечены как психосензорные центры еще следующие участки коры: задняя часть первой височной извилины - как центр чувства слуха: gyrus fornicatus в полосе, прилежащей к мозолистому телу, а также внутренний край gyri hyppocampi - как центр обоняния; наконец, cuneus (рис. 112), а также часть наружной поверхности затылочной доли (рис. 111) признаны зрительным центром.
Лобной доле, частям затылочной и височной долей, поскольку последние не заняты двигательно-чувствующим (теменным) поясом, Флексиг приписывает чисто психическую функцию.
Значение центральной дольки (insula Reilli s. lobus opertus) определить в настоящее время нельзя.
Кора мозжечка, по мнению Флексига, должна играть важную роль в сочетании функции всех долей большого мозга, так как она соединена при помощи массы волокон (волокна его ножек к мозгу и варолиеву мосту) со всеми областями коры большого мозга и сама обладает очень большим количеством ассоциирующих и коммиссуральных волокон. Что касается узлов полушарий, т. е. зрительного бугра и полосатого тела (corp. caudat. и nucl. lenticularis), то их значение очень темно. Наблюдение случаев недоразвития коры лобной доли мозга, причем обыкновенно недоразвиты и зрительные бугры, с одной стороны, с другой - совпадение недоразвития мозжечка с уменьшением полосатого тела позволяют только догадываться о сродстве функций одних узлов (зрительных бугров) с корой полушарий, других (полосатого тела) - с корой мозжечка.
При использовании материалов сайта активная ссылка обязательна:
http://anfiz.ru/ 'Анатомия и физиология человека'