Глава XXXII - 1936
В 1936 г. павловская школа разрабатывала ряд вопросов, в которых непосредственное участие принимал Иван Петрович. Некоторые вопросы были белее или менее закончены.
К числу таких относится вопрос об условном рефлексе как ассоциации. Можно ли образовать условную, временную связь между двумя одновременно применяющимися индиферентными раздражителями, т. е. образовать у собак ассоциацию? Положительный ответ на этот вопрос был дан в опытах Н. А. Подкопаева и И. О. Нарбутовича (1, 2). Эксперименты были проведены на 3 собаках. У каждой из них в течение нескольких опытов применяли свет электрической лампочки - 5 сек. и сразу же после нее тон - 5 сек., после этого свет один стал подкрепляться едой. Тогда и тон, ни разу не совпадавший с едой, стал сразу же вызывать условный пищевой рефлекс. У двух собак производили последовательное сочетание: шум - 5 сек. - вертушка 5 сек., без каких-либо подкреплений и без других условных раздражителей. Когда же вертушку превратили путем подкрепления электрическим током в условный оборонительный раздражитель, то и шум стал вызывать условный оборонительно-двигательный рефлекс. Таким образом достаточно было 20-30 сочетаний двух индиферентных раздражителей, чтобы потом можно было обнаружить образовавшуюся между ними связь путем превращения одного из них в пищевой или оборонительный. Наличие временной связи между ними можно было видеть и до сопровождения одного из них безусловным раздражением. Она выражалась в двигательных реакциях, а именно - при свете наблюдался поворот головы в сторону тона, при тоне - поворот в сторону лампочки. Следовательно, образование временной связи между двумя индиферентными (вызывающими лишь обычный ориентировочный рефлекс)* раздражителями в коре больших полушарий собаки вполне возможно. Авторы рассматривали это, как "ассоциацию по одновременности". Эта временная связь является двусторонней. Непременным условием ее образования должно быть непрерывное поддержание ориентировочной реакции на каждый из двух сочетаемых раздражителей. Раз образованная такая ассоциативная связь может держаться в течение многих месяцев и даже лег. Аналогичные опыты с положительным результатом были сделаны д-ром Панферовым (в лаборатории проф. Красногорского) на детях, проф. Ивановым-Смоленским тоже на детях, д-ром Шохором (в лаборатории проф. Зеленого) на взрослых людях. Такие же опыты, проводившиеся в лаборатории проф. Зеленого на собаках, дали отрицательный результат. Это зависело от того, что было допущено угасание ориентировочного рефлекса на применявшиеся раздражители.
* (По существу, авторы не имели дела с "индиферентными" раздражителями, поскольку последние вызывали ориентировочную реакцию.)
С самого начала работы по условным рефлексам существовала идея о том, что условный рефлекс и ассоциация - одно и то же. Красной линией эта идея прошла через весь ход исследований павловской школы. Верна она или нет? Вопрос необходимо конкретизировать и уточнить. По этому поводу Иван Петрович говорил, что "наш условный рефлекс - частный случай ассоциации, при которой один раздражитель (условный) слабее другого (безусловного)". Общий же закон условных рефлексов таков: "То, что совпадает во времени, то и образует условную связь, при этом порядок имеет значение только в зависимости от силовых отношений двух раздражителей"*. Понятие ассоциации шире, чем понятие условного рефлекса. Но понятие ассоциации совпадает с понятием корковой временной связи. И в основе условного рефлекса и в основе ассоциации лежит общий нервный механизм коркового замыкания, замыкания нервного возбуждения между двумя элементами коры. Некоторые считали, что ассоциация имеет субъективное содержание, а условный рефлекс его не имеет. Условный рефлекс есть понятие физиологическое, и в него входит не только понятие внешней, объективной реакции, но и понятие о внутренней сущности этой реакции - ее нервном механизме. Осуществление этого нервного механизма у человека может быть связано с теми или иными субъективными переживаниями, сознаваемыми или несознаваемыми. С другой стороны, не существует в природе никаких ассоциаций, только как субъективно-психологических содержаний: они имеют тот же корковый нервный механизм.
* ("Среда", 14 октября 1931 г.)
В чем же разница между условным рефлексом и ассоциацией? При условном рефлексе обязательно участие подкорки, при ассоциации оно не обязательно. Специфической особенностью коры больших полушарий человека является ее способность производить всевозможные ассоциации, корковые временные связи без непосредственного их подкрепления со стороны какой-либо безусловной, врожденной деятельности. Это возможно благодаря более высокой реактивности и лабильности высшего отдела головного мозга человека по сравнению с животными. Ассоциации, образованные у собак Подкопаевым и Нарбутовичем, нуждались для своего существования в наличии ориентировочного рефлекса на сочетаемые раздражители, т. е. в наличии врожденной деятельности, повышавшей тонус коры. Поэтому эти ассоциации не тождественны с человеческими ассоциациями, а представляют только переход к ним в процессе эволюции.
Итак, с нашей точки зрения условный рефлекс и ассоциация суть формы корковой временной связи и имеют общий нервный механизм коркового замыкания. Отличие ассоциации от условного рефлекса то, что она является такой корковой временной связью, которая для своего осуществления не требует обязательного участия подкорки, как в случае условного рефлекса.
Иван Петрович придавал большое значение этому вопросу и одно время собирался устроить на своей "среде" дискуссию в связи с главой об ассоциации, в последней книге Келера "Психологические проблемы"*.
* (W. Kohler. Psychologische Probleme.)
Переходим к другому основному вопросу - вопросу о "законе силы". Как известно, этот закон был разработан на условных положительных раздражителях и поэтому распространялся только на положительные условные рефлексы. А имеет ли он значение для другой половины условных рефлексов - тормозных? Этот вопрос был поставлен и разрешен в работе Ф. П. Майорова (3), исследовавшего в течение нескольких лет зависимость силы тормозного процесса от физической интенсивности тормозного раздражителя*. "У нас, - говорил Павлов, - торможение не исследовалось так, как возбуждение, нет полной симметрии в исследованиях того и другого"**. Указанная работа во многих отношениях заполняла существующий пробел. Опыты велись в разных вариациях на 5 животных по обычной, секреторной пищевой методике. Сначала были выработаны диференцировки различной физической силы, например: диференцировки на метроном 50 и 200 к положительному метроному 100, или диференцировки на метрономы 60 и 200 к положительному метроному 120. И физически слабая диференцировка (М. 50 и М. 60) и физически сильная (М. 200) были абсолютными и давали прочный нулевой эффект, не сказываясь на остальных условных рефлексах в стереотипной системе с 5-минутным интервалом. Испытания на концентрацию торможения обнаружили существенную разницу между обеими диференцировками. Обе диференцировки были достаточно концентрированными и выявить разницу между ними можно было лишь на очень коротких интервалах в несколько секунд. Получился такой результат: диференцировка на М. 200 всегда вызывала после себя положительную индукцию, иногда значительных размеров, диференцировка же на М. 60 положительной индукции не давала, а давала последовательное торможение. Следовательно, при действии М. 200 в коре развивался более концентрированный тормозный процесс, чем при М. 60, а более концентрированное торможение всегда более сильное. Значит, физически более сильный тормозный раздражитель вызывал и физиологически более сильный тормозной процесс. Это доказывалось и другими опытами. Таким образом закон силы должен быть распространен и на действие условных тормозных раздражителей. Далее, были проведены испытания на предел напряжения диференцировочного торможения в обоих случаях. Продление обеих диференцировок до 8 мин. вместо обычных 20 сек. также показало различие между ними. Предел напряжения торможения в случае М. 200 оказался ниже, чем в случае М. 60, в первом случае скорее наступало растормаживание и симптомы двигательного возбуждения, чем во втором, т. е. диференцировка на М. 200 была более трудной и напряженной, чем М. 60. У разных по силе нервных систем предел напряжения торможения был не одинаков. В дальнейшем были сделаны три серии опытов с растормаживанием сильной и слабой диференцировок: 1) растормаживание при помощи ориентировочных раздражителей, 2) растормаживание посредством кофеина и 3) превращение обеих диференцировок в условные положительные раздражители путем их пищевого подкрепления (т. е. односторонняя переделка). Оказалось, что при всех трех испытаниях легче и скорее растормаживалась как раз более сильная диференцировка (М. 200), а не более слабая (М. 60). В случае переделки в положительный раздражитель скорее превратилась более сильная диференцировка и за одинаковое число сочетаний дала больший секреторный эффект, чем слабая. Как же понять этот факт, что сильная диференцировка, связанная с более сильным, концентрированным торможением, растормаживается, однако легче и быстрее? Понять это можно, исходя из признания постоянного взаимодействия возбуждения и торможения и их внутреннего единства. При выработке диференцировки на физически сильный раздражитель торможению приходится преодолевать более сильный раздражительный процесс, что определенным образом, как и показали данные опыты, отражается на кривых образования обеих диференцировок. В первом случае диференцировка образуется труднее и выработанное торможение имеет большее напряжение. Поэтому сильное, диференцировочное торможение легче и скорее растормаживается, т. е. обнаруживает скрытое в нем возбуждение. Таким образом эти данные подчеркивали единство возбуждения и торможения.
* (Итоги работы были доложены еще в 1934 г. на заседании Ленинградского общества физиологов.)
** ("Среда", 27 марта 1935 г.)
Мы считаем, что это положение должно быть основным в нашей работе и определять генеральную линию исследований высшей нервной деятельности. Проблема возбуждения и торможения была "проклятым вопросом", по выражению Павлова. Она не была и не могла быть разрешена одним бесчисленным эмпирическим накоплением фактов. Для ее правильного разрешения необходимо было исходить из какой-то идеи, ибо, как говорил сам Павлов, "если нет в голове идеи, то и не видишь фактов". Такой идеей должна быть идея о взаимодействии и единстве возбуждения и торможения, как противоположных нервных процессов. Это положение указывает путь для экспериментального разрешения проблемы.
В опытах Майорова с кофеином демонстративно выступили индукционные отношения между корой и подкоркой. Применение кофеина увеличивало условные рефлексы и уменьшало безусловные, что является результатом действия отрицательной индукции с коры на подкорку.
Идея взаимодействия возбуждения и торможения получила свое развитие в двух следующих работах Ф. П. Майорова. Была поставлена задача проследить это взаимодействие при развитии запредельного торможения (4). С этой целью был выработан среди прочих условный пищевой рефлекс на тон катодного генератора. Затем постепенно увеличивалась интенсивность этого тона, но так, что на каждую интенсивность приходилось по 10 сочетаний. По мере усиления тона все более и более рос рефлекс на него, но до определенного предела, за которым рефлекс стал падать в результате вмешательства запредельного торможения. Чем более увеличивалась далее сила звука, тем более уменьшался рефлекс. Грубо говоря, запредельное торможение как бы отмеривалось по физической интенсивности звука: увеличивалась сила звука - увеличивалось и запредельное торможение. Таким образом была прослежена динамика развития запредельного торможения и его дальнейшая судьба по мере прироста силы раздражителя. В этой же работе было установлено, что чистые тона вообще и особенно слабых степеней интенсивности осложняются торможением. Не есть ли это тоже самое запредельное торможение, развивающееся в результате "задалбливания" корковых нервных клеток, к которым адресуется чистый тон? Надо думать, что чистый тон в отличие от музыкального вызывает возбуждение меньшего числа корковых клеток; следовательно, вся интенсивность звукового раздражения ложится на меньший участок мозговой коры и оказывается относительно более сильней. В этом случае эффект по "закону силы" будет зависеть не только от силы условного раздражителя и состояния корковых клеток, но и от узости пространственного диапазона возбуждаемых корковых элементов. Чистый и музыкальный тон одной и той же силы должен иметь неодинаковое действие на кору: в первом случае сила раздражения относится к меньшему числу корковых клеток, скорее может вызвать их утомление и развитие "охранительного" запредельного торможения, снижающего условно-рефлекторный эффект. Эта гипотеза имеет под собой некоторые данные и других работ.
Далее был поставлен вопрос (5): как повлияет диференцировочное торможение в той же группе корковых клеток на запредельное торможение в них. Не уменьшится ли последнее в силу того, что диференцировочное торможение будет способствовать восстановлению работоспособности данных клеток и как пойдет в этом случае выработка самой диференцировки? С этой целью вырабатывалась диференцировка на тот же чистый тон, только меньшей интенсивности: была взята оптимальная интенсивность, дававшая ранее максимальный эффект без запредельного торможения. Эта диференцировка образовалась не быстрее другой обычной диференцировки на метроном. Таким образом наличие запредельного торможения не ускорило выработки диференцировки. По мере укрепления диференцировки на более слабый тон увеличился рефлекс на запредельный тон. Как будто ответ на поставленный вопрос получился положительный. Однако он объяснялся развитием положительной индукции с диференцировочного тона на запредельный, восстанавливающее же значение диференцировочного торможения было от нас скрыто. В дальнейшем при расположении обоих тонов рядом между ними упрочились взаимно-индукционные отношения. Таким в этих опытах оказалось взаимодействие запредельного и диференцировочного торможения.
Несколько раз мы касались вопроса о градации гипнотических фаз. Снова возвращаемся к этому вопросу в связи с работой И. С. Розенталя (6) о стадиях при развитии торможения в коре больших полушарий. Он многократно повторял положительные условные рефлексы в течение опыта при отставлении раздражителей на 60 сек. В результате длительного применения таких опытов стало развиваться общее торможение. При развитии тормозного состояния наблюдался следующий порядок в развитии стадий:
1) нормальная,
2) переходная к уравнительной,
3) уравнительная (на различных уровнях),
4) переходная к наркотической,
5) наркотическая,
6) парадоксальная,
7) тормозящая.
Автор отметил большой ряд вариаций этих стадий. От себя добавим следующее: 1) наркотическая фаза может часто следовать после парадоксальной, 2) в намеченном порядке в разных местах может возникать ультрапарадоксальная фаза. Ультрапарадоксальные отношения мы встречаем в виде ультрапарадоксальной гипнотической фазы и независимо от развития гипнотических состояний. И в том и другом случае сущность одна - извращение взаимной индукции. Гипнотическое торможение, действуя на положительный очаг и вследствие этого растормаживая тормозной очаг ассоциированной пары раздражителей, может вызвать извращение взаимной индукции и появление ультра-парадоксальной фазы.
К учению о гипнотических фазах следует сделать еще одно важное замечание. По шаблону было принято говорить и писать о гипнотических фазах даже и тогда, когда нет объективных признаков развития гипнотического торможения, но когда искажаются нормальные силовые отношения между условными рефлексами, например: "гипнотические фазы" в случаях повышенной возбудимости, когда нет еще никакого запредельного торможения, а наблюдается повышение всех условных рефлексов (уравнительная фаза на высоком уровне и т. п.). Здесь вводится явное смешение видимости и сущности. Видимость одна, а сущность разная. Во избежание дальнейшей путаницы мы предлагаем ввести строгое различение двух понятий: "гипнотических фаз" (или "переходных состояний") и "фазовых изменений". Гипнотические фазы суть частный случай фазовых изменений коры. Фазовые же изменения охватывают все вариации общего тонуса коры как в сторону развития общего торможения, так и в сторону развития повышенного возбуждения. С этой точки зрения уравнительная фаза на высоком уровне вовсе не есть гипнотическая фаза, а случай фазовых изменений коры в широком смысле слова.
Ультрапарадоксальной фазе были посвящены две работы (Линдберга и Петровой). А. А. Линдберг (7) выработал дифередцировку на касалку, затем вызывал торможение положительной касалки путем угашения и в результате наблюдал отношения, близкие к ультрапарадоксальным, так как тормозная касалка приобретала значительное положительное действие. Вызывать таким путем ультрапарадоксальные отношения становилось труднее и труднее по мере укрепления диференцировки и упрочения индукционных отношений между положительным и тормозным раздражителями. Бромистый натр также ослаблял ультрапарадоксальные отношения.
Упомянутые ранее опыты М. К. Петровой (8) с ультрапарадоксальной фазой привели Павлова к заключению, что в данном случае у (положительного и тормозного метрономов) "достаточных оснований для пространственного разделения процессов возбуждения и торможения не имеется, что можно считать их существующими в одном и том же корковом пункте" (300) (8).
На ряду с идеями о высокой пластичности, тренировке, приспособляемости и работоспособности коры больших полушарий существовали идеи об "изнашиваемости" и "инвалидности" корковых клеток. Одной из работ на последнюю тему была работа И. Р. Пророкова (9) об "изнашиваемости" нервных клеток. С течением времени от повторения одних и тех же условных рефлексов наблюдалось постепенное их падение. Переход на новые раздражители вызывал повышение условных рефлексов. Подобный факт был установлен ранее Н. А. Подкопаевым (см. главу XXII) и встречался у других исследователей. Мы имеем здесь дело с утомляемостью корковых элементов под влиянием повторения однообразных раздражителей. Замена одних корковых элементов другими устраняет указанное явление. Клиницистам известно, что при психотерапии перемена жизненной обстановки часто имеет решающее значение для выздоровления.
Но стоит ли говорить об "изнашиваемости" нервных клеток там, где мы находим явления коркового утомления? По нашему мнению, до "изнашиваемости" здесь далеко и еще дальше до "инвалидности", о которой принято было говорить довольно часто. В связи с последним иногда имело также место злоупотребление излишним вмешательством запредельного торможения: некоторые старались видеть его там, где его вовсе не было.
В 1936 г. был разрешен вопрос о механизме образования условного рефлекса при неподкреплении первых трех сочетаний и подкреплении четвертого (опыты Выржиковского, Усиевича, Петровой, Яковлевой - см. главу XXVIII). В. К. Федоров (10) применял условное кожно-механическое раздражение три раза, подкрепляя только третье сочетание. Такую задачу его собака решила, т. е. первые два раздражения давали тормозный эффект, а третье - положительный. При этом оказалось следующее: если в начале опыта применялись все три касалки, то все они были положительными; если они применялись в конце опыта, то все они оказывались тормозными. Таким образом образовывалась временная связь первых двух тормозных касалок и третьей положительной касалки с состоянием коры больших полушарий в целом на разных стадиях ее деятельности*. Намек на такое решение вопроса был в работе В. В. Яковлевой (см. главу XXXI).
* ("Среда", 15 января 1936 г.)
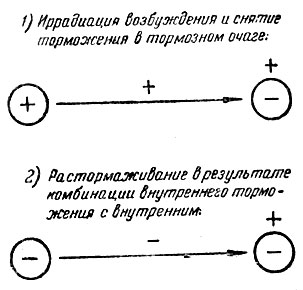
Рис. 6. Механизмы растормаживания
Новые опыты К. С. Абуладзе (11) дополнили имевшиеся данные о механизме генерализации. В первой половине опытов у него были пищевые условные рефлексы, во второй - кислотные. Экстрараздражители в первой половине опыта давали пищевой эффект, во второй - кислотный эффект. Таким образом физиологической основой генерализации являются иррациация и суммация.
Подытожим данные, касающиеся механизма растормаживания. Все явления растормаживания можно свести к двум группам:
1) иррадиация возбуждения и снятие торможения в тормозном очаге;
2) комбинация внутреннего торможения с внутренним же дает растормаживание*.
* (См. главу XXVIII и дальнейшие.)
Во втором случае надо учесть значение следующих моментов: 1) значение интервала между раздражителями, вызывающими внутреннее торможение (длинный интервал может дать иррадиацию и суммционный эффект торможения, короткий интервал может быть связан с концентрацией торможения, положительной индукцией и растормаживанием второго очага торможения); 2) значение силы торможения в обоих тормозных очагах, их концентрированности (развивающаяся при этом положительная индукция может препятствовать иррадиации торможения и т. д.); 3) значение типа нервной системы; 4) возможны истинные случаи торможения торможения, дающие, растормаживание. Это отнюдь нас не возвращает к старой, отошедшей в прошлое, концепции растормаживания по типу внешнего торможения.
Итак, растормаживание вызывается 1) иррадиацией возбуждения, 2) положительной индукцией и 3) соединением одного торможения с другим торможением. Физиологический механизм первых двух случаев понятен. Но остается неясным механизм третьего случая. Мы имеем здесь переход торможения в возбуждение. Но как получается возбуждение из соединения двух торможений? Этот вопрос не может быть разрешен средствами одной физиологии высшей нервной деятельности.
Д. А. Каменский (12) наблюдал благотворное влияние кратковременного естественного сна на восстановление условно-рефлекторной деятельности. 25-30 мин. сна собаки перед опытом давали восстановление работоспособности коры до ее уровня в бодром состоянии.
Рассмотрим теперь несколько работ о типах. В. В. Яковлева (13) сделала тщательные исследования флегматического типа ("Золотистый"), подробно характеризовав его силу, уравновешенность и инертность. Только различная подвижность нервных процессов отличает флегматика от сангвиника. "Золотистый" имел также наклонность тормозить все условные рефлексы по мере повторения одной и той же системы раздражителей. Это было проявление инертности тормозного процесса. На этой собаке было проведено также исследование величины безусловных рефлексов в зависимости от силы и деятельности условных пищевых раздражителей. Полученные данные подтверждали прежние результаты.
В. П. Головина (14) провела сравнение выработки диференцировок у сангвиника и слабого типа. Вырабатывались в разное время три диференцировки: на метроном 60 к положительному М. 120, на касалку по месту, на круг к положительному квадрату. В процессе их образования у сангвиника возникло общее возбуждение, у слабого типа - защитное торможение в виде гипноза. Головиной, как мы уже упоминали раньше, удалось получить невроз у собаки уравновешенного сангвинического типа.
На потомстве одной собаки, хорошо ранее изученной, Е. З. Строгая (15) сделала попытку учесть изменчивость высшей нервной деятельности и построить вариационные ряды вариаций процессов возбуждения, торможения, последовательного торможения и т. д.
Н. А. Подкопаев (16) установил изменения подвижности нервных процессов у собак в старческом возрасте. Применив двустороннюю переделку положительного и тормозного метрономов, он показал, что при старении происходит сдвиг в сторону инертности, застойности. Эта инертность сказалась на обоих нервных процессах, но на тормозном в большей степени, чем на процессе возбуждения.
На основании как этих исследований, так и других можно считать установленным, что старение связано 1) с понижением реактивности коры больших полушарий, 2) с уменьшением подвижности нервных процессов и 3) с ослаблением функций коркового торможения.
У собаки сильного типа В. К. Федоров (17) описал упорную тормозную реакцию на всякие новые изменения в опытах. Автор ставил это в связь с "чрезмерно задержавшимся рефлексом осторожности". В физиологической трактовке примеров из жизни людей автор допустил несомненное упрощение, что является следствием увлечения методом аналогий.
Две работы А. А. Линдберга (18, 19) о броме дополняли имевшийся материал, не внося ничего принципиально нового. Изучалось влияние бромистого натрия на развитие запаздывания пои обычном оставлении условного раздражителя на 30 сек. Производилась регистрация условной секреции по 5 сек: Под влиянием брома запаздывание усиливалось. Это подтверждало положение, что бром усиливает торможение.
На другой собаке (19) Линдберг применял хроническое бромирование разными дозами NaBr: от 0,75 до 3,0. При длительном бромировании бром утрачивал свое терапевтическое действие. Он снова приобретал полное терапевтическое действие после прекращения бромирования и усиленного вымывания брома из организма при помощи жидкостей и вытеснения брома посредством хлора физиологического раствора. При неоднократном повторении длительных периодов бромирования экспериментатор наблюдал постепенное увеличение устойчивости нервной системы к брому.
Тот же автор описал интересный случай "изолированного поражения тормозного процесса в ограниченном пункте коры" (20). Опыты производились на собаке, у которой раньше в течение долгого времени практиковалась диференцировка на метроном 200 к положительному М. 76 и делались много раз сшибки этих раздражителей. Линдберг пытался выработать диференцировку на М. 50 к положительному М. 100. Такая диференцировка не вырабатывалась. Однако без особого труда можно было образовать диференцировку на кожно-механическое раздражение по месту и на тихий шум (к положительному сильному шуму). Также можно было получить диференцировку на заглушённый М. 100 к незаглушенному М. 100. На основании этих данных, автор считал возможным сделать вывод о том, что "метрономный район являлся изолированным болезненным районом" и что на протяжении 7 мес. в этом "пункте" отмечалось исключительное поражение тормозного процесса. Почему же тогда можно было получить тонкую диференцировку на интенсивность одной и той же частоты ударов метронома (М. 100)? Повидимому, дело объясняется проще: трудность выработки, диференцировки на М. 50 обусловливалась тем, что до этого в течение долгого времени близкая частота М. 76 была укреплена как положительная.
С. В. Клещов продолжал свои исследования элементов музыкального слуха. На двух собаках он провел опыты со сравнением действия одного тона с двойным и четверным. Было доказано, что увеличение числа тонов в условном раздражителе обусловливает больший эффект слюноотделения. Это можно видеть на примере суммарного эффекта от четырех условных раздражителей:
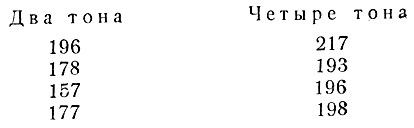
Каждый тон длился 5 сек. и следовал за другим без перерыва при общем отставлении в 20 сек. Таким образом увеличение музыкальных элементов увеличивает условно-рефлекторный эффект. Отсюда делается понятным тонизирующее действие музыкальной орнаментики.
Как несомненно прогрессивный момент в общем развитии учения с высшей нервной деятельности мы должны отметить данную П. С. Купаловым (21) критику чисто количественной точки зрения. Мы уже говорили о том, что категория качества, хотя и не игнорировалась, но не нашла надлежащего ей места в учении Павлова и работах его сотрудников. Купалов подчеркивал качественные, специфические особенности корковой деятельности в отличие от низшей нервной деятельности. Павлов открыл не только общие у коры с центральной нервной системой закономерности, но и специфические, особенные закономерности корковой деятельности. Купалов был вполне прав утверждая, что кора больших полушарий животных есть орган "принципиально новых нервных деятельностей".
В эти годы не прекращалась дальнейшая разработка проблемы корковой локализации. С. И. Лебединская и И. С. Розенталь получили бесполушарную собаку и пробовали образовать у нее условные рефлексы. Это им не удалось. Надо думать, что если произведено полное удаление обоих больших полушарий, то такие животные не способны к образованию каких-либо условных рефлексов. Подтверждение павловской теории корковой локализации было получено Розенталем (22) в его опытах на собаках или без двигательного или без кожного анализаторов. У двух животных был удален двигательный анализатор, и у двух - кожный. После экстирпации вырабатывался условный двигательный пищевой рефлекс подачи лапы на словесный приказ "правую". Условный рефлекс был выработан у всех собак, несмотря на отсутствие соответствующих корковых центров. Автор считал, что образование рефлекса произошло благодаря существованию рассеянных корковых элементов, согласно учению Павлова.
К работе Конорского и Миллера, вышедшей в 1936 г., Павлов написал предисловие "Физиологический механизм так называемых произвольных движений" (23). На основании опытов Красногорского (1911 г.), Иван Петрович считал доказанным положение, что моторная корковая зона есть анализатор. Но в настоящее время надо полагать, что анализаторную функцию этой зоны выполняют ее верхние слои, а нижние имеют эффекторную функцию. Далее Иван Петрович полагал, что в коре больших полушарий существует двусторонняя проводимость возбуждения, что коренным образом отличает кору от спинного мозга, где имеется только односторонняя проводимость. Благодаря двусторонней проводимости осуществляются произвольные движения. Конкретное физиологическое основание произвольности движений он видел в связи кинэстетических клеток коры со всеми остальными клетками коры. Какая же это связь: врожденная или приобретенная? Иван Петрович предполагал более вероятным второе, т. е. что эти корковые связи образуются в течение индивидуальной жизни. Экспериментальный анализ механизма произвольных движений - это большой вопрос современной физиологии, требующий дальнейшего исследования.
Опыты Ю. М. Конорского и С. М. Миллера (24), начатые в Варшаве и законченные в Ленинграде, принадлежат к наиболее сложным опытам, полное физиологическое объяснение которых еще не найдено. На нескольких собаках они сделали следующие эксперименты:
1. Пассивное сгибание ноги (при помощи блока) сопровождали едой. При наличии пищевого возбуждения собака производила сгибание этой ноги. Далее, они производили пищевое подкрепление комбинации какого-либо условного раздражителя и пассивного сгибания йоги. При изолированном действии условного раздражителя сгибания не происходило, как это имело место в опытах Скипина и Петровой (см. главу XXVI). Сгибание получалось только после предварительного затормаживания условного раздражителя посредством его угащения.
2. Производилось сочетание пассивного сгибания ноги с вливанием раствора соляной кислоты. Сигнал вливания кислоты вызывал активное разгибание ноги.
3. Вырабатывался условный пищевой тормоз: одно сгибание сопровождалось едой, условный раздражитель плюс сгибание не подкреплялись и были условным тормозом. В этом случае применение одного условного раздражителя вызывало активное разгибание.
4. Вырабатывался условный кислотный тормоз: одно сгибание подкреплялось кислотой, комбинация условного раздражителя со сгибанием кислотой не подкреплялась. В этом случае действие условного раздражителя сопровождалось активным сгибанием. Для того, чтобы лучше представить себе изложенные факты, изобразим их схематически:

На основании этих фактов авторы формулировали "особый закон образования двигательных условных рефлексов": "рефлекс образуется на "обстановку", рефлекс проявляется только при частичном торможении условного раздражителя, сочетаемого с пассивным движением" (275, 278) (24). По этому поводу надо сказать следующее: во-первых, всякий условный рефлекс, а не только двигательный, образуется на "обстановку", в состав которой условный агент входит, как основной и решающий элемент; во-вторых, для обнаружения рефлекса частичное торможение условного раздражителя необязательно, как показали опыты Петровой и Скипина. В этом случае, повидимому, дело сводилось к различным силовым отношениям между компонентами сложного раздражителя (из какого-либо агента плюс пассивное сгибание). В опытах Конорского и Миллера условный агент, как раздражитель, мог быть сильнее и вызывать отрицательную индукцию на действие более слабого пассивного сгибания. Поэтому эффект сгибания обнаруживается после ослабления влияния отрицательной индукции путем частичного угашения условного агента, кроме того, здесь могла еще сказаться и положительная индукция от угашенного раздражителя.
Данное Павловым в его предисловии объяснение описанных фактов не представляет объяснения их механизмов, а имеет только обще-физиологический, биологический характер.
Из четырех представленных случаев нам понятен лишь первый - активное сгибание при появлении пищевого возбуждения. Такой факт наблюдался и другими, например, Майоровым (см. главу XXVII). Три остальных случая менее понятны. Объяснения, данные Конорским и Миллером, не вполне удовлетворительны. Детальный анализ этих явлений с точки зрения их нервного механизма еще лежит впереди. Работой Конорского и Миллера не заканчивалась линия исследований павловской школы по физиологическому анализу произвольных движений. На этом пути возник ряд конкретных вопросов, еще ожидающих своих исследователей.
Павловская школа охватила своими экспериментами не только собак, но также обезьян низших и высших и человека. С 1929 г. по 1936 г. в Сухуми рядом лиц были проведены различные опыты по изучению поведения обезьян посредством метода условных рефлексов. Для этого применяли различные варианты методик, учитывая биологические особенности обезьян. Из работ на низших (собакоголовых) обезьянах отметим следующие основные результаты. Условные двигательные рефлексы большей частью образуются у них быстро и легко. То же можно сказать и относительно диференцировок и прочих тормозов. У обезьян очень развит ориентировочный рефлекс, часто растормаживающий тормоза и затормаживающий положительные рефлексы. Не трудно у них выработать условные следовые рефлексы. Низшие обезьяны не резко отличаются от собак по своей высшей нервной деятельности. Почти все, что было выработано у этих обезьян, можно было получить и у собак. Гораздо больший интерес представляют высшие, антропоидные обезьяны. Опыты на шимпанзе (взрослых и малышах) и оранг-утане касались как условных рефлексов, так и более сложного "разумного" поведения в той или иной ситуации, по типу известных опытов психолога В. Келера. Физиологи проводили экспериментальный анализ такого поведения и дали физиологическую расшифровку фактов Келера. Ими была отброшена всякая субъективно-психологическая и спиритуалистическая трактовка "разумного" поведения человекообразных обезьян. Ценный фактический материал был представлен по сравнительному изучению развития функций коркового торможения у высших и низших обезьян. В Сухуми была основана специальная лаборатория физиологии и патологии высшей нервной деятельности обезьян, где работа продолжается до сих пор.
Очень важные исследования на двух шимпанзе велись последние три года в Колтушах под непосредственным руководством Павлова. Павлов поставил перед собой задачу дать физиологическое решение проблемы интеллекта антропоидов. С этой целью эксперименты ставились так, чтобы можно было все учесть, проследить накопление жизненного опыта и его использование обезьяной при решении новых задач. Иван Петрович считал, что вся "разумность" высших обезьян сводится к образованию корковых временных связей, ассоциаций и их использованию в той или иной ситуации. Павловские эксперименты на антропоидах отличались от таких же экспериментов Келера, Иеркеса и других тем, что они были направлены на познание физиологических механизмов сложного поведения этих обезьян.
Эти исследования были продолжены Вацуро и Штодиным под руководством акад. Л. А. Орбели.
Работа по изучению корковой деятельности человека проводилась в двух тесно связанных между собою направлениях: клиническое наблюдение и эксперимент. Клиническое наблюдение патологических случаев в нервной и психиатрической павловских клиниках давало богатый фактический материал, который подвергался физиологическому анализу на основе учения о высшей нервной деятельности. Экспериментальное исследование проводилось в специальных физиологических лабораториях, организованных при обеих клиниках.
За 5 лет интенсивной работы павловские клиники внесли чрезвычайно ценный вклад как в физиологию и патологию высшей нервной деятельности, так и в практику клиники неврозов и психозов*.
* (Очерк работы нервной и психиатрической клиник см. в "Отчете ВИЭМ за II пятилетку". Изд. ВИЭМ, 1939, стр. 199. (Статьи проф. Ф. П. Майорова и проф А. Г. Иванова-Смоленского).)
Со смертью Ивана Петровича Павлова оборвалась драгоценная нить его личной творческой жизни, но не умерла павловская школа. Она продолжает жить и работать. Свидетельство этому - ежегодные павловские совещания физиологов - его учеников и последователей, работе которых мы посвятили несколько специальных статей*.
* (См. журнал "Под знаменем марксизма", 1939, № 7; 1940, № 7; 1941, № 4.)
Литература
1. Н. А. Подкопаев. Условный рефлекс как ассоциация. Материалы V Всесоюзного физиологического съезда, 1934, стр. 62.
2. И. С. Нарбутович и Н. А. Подкопаев. Условный рефлекс, как ассоциация. Труды физиологических лабораторий акад. И. П. Павлова, 1936, т. VI. вып. 2, стр. 5.
3. Ф. П. Майоров. О зависимости силы тормозного процесса от физической интенсивности тормозного раздражителя. Доклад в Ленинградском Обществе физиологов, 1934. Доклад на XV Международном физиологическом конгрессе 1935 г. Докторская диссертация 1936 г. Труды физиологических лабораторий акад. И. П. Павлова, 1938, т. VIII, стр. 62.
4. Ф. П. Майоров. О взаимодействии возбуждения и торможения при развитии запредельного торможения. Труды физиологических лабораторий акад. И. П. Павлова, 1940, т. IX, стр. 426.
5. Ф. П. Майоров. Взаимодействие запредельного и диференцировочного торможения. Там же, 1940, стр. 169.
6. И. С. Розенталь. Стадии при развитии торможения в больших полушариях собаки. Архив биологических наук, 1936, 41, 1.
7. А. А. Линдберг. Нарушение индукционных отношений между положительным и тормозным рефлексами и последствие этого у собаки возбудимого типа. Физиологический журнал СССР, 1936, т. XX, 6.
8. М. К. Петрова. Взгляд акад. И. П. Павлова на механизм, происхождения взрывчатости и ультрапарадоксальной фазы на основании последних фактических данных. Физиологический журнал СССР, 1938, т. XXIV, вып. 1-2.
9. И. Р. Пророков. Об "изнашиваемости" нервных клеток. Физиологический журнал СССР, 1938, т. XXIV, вып. 1-2.
10. В. К. Федоров. Опыты.
11. К. С. Абуладзе. Опыты.
12. Д. А. Каменский. Влияние кратковременного естественного сна на восстановление условно-рефлекторной деятельности головного мозга. Труды физиологических лабораторий акад. И. И. Павлова, 1940, т. IX, стр. 417.
13. В. В. Яковлева. Исследование высшей нервной деятельности собаки типа флегматика (сильный, уравновешенный, инертный). Труды физиологических лабораторий акад. И. П. Павлова, 1936, т. VI, 2.
14. В. П. Головина. Сравнение выработки диференцировок у собак различного типа нервной системы. Труды физиологических лабораторий акад. И. П. Павлова, 1938, т. VIII.
15. Е. З. Строгая. Изменчивость высшей нервной деятельности у собак. Там же.
16. Н. А. Подкопаев. К характеристике высшей нервной деятельности собаки в старческом возрасте. Физиологический журнал СССР, 1938, т. XXIV, вып. 1-2.
17. В. К. Федоров. Настойчивая тормозная реакция на новые изменения в окружающем у собаки сильного типа. Труды физиологических лабораторий акад. И. П. Павлова, 1936, т. VI, вып. 2, стр. 55.
18. А. А. Линдберг. О механизме действия брома. Физиологический журнал СССР, 1936, т. XX, вып. 5.
19. А. А. Линдберг. К фармакологии брома. Там же.
20. А. А. Линдберг. Случай изолированного поражения тормозного процесса в ограниченном пункте звукового анализатора мозговой коры. Физиологический журнал СССР, 1936, т. XX, 6.
21. П. С. Купалов. Внешняя среда и деятельность коры больших полушарий. Физиологический журнал СССР, 1938, т. XXIV, вып. 1-2.
22. И. С. Розенталь. Условный двигательный пищевой рефлекс у собак или без двигательного или без кожного анализатора. Физиологический журнал СССР, 1938, т. XXIV, вып. 1-2, стр. 345.
23. И. П. Павлов. Физиологический механизм так называемых произвольных движений (предисловие к работе д-ров Ю. М. Конорского и С. М. Миллера). Последние сообщения по физиологии высшей нервной деятельности. III. Труды физиологических лабораторий акад. И. П. Павлова, 1936, т. VI, 1. Двадцатилетний опыт, 1938.
24. Ю. М. Конорский и С. М. Миллер. Условные рефлексы двигательного анализатора. Труды физиологических лабораторий акад. И. П. Павлова, 1936, т. VI, I.
25. Л. А. Андреев. Характеристика слухового анализатора собаки на основании экспериментальных данных, полученных по методу условных рефлексов*. Журнал технической физики, 1936, т. VI, вып. 12.
* (Дано краткое изложение результатов работы автора по данному вопросу.)
При использовании материалов сайта активная ссылка обязательна:
http://anfiz.ru/ 'Анатомия и физиология человека'