Принципы координации в деятельности центральной нервной системы
В условиях физиологической нормы работа всех органов и систем тела является согласованной: на воздействия из внешней и внутренней среды организм реагирует как единое целое. Согласованное проявление отдельных рефлексов, обеспечивающих выполнение целостных рабочих актов, носит название координации.
Явления координации играют важную роль в деятельности двигательного аппарата. Координация таких двигательных актов, как ходьба или бег, обеспечивается взаимосвязанной работой нервных центров.
За счет координированной работы нервных центров осуществляется совершенное приспособление организма к условиям существования. Это происходит не только за счет деятельности двигательного аппарата, но и за счет изменений вегетативных функций организма (процессов дыхания, кровообращения, пищеварения, обмена веществ и т. д.).
Установлен ряд общих закономерностей - принципов координации: 1) принцип конвергенции; 2) принцип иррадиации возбуждения; 3) принцип реципрокности; 4) принцип последовательной смены возбуждения торможением и торможения возбуждением; 5) феномен "отдачи"; 6) цепные и ритмические рефлексы; 7) принцип общего конечного пути; 8) принцип обратной связи; 9) принцип доминанты.
Разберем некоторые из них.
Принцип конвергенции. Этот принцип установлен английским физиологом Шеррингтоном. Импульсы, приходящие в центральную нервную систему по различным афферентным волокнам, могут сходиться (конвергировать) к одним и тем же вставочным и эфферентным нейронам. Конвергенция нервных импульсов объясняется тем, что афферентных нейронов в несколько раз больше, чем эфферентных, поэтому афферентные нейроны образуют на телах и дендритах эфферентных и вставочных нейронов многочисленные синапсы.
Принцип иррадиации. Импульсы, поступающие в центральную нервную систему при сильном и длительном раздражении рецепторов, вызывают возбуждение не только данного рефлекторного центра, но и других нервных центров. Это распространение возбуждения в центральной нервной системе получило название иррадиации. Процесс иррадиации связан с наличием в центральной нервной системе многочисленных ветвлений аксонов и особенно дендритов нервных клеток и цепей вставочных нейронов, которые объединяют друг с другом различные нервные центры.
Принцип реципрокности (сопряженности) в работе нервных центров. Это явление было изучено И. М. Сеченовым, Н. Е. Введенским, Шеррингтоном. Суть его заключается в том, что при возбуждении одних нервных центров деятельность других может затормаживаться. Принцип реципрокности был показан по отношению к нервным центрам мышц-антагонистов - сгибателей и разгибателей конечностей. Наиболее отчетливо он проявляется у животных с удаленным головным мозгом и сохраненным спинным (спинальное животное). Если раздражать у спинального животного (кошка) кожу конечностей, отмечается сгибательный рефлекс данной конечности, а на противоположной стороне в это время наблюдается рефлекс разгибания. Описанные явления связаны с тем, что при возбуждении центра сгибания одной конечности происходит реципрокное торможение центра разгибания этой же конечности. На симметричной стороне имеются обратные взаимоотношения: возбужден центр разгибателей и заторможен центр сгибателей. Только при такой взаимосочетанной (реципрокной) иннервации возможен акт ходьбы.
Может происходить сопряженное, реципрокное торможение и других рефлексов. Под влиянием головного мозга реципрокные отношения могут изменяться. Человек или животное в случае необходимости может сгибать обе конечности, совершать прыжки и т. д.
Реципрокные взаимоотношения центров головного мозга определяют возможность человека овладеть сложными трудовыми процессами и не менее сложными специальными движениями, совершающимися при плавании, акробатических упражнениях и пр.
Принцип общего конечного пути. Этот принцип связан с особенностью строения центральной нервной системы. Эта особенность, как уже указывалось, состоит в том, что афферентных нейронов в несколько раз больше, чем эфферентных, в результате чего различные афферентные импульсы сходятся к общим выходящим путям. Количественные соотношения между нейронами схематически можно представить в виде воронки: возбуждение вливается в центральную нервную систему через широкий раструб (афферентные нейроны) и вытекает из нее через узкую трубку (эфферентные нейроны). Общими путями могут быть не только конечные эфферентные нейроны, но и вставочные.
Импульсы, сходящиеся в общем пути, "конкурируют" друг с другом за использование этого пути. Так достигается упорядочение рефлекторного ответа, соподчинение рефлексов и затормаживание менее существенных. Вместе с тем организм получает возможность реагировать на различные раздражения из внешней и внутренней среды при помощи сравнительно небольшого количества исполнительных органов.
Принцип обратной связи. Этот принцип изучен И. М. Сеченовым, Шеррингтоном, П. К. Анохиным и рядом других исследователей. При рефлекторном сокращении скелетных мышц возбуждаются проприорецепторы. От проприорецепторов нервные импульсы вновь поступают в центральную нервную систему. Этим контролируется точность совершаемых движений. Подобные афферентные импульсы, возникающие в организме в результате рефлекторной деятельности органов и тканей (эффекторов), получили название вторичных афферентных импульсов, или обратной связи.
Обратные связи могут быть положительными и отрицательными. Положительные обратные связи способствуют усилению рефлекторных реакций, отрицательные - их угнетению.
За счет положительных и отрицательных обратных связей осуществляется, например, регуляция относительного постоянства величины артериального давления.
При повышении артериального давления происходит возбуждение механорецепторов дуги аорты, каротидных синусов. Импульсы поступают в сосудодвигательный центр и центр сердечной деятельности, тонус сосудов рефлекторно снижается, одновременно замедляется деятельность сердца и величина артериального давления уменьшается. При понижении артериального давления раздражение механорецепторов указанных рефлексогенных зон вызывает рефлекторное повышение тонуса сосудов, увеличение работы сердца. В этом случае величина артериального давления возрастает.
Вторичные афферентные импульсы (обратные связи) играют также важную роль в регуляции других вегетативных функций: дыхания, пищеварения, выделения.
Принцип доминанты. Принцип доминанты сформулирован А. А. Ухтомским. Этот принцип играет важную роль в согласованной работе нервных центров. Доминанта - это временно господствующий очаг возбуждения в центральной нервной системе, определяющий характер ответной реакции организма на внешние и внутренние раздражения.
Доминантный очаг возбуждения характеризуется следующими основными свойствами: 1) повышенной возбудимостью; 2) стойкостью возбуждения; 3) способностью к суммированию возбуждения; 4) инерцией - доминанта в виде следов возбуждения может длительно сохраняться и после прекращения вызвавшего ее раздражения.
Доминантный очаг возбуждения способен притягивать (привлекать) к себе нервные импульсы из других нервных центров, менее возбужденных в данный момент. За счет этих импульсов активность доминанты еще больше увеличивается, а деятельность других нервных центров подавляется.
Доминанты могут быть экзогенного и эндогенного происхождения. Экзогенная доминанта возникает под влиянием факторов окружающей среды. Например, при чтении интересной книги человек может не слышать звучащую в это время по радио музыку.
Эндогенная доминанта возникает под влиянием факторов внутренней среды организма, главным образом гормонов и других физиологически активных веществ. Например, при понижении содержания питательных веществ в крови, особенно глюкозы, происходит возбуждение пищевого центра, что является одной из причин пищевой установки организма животных и человека.
Доминанта может быть инертной (стойкой), и для ее разрушения необходимо возникновение нового более мощного очага возбуждения.
Доминанта лежит в основе координационной деятельности организма, обеспечивая поведение человека и животных в окружающей среде, а также эмоциональных состояний, реакций внимания. Формирование условных рефлексов и их торможение также связано с наличием доминантного очага возбуждения.
Спинной мозг
Особенности строения спинного мозга. Спинной мозг является наиболее древним и примитивным отделом центральной нервной системы. В центральной части спинного мозга находится серое вещество. Оно состоит преимущественно из нервных клеток и образует выступы - задние, передние и боковые рога. В прилежащих спинальных ганглиях располагаются афферентные нервные клетки. Длинный отросток афферентной клетки находится на периферии и образует воспринимающее окончание (рецептор), а короткий заканчивается у клеток задних рогов. В передних рогах расположены эфферентные клетки (мотонейроны), аксоны которых иннервируют скелетные мышцы; в боковых рогах - нейроны вегетативной нервной системы. В сером веществе находятся многочисленные вставочные нейроны. Среди них обнаружены особые тормозные нейроны - клетки Реншоу, названные так по имени автора, который впервые их описал. Вокруг серого вещества располагается белое вещество спинного мозга. Оно образовано нервными волокнами восходящих и нисходящих путей, соединяющих различные участки спинного мозга друг с другом, а также спинной мозг с головным (рис. 75).
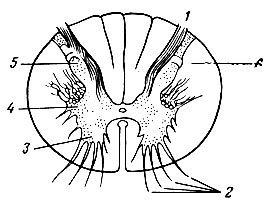
Рис. 75. Поперечный разрез спинного мозга. 1 - задние корешки; 2 - передние корешки; 3 - передний рог; 4 - боковой рог; 5 - задний рог; 6 - белое вещество
Функции спинномозговых корешков. Связь спинного мозга с периферией осуществляется посредством нервных волокон, проходящих в спинномозговых корешках; по ним к спинному мозгу поступают афферентные импульсы и проходят от него на периферию эфферентные импульсы. По обеим сторонам спинного мозга располагается 31 пара передних и задних корешков.
Функции спинномозговых корешков были выяснены при помощи методов перерезки и раздражения.
Выдающийся шотландский анатом и физиолог Белл и французский исследователь Мажанди установили, что при односторонней перерезке передних корешков спинного мозга отмечается паралич конечностей этой же стороны, чувствительность же сохраняется полностью. Перерезка задних корешков приводит к утрате чувствительности, двигательная функция при этом сохраняется.
Таким образом, было показано, что афферентные импульсы поступают в спинной мозг через задние корешки (чувствительные), эфферентные импульсы выходят через передние корешки (двигательные).
Функции и центры спинного мозга. Спинной мозг выполняет две функции: рефлекторную и проводниковую.
Рефлекторная функция спинного мозга. В спинной мозг поступают афферентные импульсы от рецепторов кожи, проприорецепторов двигательного аппарата, интерорецепторов кровеносных сосудов, пищеварительного тракта, выделительных и половых органов. Эфферентные импульсы от спинного мозга идут к скелетным мышцам (за исключением мышц лица), в том числе к дыхательным - межреберным и диафрагме. Кроме того, от спинного мозга по вегетативным нервным волокнам импульсы поступают ко всем внутренним органам, кровеносным сосудам, потовым железам и т. д.
Мотонейроны спинного мозга возбуждаются за счет афферентных импульсов, поступающих к ним от различных рецепторов организма. Однако уровень активности мотонейронов зависит не только от этой афферентации, но и от сложных внутрицентральных взаимоотношений. Большая роль в регуляции деятельности мотонейронов принадлежит нисходящим влияниям головного мозга (коры больших полушарий, ретикулярной формации ствола мозга, мозжечка и др.), а также внутриспинальным воздействиям многочисленных вставочных нейронов. Среди вставочных нейронов особая роль принадлежит клеткам Реншоу. Эти клетки образуют на мотонейронах тормозные синапсы. При возбуждении клеток Реншоу активность мотонейронов притормаживается, что предупреждает перевозбуждение и контролирует их работу. Деятельность мотонейронов спинного мозга контролируется также потоком импульсов, идущих от проприорецепторов мышц (обратная афферентация).
Спинальные рефлексы, т. е. рефлексы, присущие самому спинному мозгу, можно изучить в чистом виде только после отделения спинного мозга от головного (спинальное животное). Первым следствием поперечной перерезки между продолговатым и спинным мозгом является спинальный шок, который длится от нескольких минут до нескольких недель в зависимости от уровня развития центральной нервной системы. Спинальный шок проявляется резким падением возбудимости и угнетением рефлекторных функций всех нервных центров, расположенных ниже места перерезки. В возникновении спинального шока большое значение имеет устранение нервных импульсов, поступающих к спинному мозгу из вышележащих отделов центральной нервной системы, в том числе от нейронов ретикулярной формации ствола мозга.
По прекращении спинального шока постепенно восстанавливаются рефлекторная деятельность скелетных мышц, величина кровяного давления, рефлексы мочеиспускания, дефекации и ряд половых рефлексов. У спинального животного не восстанавливаются произвольные движения, чувствительность и температура тела, а также дыхание. Спинальные животные могут жить только при условии искусственного дыхания. Следовательно, центры, регулирующие эти функции, находятся в вышележащих отделах центральной нервной системы.
Рефлекторные центры спинного мозга. В шейном отделе спинного мозга находятся центр диафрагмального нерва и центр сужения зрачка, в шейном и грудном отделах - центры мышц верхних конечностей, мышц груди, спины и живота, в поясничном отделе - центры мышц нижних конечностей, в крестцовом отделе - центры мочеиспускания, дефекации и половой деятельности, в боковых рогах грудного и поясничного отделов спинного мозга - центры потоотделения и спинальные сосудодвигательные центры.
Изучая нарушения деятельности тех или других групп мышц или отдельных функций у больных людей, можно установить, какой отдел спинного мозга поврежден или функция какого отдела нарушена.
Рефлекторные дуги отдельных рефлексов проходят через определенные сегменты спинного мозга. Возбуждение, возникшее в рецепторе, по центростремительному нерву поступает в соответствующий отдел спинного мозга. Центробежные волокна, выходящие из спинного мозга в составе передних корешков, иннервируют строго определенные участки тела. Схема на рис. 76 показывает, какими сегментами иннервируется каждый участок тела.
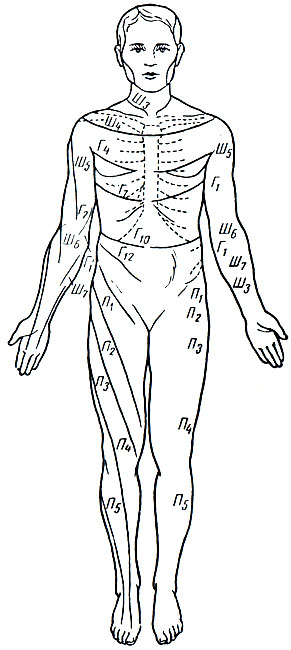
Рис. 76. Схема иннервации отдельных участков кожной поверхности тела человека соответствующими сегментами спинного мозга. Ш - шейный отдел спинного мозга; Г - грудной отдел; П - поясничный отдел. Цифры, находящиеся рядом с указанными буквами, являются показателями позвонков, на уровне которых находится соответствующий отрезок спинного мозга
Проводниковая функция спинного мозга. Через спинной мозг проходят восходящие и нисходящие пути.
Восходящие нервные пути передают информацию от тактильных, болевых, температурных рецепторов кожи и от проприорецепторов мышц через нейроны спинного мозга и другие отделы центральной нервной системы к мозжечку и коре головного мозга.
Нисходящие нервные пути (пирамидный и экстрапирамидный) связывают кору головного мозга, подкорковые ядра и образования ствола мозга с мотонейронами спинного мозга. Они обеспечивают влияние высших отделов центральной нервной системы на деятельность скелетных мышц.
Продолговатый мозг
Непосредственным продолжением спинного мозга у всех позвоночных животных и человека является продолговатый мозг.
Продолговатый мозг и варолиев мост (мост мозга) объединяют под общим названием заднего мозга. Задний мозг вместе со средним и промежуточным мозгом образует ствол мозга. В состав ствола мозга входит большое количество ядер, восходящих и нисходящих путей. Важное функциональное значение имеет находящаяся в стволе мозга, в частности в заднем мозге, ретикулярная формация.
В продолговатом мозге по сравнению со спинным мозгом нет четкого сегментарного распределения серого и белого вещества.
Скопление нервных клеток приводит к образованию ядер, являющихся центрами более или менее сложных рефлексов. Из 12 пар черепных нервов, связывающих головной мозг с периферией организма - его рецепторами и эффекторами, восемь пар (V-XII) берут свое начало в продолговатом мозге.
Продолговатый мозг выполняет две функции - рефлекторную и проводниковую.
Рефлекторная функция продолговатого мозга. В продолговатом мозге находятся центры как относительно простых, так и более сложных рефлексов. За счет продолговатого мозга осуществляются: 1) защитные рефлексы (мигание, слезоотделение, чиханье, кашлевой рефлекс и рефлекс акта рвоты); 2) установочные рефлексы, обеспечивающие тонус мускулатуры, необходимый для поддержания позы и осуществления рабочих актов; 3) лабиринтные рефлексы, способствующие правильному распределению мышечного тонуса между отдельными группами мышц и установке той или иной позы тела; 4) рефлексы, связанные с функциями систем дыхания, кровообращения, пищеварения.
Проводниковая функция продолговатого мозга. Через продолговатый мозг проходят восходящие пути от спинного мозга к головному и нисходящие пути, связывающие кору больших полушарий со спинным мозгом. Продолговатый мозг и варолиев мост имеют собственные проводящие пути, соединяющие ядро и оливу вестибулярного нерва с мотонейронами спинного мозга.
Через восходящие пути и черепные нервы продолговатый мозг получает импульсы от рецепторов мышц лица, шеи, конечностей и туловища, от кожи лица, слизистых оболочек глаз, носовой и ротовой полости, от рецепторов слуха, вестибулярного аппарата, рецепторов гортани, трахеи, легких, интерорецепторов пищеварительного аппарата и сердечно-сосудистой системы.
Функции продолговатого мозга были изучены на бульбарных животных, у которых поперечным разрезом продолговатый мозг отделен от среднего. Следовательно, жизнь бульбарных животных осуществляется за счет деятельности спинного и продолговатого мозга. У таких животных отсутствуют произвольные движения, отмечается потеря всех видов чувствительности, нарушается регуляция температуры тела (теплокровное животное превращается в холоднокровное). У бульбарных животных сохраняются рефлекторные реакции организма и осуществляется регуляция функций внутренних органов.
Рефлекторные центры продолговатого мозга. В продолговатом мозге располагается ряд жизненно важных центров. К ним относятся дыхательный, сердечно-сосудистый и пищевой центры. За счет деятельности этих центров осуществляется регуляция дыхания, кровообращения и пищеварения. Таким образом, основная биологическая роль продолговатого мозга заключается в обеспечении постоянства состава внутренней среды организма.
За счет связей с проприорецепторами продолговатый мозг выполняет роль регулятора тонуса скелетной мускулатуры, прежде всего обеспечивая тоническое напряжение мышц-разгибателей, предназначенных для преодоления силы тяжести организма.
Продолговатый мозг регулирует работу спинного мозга. Эта координационная функция направлена на функциональное объединение всех сегментов спинного мозга, на обеспечение условий для целостной его деятельности. Продолговатый мозг осуществляет более тонкие формы приспособительных реакций организма к внешней среде по сравнению со спинным мозгом.
Средний мозг
К образованиям среднего мозга относят ножки мозга, ядра III (глазодвигательный) и IV (блоковый) пар черепных нервов, четверохолмие, красные ядра и черное вещество. В ножках мозга проходят восходящие и нисходящие нервные пути.
В строении среднего мозга полностью утрачиваются сегментарные признаки. В среднем мозге клеточные элементы образуют сложные скопления в виде ядер. Ядерные образования относятся непосредственно к среднему мозгу, а также к входящей в его состав ретикулярной формации.
Передние бугры четверохолмия получают импульсы от сетчатой оболочки глаз. В ответ на эти сигналы осуществляется регуляция просвета зрачка и аккомодация глаза. Аккомодация - приспособление глаза к ясному видению разноудаленных предметов за счет изменения кривизны хрусталика.
Задние бугры четверохолмия получают импульсы от ядер слуховых нервов, расположенных в продолговатом мозге. Благодаря этому происходит рефлекторная регуляция тонуса мышц среднего уха, а у животных - поворот ушной раковины к источнику звука. Таким образом, при участии передних и задних бугров четверохолмия осуществляются установочные, ориентировочные рефлекторные реакции на световые и звуковые раздражения (движения глаз, поворот головы и даже туловища в сторону светового или звукового раздражителя). При разрушении ядер четверохолмия зрение и слух сохраняются, но отсутствуют ориентировочные реакции на свет и звук.
С деятельностью бугров четверохолмия тесно связана функция ядер III и IV пар черепных нервов, возбуждение которых определяет движение глаз вверх, вниз, в стороны, а также сведение (конвергенция) и разведение глазных осей при переносе взора с удаленных предметов на близкие и обратно,
Красные ядра участвуют в регуляции мышечного тонуса и в проявлении установочных рефлексов, обеспечивающих сохранение правильного положения тела в пространстве. При отделении заднего мозга от среднего тонус мышц-разгибателей повышается, конечности животного напрягаются и вытягиваются, голова запрокидывается. Следовательно, у здорового животного и человека красные ядра несколько притормаживают тонус мышц-разгибателей.
Черное вещество также регулирует мышечный тонус и поддержание позы, участвует в регуляции актов жевания, глотания, кровяного давления и дыхания, т. е. деятельность черного вещества, как и красных ядер, тесно связана с работой продолговатого мозга.
Таким образом, средний мозг регулирует тонус мышц, соответствующим образом его распределяет, что является необходимым условием координированных движений. Средний мозг регулирует ряд вегетативных функций организма (жевание, глотание, кровяное давление, дыхание). За счет среднего мозга расширяется, становится многообразнее рефлекторная деятельность организма (ориентировочные рефлексы на звуковые и зрительные раздражения).
Тонические рефлексы ствола мозга
Образования ствола мозга обеспечивают правильное распределение тонуса между отдельными группами мышц. Рефлексы, обеспечивающие мышечный тонус, получили название тонических. В осуществлении этих рефлексов участвуют мотонейроны спинного мозга, вестибулярные ядра продолговатого мозга, мозжечок, образования среднего мозга (красные ядра). В целостном организме проявление тонических рефлексов контролируется клетками моторной зоны коры больших полушарий.
Тонические рефлексы возникают при изменении положения тела и головы в пространстве за счет возбуждения проприорецепторов мышц, рецепторов вестибулярного аппарата внутреннего уха и тактильных рецепторов кожи.
Тонические рефлексы делят на две группы: статические и статокинетические. Статические рефлексы возникают при изменении положения тела, особенно головы, в пространстве. Статокинетические рефлексы проявляются при перемещении тела в пространстве, при изменении скорости движения (вращательного или прямолинейного).
Таким образом, тонические рефлексы предотвращают возможность нарушения равновесия, потерю активной позы и способствуют восстановлению нарушенной позы.
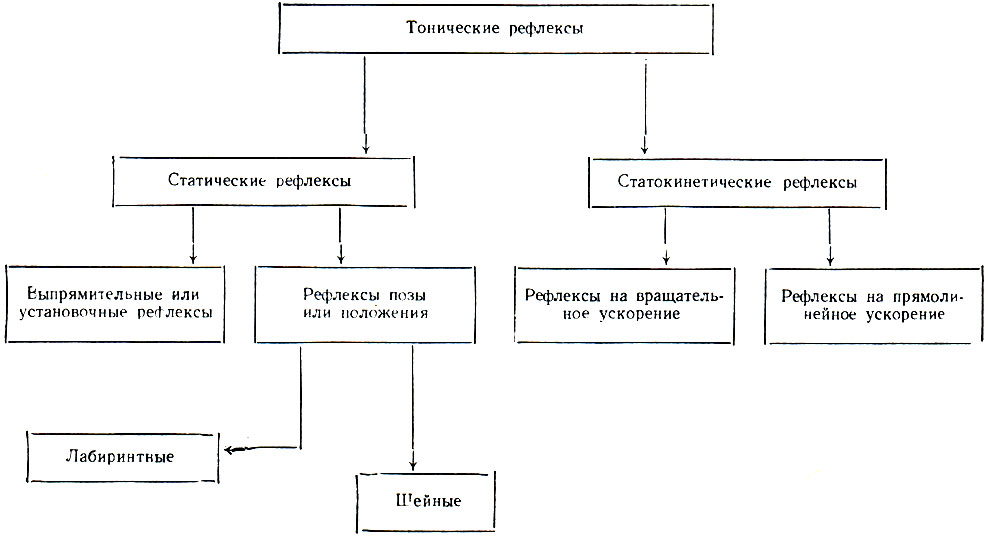
Тонические рефлексы ствола мозга
Промежуточный мозг
Промежуточный мозг - часть переднего отдела ствола мозга. Основными образованиями промежуточного мозга являются зрительные бугры (таламус) и подбугорная область (гипоталамус).
Зрительные бугры - массивное парное образование, они занимают основную массу промежуточного мозга. Наибольших размеров и наивысшей сложности строения зрительные бугры достигают у человека.
Зрительные бугры являются центром всех афферентных импульсов. Через зрительные бугры к коре головного мозга поступает информация от всех рецепторов нашего организма, за исключением обонятельных. Кроме того, от зрительных бугров нервные импульсы передаются к различным образованиям ствола мозга. В зрительных буграх обнаружено большое количество ядерных образований. Функционально их можно разделить на две группы: специфические и неспецифические ядра.
Специфические ядра получают информацию от рецепторов, перерабатывают ее и передают в определенные области коры головного мозга, где возникают соответствующие ощущения (зрительные, слуховые и т. д.).
Неспецифические ядра не имеют прямой связи с рецепторами организма. Они получают импульсы от рецепторов через большое количество переключений (синапсов). Импульсы от этих образований через подкорковые ядра поступают к множеству нейронов, расположенных в различных областях коры головного мозга, вызывая повышение их возбудимости.
При повреждении зрительных бугров у человека наблюдается полная потеря чувствительности или ее снижение на противоположной стороне, выпадает сокращение мимической мускулатуры, которое сопровождает эмоции, также могут возникать расстройства сна, понижение слуха, зрения и т. д.
Гипоталамическая (подбугорная) область участвует в регуляции различных видов обмена веществ (белков, жиров, углеводов, солей, воды), регулирует теплообразование и теплоотдачу, состояние сна и бодрствования. В ядрах гипоталамуса происходит образование ряда гормонов, которые затем депонируются в задней доле гипофиза. Передние отделы гипоталамуса являются высшими центрами парасимпатической нервной системы, задние - симпатической нервной системы. Гипоталамус участвует в регуляции многих вегетативных функций организма.
Базальные ядра
К подкорковым, или базальным, ядрам относятся три парных образования: хвостатое ядро, скорлупа и бледный шар. Базальные ядра расположены внутри больших полушарий, в нижней их части, между лобными долями и промежуточным мозгом. Развитие и клеточное строение у хвостатого ядра и скорлупы одинаковы, поэтому их рассматривают как единое образование - полосатое тело.
Полосатое тело ведает сложными двигательными функциями, участвует в осуществлении безусловно-рефлекторных реакций цепного характера - бег, плавание, прыжки. Эту функцию полосатое тело осуществляет через бледный шар, притормаживая его деятельность. Кроме того, полосатое тело через гипоталамус регулирует вегетативные функции организма, а также вместе с ядрами промежуточного мозга обеспечивает осуществление сложных безусловных рефлексов цепного характера - инстинктов.
Бледный шар является центром сложных двигательных рефлекторных реакций (ходьба, бег), формирует сложные мимические реакции, участвует в обеспечении правильного распределения мышечного тонуса. Свои функции бледный шар осуществляет опосредованно через образования среднего мозга (красные ядра и черное вещество). При раздражении бледного шара наблюдается общее сокращение скелетных мышц противоположной стороны тела. При поражении бледного шара движения теряют свою плавность, становятся неуклюжими, скованными.
Следовательно, деятельность подкорковых ядер не ограничивается их участием в формировании сложных двигательных актов. Они благодаря связям с гипоталамусом участвуют в регуляции обмена веществ и функций внутренних органов.
Таким образом, базальные ядра являются высшими подкорковыми центрами объединения (интеграции) функций организма. У человека и высших позвоночных животных деятельность подкорковых ядер контролируется корой головного мозга.
Ретикулярная формация ствола мозга
Особенности строения. Ретикулярная формация представляет собой скопление особых нейронов, которые своими волокнами образуют своеобразную сеть. Нейроны ретикулярной формации в области ствола мозга были описаны в прошлом веке немецким ученым Дейтерсом. В. М. Бехтерев подобные же структуры обнаружил в области спинного мозга. Нейроны ретикулярной формации образуют скопления, или ядра. Дендриты этих клеток относительно длинные, мало ветвистые, аксоны, напротив, короткие, имеют много ответвлений (коллатералей). Эта особенность обусловливает многочисленные синаптические контакты нейронов ретикулярной формации.
Ретикулярная формация ствола мозга занимает центральное положение в продолговатом мозге, варолиевом мосту, среднем и промежуточном мозге (рис. 77).
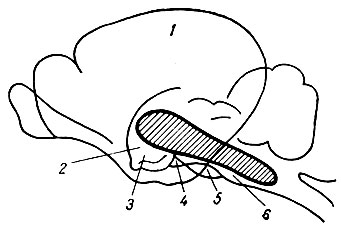
Рис. 77. Схематическое изображение ретикулярной формации. 1 - кора; 2 - зрительный бугор; 3 - подбугорье; 4 - средний мозг; 5 - мост; 6 - продолговатый мозг
Нейроны ретикулярной формации не имеют непосредственных контактов с рецепторами организма. Нервные импульсы при возбуждении рецепторов поступают к ретикулярной формации по коллатералям волокон вегетативной и соматической нервной системы.
Физиологическая роль. Ретикулярная формация ствола мозга оказывает восходящее влияние на клетки коры головного мозга и нисходящее на мотонейроны спинного мозга. Оба эти влияния ретикулярной формации могут быть активирующими или тормозными.
Афферентная импульсация к коре головного мозга поступает по двум путям: специфическому и неспецифическому. Специфический нервный путь обязательно проходит через зрительные бугры и несет нервные импульсы к определенным зонам коры головного мозга, в результате осуществляется какая-либо специфическая деятельность. Например, при раздражении фоторецепторов глаз импульсы через зрительные бугры поступают в затылочную область коры головного мозга и у человека возникают зрительные ощущения.
Неспецифический нервный путь обязательно проходит через нейроны ретикулярной формации ствола мозга. Импульсы к ретикулярной формации поступают по коллатералям специфического нервного пути. Благодаря многочисленным синапсам на одном и том же нейроне ретикулярной формации могут сходиться (конвергировать) импульсы различных значений (световые, звуковые и т. д.), при этом они теряют свою специфичность. От нейронов ретикулярной формации эти импульсы поступают не в какую-то определенную область коры головного мозга, а веерообразно распространяются по ее клеткам, повышая их возбудимость и облегчая тем самым выполнение специфической функции (рис. 78).
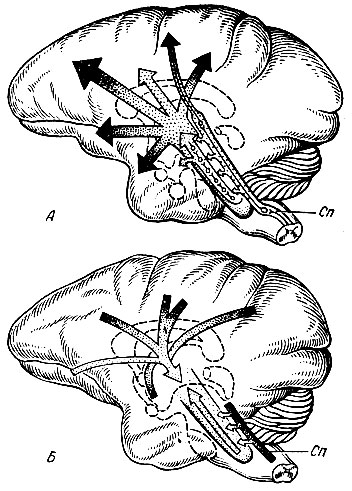
Рис. 78. Связи ретикулярной формации с корой головного мозга. А - схематическое изображение путей распространения восходящих активирующих кору больших полушарий влияний ретикулярной формации; Б - схематическое изображение нисходящих путей коры больших полушарий к ретикулярной формации; Сn - специфические афферентные пути к коре, отдающие коллатерали к ретикулярной формации
В опытах на кошках с вживленными в область ретикулярной формации ствола мозга электродами было показано, что раздражение ее нейронов вызывает пробуждение спящего животного. При разрушении ретикулярной формации животное впадает в длительное сонное состояние. Эти данные свидетельствуют о важной роли ретикулярной формации в регуляций состояния сна и бодрствования. Ретикулярная формация не только оказывает влияние на кору головного мозга, но также посылает в спинной мозг к его двигательным нейронам тормозящие и возбуждающие импульсы. Благодаря этому она участвует в регуляции тонуса скелетной мускулатуры.
В спинном мозге, как уже указывалось, также имеются нейроны ретикулярной формации. Полагают, что они поддерживают на высоком уровне активность нейронов спинного мозга. Функциональное состояние самой ретикулярной формации регулируется корой головного мозга.
Мозжечок
Особенности строения мозжечка. Связи мозжечка с другими отделами центральной нервной системы. Мозжечок - это непарное образование; он располагается позади продолговатого мозга и варолиева моста, граничит с четверохолмиями, сверху прикрыт затылочными долями больших полушарий. В мозжечке различают среднюю часть - червь и расположенные по бокам от него два полушария. Поверхность мозжечка состоит из серого вещества, называемого корой, которая включает тела нервных клеток. Внутри мозжечка располагается белое вещество, представляющее собой отростки этих нейронов.
Мозжечок имеет обширные связи с различными отделами центральной нервной системы за счет трех пар ножек. Нижние ножки соединяют мозжечок со спинным и продолговатым мозгом, средние - с варолиевым мостом и через него с двигательной областью коры головного мозга, верхние - со средним мозгом и гипоталамусом.
Функции мозжечка были изучены на животных, у которых мозжечок удаляли частично или полностью, а также путем регистрации его биоэлектрической активности в покое и при раздражении.
При удалении половины мозжечка отмечается повышение тонуса мышц-разгибателей, поэтому конечности животного вытягиваются, наблюдаются изгиб туловища и отклонение головы в оперированную сторону, иногда качательные движения головой. Часто движения совершаются по кругу в оперированную сторону ("манежные движения"). Постепенно отмеченные нарушения сглаживаются, однако сохраняется некоторая неловкость движений.
При удалении всего мозжечка наступают более выраженные двигательные расстройства. В первые дни после операции животное лежит неподвижно с запрокинутой головой и вытянутыми конечностями. Постепенно тонус мышц-разгибателей ослабевает, появляется дрожание мышц, особенно шейных. В дальнейшем двигательные функции частично восстанавливаются. Однако до конца жизни животное остается двигательным инвалидом: при ходьбе такие животные широко расставляют конечности, высоко поднимают лапы, т. е. у них нарушена координация движений.
Двигательные расстройства при удалении мозжечка были описаны известным итальянским физиологом Лючиани. Основными из них являются: атония - исчезновение или ослабление мышечного тонуса; астения - снижение силы мышечных сокращений. Для такого животного характерно быстро наступающее мышечное утомление; астазия - потеря способности к слитным тетаническим сокращениям. У животных наблюдаются дрожательные движения конечностей и головы. Собака после удаления мозжечка не может сразу поднять лапы, животное делает ряд колебательных движений лапой, перед тем как ее поднять. Если поставить такую собаку, то тело ее и голова все время качаются из стороны в сторону.
В результате атонии, астении и астазии у животного нарушается координация движений: отмечаются шаткая походка, размашистые, неловкие, неточные движения. Весь комплекс двигательных расстройств при поражении мозжечка получил название мозжечковой атаксии (рис. 79).
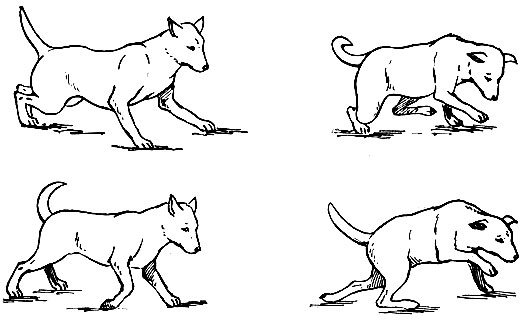
Рис. 79. Походка собаки с удаленным мозжечком (по Дюссер де Берену)
Подобные нарушения наблюдаются и у человека при поражении мозжечка.
Через некоторое время после удаления мозжечка, как уже указывалось, все двигательные расстройства постепенно сглаживаются. Если у таких животных удалить моторную область коры головного мозга, то двигательные нарушения вновь усиливаются. Следовательно, компенсация (восстановление) двигательных расстройств при поражении мозжечка осуществляется при участии коры головного мозга, ее моторной области.
Исследованиями Л. А. Орбели было показано, что при удалении мозжечка наблюдается не только падение мышечного тонуса (атония), но и неправильное его распределение (дистония). Л. А. Орбели установил, что мозжечок влияет и на состояние рецепторного аппарата, а также на вегетативные процессы. Мозжечок оказывает адаптационно-трофическое влияние на все отделы мозга через симпатическую нервную систему, он регулирует обмен веществ в головном мозге и тем самым способствует приспособлению нервной системы к изменяющимся условиям существования.
Таким образом, основными функциями мозжечка являются координация движений, нормальное распределение мышечного тонуса и регуляция вегетативных функций. Свое влияние мозжечок реализует через ядерные образования среднего и продолговатого мозга, через двигательные нейроны спинного мозга. Большая роль в этом влиянии принадлежит двусторонней связи мозжечка с моторной зоной коры головного мозга и ретикулярной формацией ствола мозга (рис. 80).
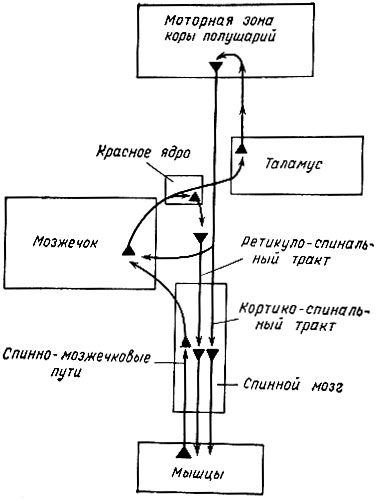
Рис. 80. Связи между мозжечком и красным ядром, зрительным бугром, корой больших полушарий и спинным мозгом (по Гайтону)
При использовании материалов сайта активная ссылка обязательна:
http://anfiz.ru/ 'Анатомия и физиология человека'