Дыхательный центр, его локализация, строение и регуляция активности
Ритмическая последовательность вдоха и выдоха, а также изменение характера дыхательных движений в зависимости от состояния организма (покой, работа различной интенсивности, эмоциональные проявления и т. д.) обусловлены наличием дыхательного центра, расположенного в продолговатом мозге (рис. 27). Дыхательным центром называется совокупность нейронов, обеспечивающих деятельность аппарата дыхания и его приспособление к изменяющимся условиям внешней и внутренней среды.
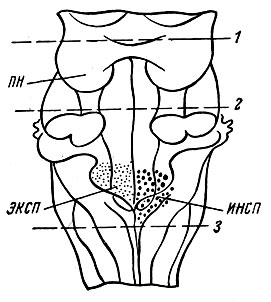
Рис. 27. Функциональная организация дыхательного центра. На рисунке - нижняя часть ствола мозга (вид сзади). ПН - нейроны пневмотаксиса; ИНСП - инспираторные нейроны; ЭКСП - экспираторные нейроны. Центр является двусторонним, но для упрощения схемы на каждой из сторон изображены только одни нейроны. Перерезка выше линии 1 на дыхании не отражается. Перерезка по линии 2 отделяет центр пневмотаксиса. Перерезка ниже линии 3 вызывает прекращение дыхания
Решающее значение в определении локализации дыхательного центра и его активности имели исследования отечественного физиолога Н. А. Миславского, который в 1885 г. показал, что дыхательный центр у млекопитающих находится в продолговатом мозге на две IV желудочка в области ретикулярной формации. Дыхательный центр - это парное, симметрично расположенное образование, в состав которого входят вдыхательная и выдыхательная части.
Результаты исследований Н. А. Миславского легли в основу современных представлений о локализации, строении и функции дыхательного центра. Они подтверждены в экспериментах с использованием микроэлектродной техники и отведения биопотенциалов от различных структур продолговатого мозга. Было показано, что в дыхательном центре имеются две группы нейронов - инспираторные (вдыхательные) и экспираторные (выдыхательные). Обнаружены некоторые особенности в работе дыхательного центра. При спокойном дыхании активна только небольшая часть дыхательных нейронов и, следовательно, в дыхательном центре есть резерв нейронов, который используется при повышенной потребности организма в кислороде. Установлено, что между инспираторными и экспираторными нейронами дыхательного центра существуют функциональные взаимосвязи. Они выражаются в том, что при возбуждении инспираторных нейронов, обеспечивающих фазу вдоха, деятельность экспираторных нервных клеток заторможена и наоборот. Таким образом, одной из причин ритмичной, автоматической деятельности дыхательного центра являются взаимосвязанные функциональные отношения между вдыхательными и выдыхательными нейронами.
Существуют и другие представления о локализации и организации дыхательного центра, которые поддерживают ряд советских и зарубежных физиологов. Предполагают, что в продолговатом мозге локализованы центры вдоха, выдоха и судорожного дыхания. В верхней части моста мозга (варолиев мост) находится пнеймотаксический центр, который контролирует деятельность расположенных ниже центров вдоха и выдоха и обеспечивает правильное чередование циклов дыхательных движений.
Дыхательный центр, расположенный в продолговатом мозге, посылает импульсы к мотонейронам спинного мозга, иннервирующим дыхательные мышцы. Диафрагма иннервируется аксонами мотонейронов, расположенных на уровне III-IV шейных сегментов спинного мозга. Мотонейроны, отростки которых образуют межреберные нервы, иннервирующие межреберные мышцы, расположены в передних рогах грудных сегментов спинного мозга (III-XII).
Регуляция деятельности дыхательного центра
Регуляция деятельности дыхательного центра осуществляется гуморально, за счет рефлекторных воздействий и нервных импульсов, поступающих из вышележащих отделов головного мозга.
По И. П. Павлову, деятельность дыхательного центра зависит от химических свойств крови и от рефлекторных влияний, в первую очередь с легочной ткани.
Гуморальные влияния. Специфическим регулятором активности нейронов дыхательного центра является углекислый газ, который действует на дыхательные нейроны непосредственно и опосредованно. В процессе деятельности нейронов дыхательного центра в них образуются продукты обмена веществ (метаболиты), в том числе и углекислый газ, который оказывает непосредственное влияние на инспираторные нервные клетки, возбуждая их. В ретикулярной формации продолговатого мозга вблизи дыхательного центра обнаружены хеморецепторы, чувствительные к углекислому газу. При увеличении напряжения углекислого газа в крови хеморецепторы возбуждаются и передают эти возбуждения инспираторным нейронам, что приводит к повышению их активности. В лаборатории М. В. Сергиевского получены данные, свидетельствующие о том, что углекислый газ повышает возбудимость нейронов коры головного мозга. В свою очередь клетки коры головного мозга стимулируют активность нейронов дыхательного центра. В механизме стимулирующего влияния углекислого газа на дыхательный центр важное место принадлежит хеморецепторам сосудистого русла. В области каротидных синусов и дуги аорты обнаружены хеморецепторы, чувствительные к изменениям напряжения углекислого газа и кислорода в крови.
Показано, что промывание каротидного синуса или дуги аорты, изолированных в гуморальном отношении, но с сохраненными нервными связями, жидкостью с повышенным содержанием углекислого газа сопровождается стимуляцией дыхания (рефлекс Гейманса). В аналогичных экспериментах было установлено, что повышение напряжения кислорода тормозит активность дыхательного центра.
Опыт с перекрестным кровообращением (опыт Фредерика). Влияние газового состава крови на активность нейронов дыхательного центра доказано в опыте с перекрестным кровообращением (опыт Фредерика). Для этого у двух наркотизированных собак перерезают и перекрестно соединяют сонные артерии и яремные вены (рис. 28). В результате операции голова первой собаки получала кровь от туловища второй, голова же второй собаки - от туловища первой. После установления перекрестного кровообращения зажимают трахею первой собаки, т. е. производят ее удушение. В результате у этой собаки наблюдается остановка дыхания, у второй - резкая одышка.
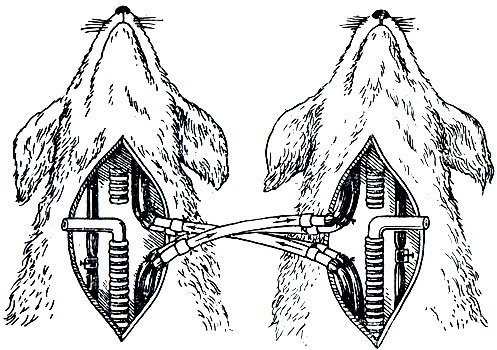
Рис. 28. Схема опыта Фредерика с перекрестным кровообращением
Установленные факты связаны с тем, что в крови первой собаки накапливается избыточное количество углекислого газа, который, поступая с кровью к голове второй собаки, стимулирует активность нейронов дыхательного центра, в результате чего и наблюдается одышка. За счет гипервентиляции кровь второй собаки содержит повышенное количество кислорода и уменьшенное углекислого газа. Поступая к голове первой собаки, кровь второй собаки, богатая кислородом и бедная углекислым газом, тормозит активность нейронов дыхательного центра, и у первой собаки наблюдается остановка дыхания.
Из опыта Фредерика следует, что деятельность дыхательного центра стимулируется при избытке в крови углекислого газа и тормозится при повышении напряжения кислорода. Противоположные сдвиги в активности дыхательного центра наблюдают при снижении концентрации углекислого газа и уменьшении напряжения кислорода в крови.
Механизм влияния углекислого газа на активность нейронов дыхательного центра сложен. Углекислый газ оказывает на дыхательные нейроны прямое (возбуждение клеток коры головного мозга, нейронов ретикулярной формации), а также рефлекторное действие за счет раздражения специальных хеморецепторов сосудистого русла. Следовательно, в зависимости от газового состава внутренней среды организма меняется активность нейронов дыхательного центра, что отражается на характере дыхательных движений.
При оптимальном содержании в крови углекислого газа и кислорода наблюдаются дыхательные движения, отражающие умеренную степень возбуждения нейронов дыхательного центра. Эти дыхательные движения грудной клетки получили название эйпноэ.
Избыточное содержание углекислого газа и недостаток кислорода в крови усиливают активность дыхательного центра, что обусловливает возникновение частых и глубоких дыхательных движений - гиперпноэ. Еще большее нарастание количества углекислого газа в крови приводит к нарушению ритма дыхания и появлению одышки - диспноэ. Понижение концентрации углекислого газа и избыток кислорода в крови угнетают активность дыхательного центра. В этом случае дыхание становится поверхностным, редким и может наступить его остановка - апноэ.
Периодическим называют такой тип дыхания, при котором группы дыхательных движений чередуются с паузами. Продолжительность пауз колеблется в пределах от 5 до 20 с и даже более. При периодическом дыхании типа Чейна-Стокса после паузы появляются слабые, впоследствии усиливающиеся дыхательные движения. При достижении максимума вновь наблюдается ослабление дыхания, а затем оно прекращается - наступает новая пауза. По окончании паузы цикл вновь повторяется. Продолжительность цикла 30-60 с. При снижении возбудимости дыхательного центра, обусловленном недостатком кислорода, наблюдаются и другие типы периодического дыхания.
Причины первого вдоха новорожденного. В организме матери газообмен плода происходит через пупочные сосуды, тесно контактирующие с плацентарной кровью матери. После рождения ребенка и отделения его от плаценты указанная связь нарушается. Метаболические процессы в организме новорожденного приводят к образованию и накоплению углекислого газа, который гуморально возбуждает дыхательный центр. Кроме того, изменение условий существования ребенка приводит к возбуждению экстеро- и проприорецепторов, что также является одним из механизмов, принимающих участие в возникновении первого вдоха.
Рефлекторные влияния на активность нейронов дыхательного центра. На активность нейронов дыхательного центра выраженное влияние оказывают рефлекторные воздействия. Различают постоянные и непостоянные (эпизодические) рефлекторные влияния на дыхательный центр.
Постоянные рефлекторные влияния возникают в результате раздражения рецепторов альвеол (рефлекс Геринга-Брейера), корня легкого и плевры (пульмоторакальный рефлекс), хеморецепторов дуги аорты и каротидных синусов (рефлекс Гейманса), механорецепторов указанных сосудистых областей, проприорецепторов дыхательных мышц.
Наиболее важным рефлексом этой группы является рефлекс Геринга - Брейера. В альвеолах легких заложены механорецепторы растяжения и спадения, являющиеся чувствительными нервными окончаниями блуждающего нерва. Рецепторы растяжения возбуждаются при обычном и максимальном вдохе, т. е. любое увеличение объема легочных альвеол возбуждает эти рецепторы. Рецепторы спадения становятся активными только в условиях патологии (при максимальном спадении альвеол).
В экспериментах на животных установлено, что при увеличении объема легких (вдувание в легкие воздуха) наблюдается рефлекторный выдох, выкачивание же воздуха из легких приводит к быстрому рефлекторному вдоху. Указанные реакции не возникали при перерезке блуждающих нервов. Следовательно, нервные импульсы в центральную нервную систему поступают по блуждающим нервам.
Рефлекс Геринга - Брейера относится к механизмам саморегуляции дыхательного процесса, обеспечивая смену актов вдоха и выдоха. При растяжении альвеол во время вдоха нервные импульсы от рецепторов растяжения по блуждающему нерву идут к экспираторным нейронам, которые, возбуждаясь, тормозят активность инспираторных нейронов, что приводит к пассивному выдоху. Легочные альвеолы спадаются, и нервные импульсы от рецепторов растяжения уже не поступают к экспираторным нейронам. Активность их падает, что создает условия для повышения возбудимости инспираторной части дыхательного центра и активного вдоха. Кроме того, активность инспираторных нейронов повышается при нарастании концентрации углекислого газа в крови, что также способствует осуществлению акта вдоха.
Таким образом, саморегуляция дыхания осуществляется на основе взаимодействия нервного и гуморального механизмов регуляции активности нейронов дыхательного центра.
Пульмоторакальный рефлекс возникает при возбуждении рецепторов, заложенных в легочной ткани и плевре. Проявляется этот рефлекс при растяжении легких и плевры. Рефлекторная дуга замыкается на уровне шейных и грудных сегментов спинного мозга. Конечным эффектом рефлекса является изменение тонуса дыхательной мускулатуры, благодаря чему происходит увеличение или уменьшение среднего объема легких.
К дыхательному центру постоянно идут нервные импульсы от проприорецепторов дыхательных мышц. Во время вдоха происходит возбуждение проприорецепторов дыхательных мышц и нервные импульсы от них поступают к инспираторным нейронам дыхательного центра. Под влиянием нервных импульсов активность инспираторных нейронов тормозится, что способствует наступлению выдоха.
Непостоянные рефлекторные влияния на активность дыхательных нейронов связаны с возбуждением разнообразных по своим функциям экстеро- и интерорецепторов.
К непостоянным рефлекторным воздействиям, оказывающим влияние на активность дыхательного центра, относятся рефлексы, возникающие при раздражении рецепторов слизистой оболочки верхних дыхательных путей, носа, носоглотки, температурных и болевых рецепторов кожи, проприорецепторов скелетных мышц, интерорецепторов. Так, например, при внезапном вдыхании паров аммиака, хлора, сернистого ангидрида, табачного дыма и некоторых других веществ происходит раздражение рецепторов слизистой оболочки носа, глотки, гортани, что приводит к рефлекторному спазму голосовой щели, а иногда даже мускулатуры бронхов и рефлекторной задержке дыхания.
При раздражении эпителия дыхательных путей накопившейся пылью, слизью, а также попавшими химическими раздражителями и инородными телами наблюдаются чиханье и кашель. Чиханье возникает при раздражении рецепторов слизистой оболочки носа, а кашель - при возбуждении рецепторов гортани, трахеи, бронхов.
Кашель и чиханье начинаются с глубокого вдоха, который возникает рефлекторно. Затем происходит спазм голосовой щели и одновременно активный выдох. Вследствие этого давление в альвеолах и воздухоносных путях значительно возрастает. Следующее за этим раскрытие голосовой щели приводит к выбросу воздуха из легких толчком в дыхательные пути и наружу через нос (при чиханье) или через рот (при кашле). Пыль, слизь, инородные тела увлекаются этой струей воздуха и выбрасываются из легких и дыхательных путей.
Кашель и чиханье в условиях нормы относят к категории защитных рефлексов. Эти рефлексы называют защитными потому, что они препятствуют попаданию вредных веществ в дыхательные пути или же способствуют их удалению.
Раздражение температурных рецепторов кожи, в частности Холодовых, приводит к рефлекторной задержке дыхания. Возбуждение болевых рецепторов кожи, как правило, сопровождается учащением дыхательных движений.
Возбуждение проприорецепторов скелетных мышц обусловливает стимуляцию акта дыхания. Повышенная активность дыхательного центра в этом случае является важным приспособительным механизмом, обеспечивающим увеличенные потребности организма в кислороде при мышечной работе.
Раздражение интерорецепторов, например механорецепторов желудка при его растяжении, приводит к торможению не только сердечной деятельности, но и дыхательных движений.
При возбуждении механорецепторов сосудистых рефлексогенных зон (дуга аорты, каротидные синусы) в результате изменения величины артериального давления наблюдаются сдвиги в активности дыхательного центра. Так, повышение артериального давления сопровождается рефлекторной задержкой дыхания, понижение приводит к стимуляции дыхательных движений.
Таким образом, нейроны дыхательного центра чрезвычайно чувствительны к воздействиям, обусловливающим возбуждение экстеро-, проприо- и интерорецепторов, что приводит к изменению глубины и ритма дыхательных движений в соответствии с условиями жизнедеятельности организма.
Влияние коры головного мозга на активность дыхательного центра. Регуляция дыхания корой больших полушарий имеет свои качественные особенности. В опытах с прямым раздражением электрическим током отдельных областей коры головного мозга было показано выраженное влияние ее на глубину и частоту дыхательных движений. Результаты исследований М. В. Сергиевского и его сотрудников, полученные при непосредственном раздражении различных участков коры больших полушарий электрическим током в острых, полухронических и хронических опытах (вживленные электроды), свидетельствуют о том, что нейроны коры не всегда оказывают однозначное влияние на дыхание. Конечный эффект зависит от ряда факторов, главным образом от силы, продолжительности и частоты применяемых раздражений, функционального состояния коры головного мозга и дыхательного центра.
Важные факты были установлены Э. А. Асратяном и его сотрудниками. Было обнаружено, что у животных с удаленной корой головного мозга отсутствовали приспособительные реакции внешнего дыхания на изменения условий жизнедеятельности. Так, мышечная активность у таких животных не сопровождалась стимуляцией дыхательных движений, а приводила к длительной одышке и дискоординации дыхания.
Для оценки роли коры головного мозга в регуляции дыхания большое значение имеют данные, полученные с помощью метода условных рефлексов. Если у человека или животных звук метронома сопровождать вдыханием газовой смеси с повышенным содержанием углекислого газа, то это приведет к увеличению легочной вентиляции. Через 10-15 сочетаний изолированное включение метронома (условный сигнал) вызовет стимуляцию дыхательных движений - образовался условный дыхательный рефлекс на избранное количество ударов метронома в единицу времени.
Учащение и углубление дыхания, которые наступают до начала физической работы или спортивных состязаний, также осуществляются по механизму условных рефлексов. Эти изменения в дыхательных движениях отражают сдвиги в активности дыхательного центра и имеют приспособительное значение, способствуя подготовке организма к выполнению работы, требующей большой затраты энергии и усиления окислительных процессов.
По мнению М. Е. Маршака, корковая регуляция дыхания обеспечивает необходимый уровень легочной вентиляции, темп и ритм дыхания, постоянство уровня углекислого газа в альвеолярном воздухе и артериальной крови.
Приспособление дыхания к внешней среде и сдвигам, наблюдаемым во внутренней среде организма, связано с обширной нервной информацией, поступающей в дыхательный центр, которая предварительно перерабатывается, главным образом в нейронах моста мозга (варолиев мост), среднего и промежуточного мозга и в клетках коры головного мозга.
Таким образом, регуляция активности дыхательного центра сложна. По М. В. Сергиевскому, она состоит из трех уровней.
Первый уровень регуляции представлен спинным мозгом. Здесь располагаются центры диафрагмальных и межреберных нервов. Эти центры обусловливают сокращение дыхательных мышц. Однако этот уровень регуляции дыхания не может обеспечить ритмичную смену фаз дыхательного цикла, так как огромное количество афферентных импульсов от дыхательного аппарата, минуя спинной мозг, направляется непосредственно в продолговатый мозг.
Второй уровень регуляции связан с функциональной активностью продолговатого мозга. Здесь находится дыхательный центр, который воспринимает разнообразные афферентные импульсы, идущие от дыхательного аппарата, а также от основных рефлексогенных сосудистых зон. Этот уровень регуляции обеспечивает ритмичную смену фаз дыхания и активность спинномозговых мотонейронов, аксоны которых иннервируют дыхательную мускулатуру.
Третий уровень регуляции - это верхние отделы головного мозга, включающие и корковые нейроны. Только при наличии коры полушарий головного мозга возможно адекватное приспособление реакций системы дыхания к изменяющимся условиям существования организма.
Дыхание при физической работе
Физическая нагрузка сопровождается значительными сдвигами в активности органов и физиологических систем организма. Повышенные энерготраты обеспечиваются увеличением утилизации кислорода, что приводит к нарастанию содержания углекислого газа в жидкостях и тканях организма. Сдвиги в химическом составе внутренней среды организма обусловливают повышение функциональной активности системы органов дыхания. Так, у тренированных людей при напряженной мышечной работе объем легочной вентиляции возрастает до 5·10-2 м3 и даже до 1·10-1 м3 (50 и даже 100 л/мин) по сравнению с 5·10-3-8·10-3 м3 (5-8 л/мин) в состоянии относительного физиологического покоя.
Повышение минутного объема дыхания при физической нагрузке связано с увеличением глубины и частоты дыхательных движений. При этом у тренированных людей в основном изменяется глубина дыхания, у нетренированных - частота дыхательных движений.
Сдвиги в функциональной активности системы дыхания при физической нагрузке обусловливаются нервными и гуморальными механизмами. При физической нагрузке увеличивается концентрация в крови и тканях углекислого газа и молочной кислоты, которые стимулируют нейроны дыхательного центра как гуморальным путем, так и за счет нервных импульсов, поступающих от сосудистых рефлексогенных зон. Кроме того, нейроны дыхательного центра стимулируются нервными влияниями, идущими от проприорецепторов дыхательных и скелетных мышц. Наконец, активность нейронов дыхательного центра обеспечивается потоком нервных импульсов, поступающих из клеток коры головного мозга, обладающих высокой чувствительностью к недостатку кислорода и избытку углекислого газа.
Одновременно с изменениями в системе дыхания при физической нагрузке возникают приспособительные реакции в сердечно-сосудистой системе. Увеличиваются частота и сила сердечных сокращений, повышается кровяное давление, происходит перераспределение сосудистого тонуса - расширяются сосуды работающих мышц и суживаются сосуды других областей. Кроме того, открывается дополнительное количество капилляров в работающих органах и происходит выброс крови из депо.
Значительную роль в координации функций органов и физиологических систем при физической нагрузке играет кора головного мозга. Так, в предстартовом состоянии у спортсменов отмечается увеличение силы и частоты сердечных сокращений, возрастает легочная вентиляция, повышается кровяное давление. Следовательно, условнорефлекторный механизм - один из важнейших нервных механизмов адаптации организма к меняющимся условиях внешней среды.
Система дыхания обеспечивает возросшие потребности организма в кислороде. Системы же кровообращения и крови, перестраиваясь на новый функциональный уровень, способствуют транспорту кислорода к тканям и углекислого газа к легким.
При использовании материалов сайта активная ссылка обязательна:
http://anfiz.ru/ 'Анатомия и физиология человека'