Глава IV. Физиология системы дыхания
Сущность и значение дыхания для организма
Дыхание - это неотъемлемый признак жизни. Мы дышим постоянно с момента рождения и до самой смерти, дышим днем и ночью во время глубокого сна, в состоянии здоровья и болезни.
В организме человека и животных запасы кислорода ограничены, поэтому организм нуждается в непрерывном поступлении кислорода из окружающей среды. Также постоянно, и непрерывно из организма должен удаляться углекислый газ, который всегда образуется в процессе обмена веществ и в больших количествах является токсичным соединением.
Дыхание - сложный непрерывный процесс, в результате которого постоянно обновляется газовый состав крови и происходит биологическое окисление в тканях. В этом заключается его сущность.
Нормальное функционирование организма человека возможно только при условии пополнения энергией, которая непрерывно расходуется. Организм получает энергию за счет окисления органических веществ - белков, жиров, углеводов. При этом освобождается скрытая химическая энергия, которая является источником жизнедеятельности, развития и роста организма. Таким образом, значение дыхания состоит в поддержании в организме оптимального уровня окислительно-восстановительных процессов.
В процессе дыхания принято различать три звена: внешнее, или легочное, дыхание, транспорт газов кровью и внутреннее, или тканевое, дыхание.
Внешнее дыхание - это газообмен между организмом и окружающим его атмосферным воздухом.
Внешнее дыхание может быть разделено на два этапа - обмен газов между атмосферным и альвеолярным воздухом и газообмен между кровью легочных капилляров и альвеолярным воздухом.
Внешнее дыхание осуществляется за счет активности аппарата внешнего дыхания.
Аппарат внешнего дыхания включает в себя дыхательные пути, легкие, плевру, скелет грудной "клетки и ее мышцы, а также диафрагму. Основной функцией аппарата внешнего дыхания является обеспечение организма кислородом и освобождение его от избытка углекислого газа.
О функциональном состоянии аппарата внешнего дыхания можно судить по ритму, глубине, частоте дыхания, по величине легочных объемов, по показателям поглощения кислорода и выделения углекислого газа и т. д.
Транспорт газов осуществляется кровью. Он обеспечивается разностью парциального давления газов по пути их следования: кислорода от легких к тканям, углекислого газа от клеток к легким.
Внутреннее, или тканевое, дыхание также может быть разделено на два этапа. Первый этап - это обмен газов между кровью и тканями, второй связан с потреблением кислорода клетками и выделением ими углекислого газа (клеточное дыхание).
Состав вдыхаемого, выдыхаемого и альвеолярного воздуха
Человек дышит атмосферным воздухом, который имеет следующий состав: 20,94% кислорода, 0,03% углекислого газа, 79,03% азота. В выдыхаемом воздухе обнаруживается 16,3% кислорода, 4% углекислого газа, 79,7% азота.
Состав выдыхаемого воздуха весьма непостоянен и зависит от интенсивности обмена веществ, а также от частоты и глубины дыхания. Стоит задержать дыхание или сделать несколько глубоких дыхательных движений, как состав выдыхаемого воздуха изменится.
Сравнение состава вдыхаемого и выдыхаемого воздуха служит доказательством существования внешнего дыхания.
Альвеолярный воздух по составу во многом отличается от атмосферного, что вполне закономерно. Именно в альвеолах происходит обмен газов между воздухом и кровью, при этом в кровь диффундирует кислород, а из крови - углекислый газ. В результате в альвеолярном воздухе резко уменьшается содержание кислорода и возрастает количество углекислого газа. Процентное содержание отдельных газов в альвеолярном воздухе следующее: 14,2-14,6% кислорода, 5,2-5,7% углекислого газа, 79,7-80% азота. Альвеолярный воздух отличается по составу и от выдыхаемого воздуха. Это объясняется тем, что выдыхаемый воздух содержит смесь газов из альвеол и вредного пространства.
Строение легких
Легкие - парные образования сложного строения, расположенные в герметически замкнутой грудной полости. Их воздухоносные пути представлены носоглоткой, гортанью, трахеей. Трахея в грудной полости делится на два бронха - правый и левый, каждый из которых, многократно разветвляясь, образует так называемое бронхиальное дерево. Мельчайшие бронхи - бронхиолы - на концах расширяются в слепые пузырьки - легочные альвеолы. Совокупность альвеол и образует ткань легких (рис. 21).
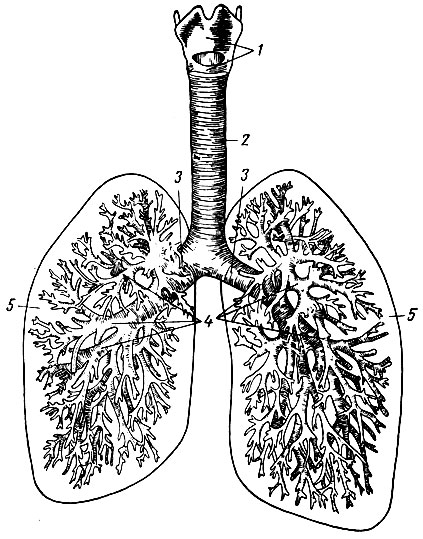
Рис. 21. Схема воздухоносных путей. 1 - гортань; 2 - трахея (дыхательное горло); 3 - бронхи; 4 - бронхиальное дерево; 5 - легкое
Слизистая оболочка трахеи и бронхов покрыта многослойным мерцательным эпителием, реснички которого колеблются по направлению к ротовой, полости. Кроме того, слизистая оболочка содержит многочисленные железы, выделяющие слизь. Слизь увлажняет вдыхаемый воздух. Благодаря наличию носовых раковин и густой сети капилляров в слизистой оболочке, а также мерцательному эпителию воздух, поступая в дыхательные пути, прежде чем достигнуть альвеол, согревается, увлажняется и в значительной степени очищается от механических примесей (частичек пыли).
В дыхательных путях воздух близко не соприкасается с кровью, поэтому газообмен здесь не происходит и состав воздуха не меняется. Пространство, заключенное в этих дыхательных путях, называется мертвым, или вредным. При спокойном дыхании объем воздуха в мертвом пространстве составляет 1,4·10-4-1,5·10-4 м3 (140-150 мл).
Строение легких обеспечивает выполнение ими дыхательной функции. Тонкая стенка альвеол состоит из однослойного эпителия, легко проходимого для газов. Наличие эластических элементов и гладких мышечных волокон обеспечивает быстрое и легкое растяжение альвеол, благодаря чему они могут вмещать большие количества воздуха. Каждая альвеола оплетена густой сетью капилляров, на которые разветвляется легочная артерия (рис. 22). Оба легких содержат 300-400 млн. микроскопических альвеол, диаметр которых у взрослого человека составляет 0,2·10-3 м (0,2 мм). Благодаря большому количеству альвеол образуется громадная дыхательная поверхность легких. У человека массой 70 кг в состоянии вдоха дыхательная поверхность легких равна 80-100 м2, при выдохе - 40-50 м2.
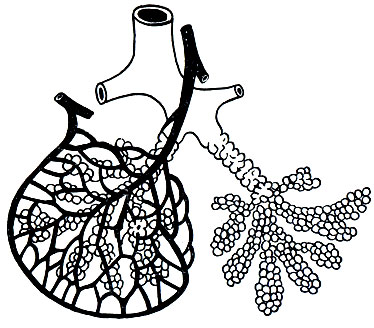
Рис. 22. Схема доли легкого. Левая доля оплетена сетью капилляров
Каждое легкое покрыто снаружи серозной оболочкой - плеврой, состоящей из двух листков - пристеночного и висцерального. Между листками плевры имеется узкая капиллярная щель, содержащая небольшое количество серозной жидкости. Неправильно эту щель называть полостью. В норме полости нет, она потенциальна, т. е. может возникнуть, если листки плевры будут раздвинуты экссудатом, образующимся в условиях патологии, или же воздухом, например при травме грудной клетки.
Расправление и спадение легочных альвеол, а также движение воздуха по воздухоносным путям сопровождается возникновением дыхательных шумов, которые можно исследовать методом выслушивания - аускультации.
Отрицательное давление в плевральной щели
Давление в плевральной щели и в средостении в норме всегда отрицательное. Убедиться в этом можно, измерив давление в плевральной щели. Для этого полую иглу, соединенную с манометром, вводят между двумя листками плевры. Во время спокойного вдоха давление в плевральной щели на 1,197 кПа (9 мм. рт. ст.) ниже атмосферного, во время спокойного выдоха на 0,798 кПа (6 мм рт. ст.).
Отрицательное внутригрудное давление и увеличение его во время вдоха имеет большое физиологическое значение. За счет отрицательного внутриплеврального давления альвеолы всегда находятся в растянутом состоянии, что значительно увеличивает дыхательную поверхность легких, особенно во время вдоха. Отрицательное внутригрудное давление играет значительную роль в гемодинамике, обеспечивая венозный возврат крови к сердцу и улучшая кровообращение в легочном круге, особенно в фазу вдоха. Присасывающее действие грудной клетки способствует также и лимфообращению. Наконец, отрицательное внутригрудное давление является фактором, способствующим продвижению пищевого комка по пищеводу, в нижнем отделе которого давление на 0,46 кПа (3,5 мм рт. ст.) ниже атмосферного.
Дыхательный цикл
Дыхательный цикл состоит из вдоха, выдоха и дыхательной паузы. Обычно вдох короче выдоха. Длительность вдоха у взрослого человека 0,9-4,7 с, длительность выдоха 1,2-6 с. Продолжительность вдоха и выдоха зависит в основном от рефлекторных воздействий, идущих от рецепторов легочной ткани. Дыхательная пауза - это непостоянная составная часть дыхательного цикла. Она различна по продолжительности и даже может отсутствовать.
Дыхательные движения совершаются с определенным ритмом и частотой, которые устанавливают по количеству экскурсий грудной клетки в 60 с (1 мин). У взрослого человека частота дыхательных движений составляет 12-18 в 60 с (1 мин). У детей дыхание поверхностное и поэтому более частое, чем у взрослых. Так, новорожденный дышит около 60 раз в 60с (1 мин), ребенок 5-летнего возраста - 25 раз в 60 с (1 мин). В любом возрасте частота дыхательных движений меньше количества сердечных сокращений в 4-5 раз.
Глубину дыхательных движений определяют по амплитуде экскурсий грудной клетки и с помощью специальных методов, позволяющих исследовать величины легочных объемов.
На частоту и глубину дыхания влияют многие факторы, в частности эмоциональное состояние, умственная нагрузка, изменение химического состава крови, степень тренированности организма, уровень и интенсивность обмена веществ. Чем чаще и глубже дыхательные движения, тем больше кислорода поступает в легкие и соответственно большее количество углекислого газа выводится. Редкое и поверхностное дыхание может привести к недостатку снабжения клеток и тканей организма кислородом. Это в свою очередь сопровождается снижением их функциональной активности. В значительной степени изменяется частота и глубина дыхательных движений при патологических состояниях организма, особенно при заболеваниях органов дыхания.
Механизм вдоха. Акт вдоха (инспирация) совершается вследствие увеличения объема грудной клетки в трех направлениях - вертикальном, сагиттальном (переднезаднем, грудинно-позвоночном) и фронтальном (реберном). Изменение размеров грудной полости происходит за счет сокращения дыхательных мышц - наружных межреберных, межхрящевых и диафрагмы (рис. 23). При сокращении наружных межреберных и межхрящевых мышц ребра принимают более горизонтальное положение, поднимаясь кверху, при этом нижний конец грудины отходит вперед. Благодаря движению ребер при вдохе размеры грудной клетки увеличиваются в поперечном и продольном направлении. В результате сокращения мышечных волокон диафрагмы купол ее уплощается и опускается: органы брюшной полости оттесняются вниз, в стороны и вперед, в итоге объем грудной клетки увеличивается в вертикальном направлении.
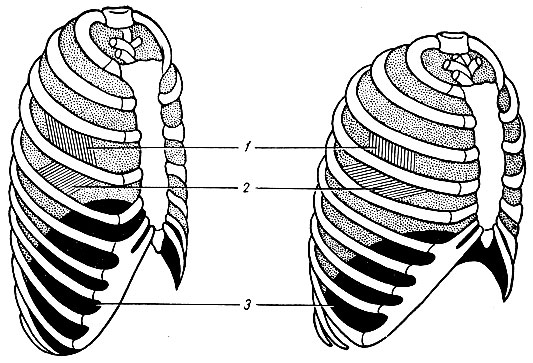
Рис. 23. Схема положения грудной клетки и диафрагмы при выдохе (слева) и вдохе (справа). 1 - наружные межреберные мышцы; 2 - внутренние межреберные мышцы; 3 - диафрагма
В зависимости от преимущественного участия в акте вдоха мышц грудной клетки и диафрагмы различают грудной, или реберный, и брюшной, или диафрагмальный, тип дыхания. У мужчин преобладает брюшной тип дыхания, у женщин - грудной.
В некоторых случаях, например при физической работе, при одышке, в акте вдоха могут участвовать так называемые вспомогательные мышцы - мышцы плечевого пояса и шеи (грудные и передние зубчатые мышцы, грудино-ключично-сосцевидные).
При вдохе легкие пассивно следуют за увеличивающейся в размерах грудной клеткой. Дыхательная поверхность легких увеличивается, давление же в них понижается и становится на 0,26 кПа (2 мм рт. ст.) ниже атмосферного. Это способствует поступлению воздуха через воздухоносные пути в легкие. Однако только на высоте вдоха происходит заполнение воздухом расширенных альвеол. Быстрому выравниванию давления в легких препятствует голосовая щель, так как в этом месте воздухоносные пути сужены.
Механизм выдоха. Акт выдоха (экспирация) осуществляется в результате расслабления наружных межреберных мышц и поднятия купола диафрагмы. При этом грудная клетка возвращается в исходное положение и дыхательная поверхность легких уменьшается. Сужение воздухоносных путей в области голосовой щели обусловливает медленный выход воздуха из легких. В начале фазы выдоха давление в легких становится на 0,40-0,53 кПа (3-4 мм рт. ст.) выше атмосферного, что облегчает выход воздуха из них в окружающую среду.
Механизм изменений объема легких при дыхании может быть продемонстрирован с помощью модели Дондерса (рис. 24). Нижнюю часть лишенной дна широкой стеклянной бутыли затягивают резиновой пленкой, которая имитирует работу диафрагмы. Горлышко бутыли закрывают пробкой, через которую пропускают стеклянную трубку, к концу которой привязывают трахею с легкими мелкого лабораторного животного (крыса, кошка, кролик).
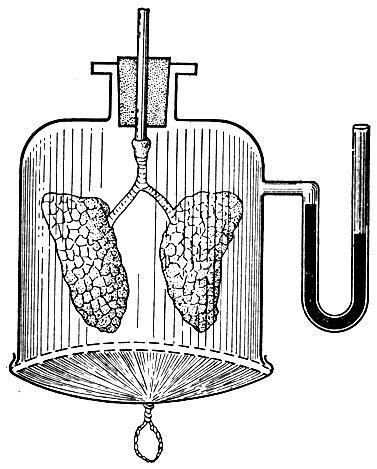
Рис. 24. Модель Дондерса, демонстрирующая механику дыхательного акта. Объяснение в тексте
Таким образом, сосуд герметически закрыт и не сообщается с атмосферным воздухом, так как стеклянная трубка соединена с легкими. Если оттянуть резиновое дно бутыли книзу, то объем ее увеличится и давление в ней станет ниже атмосферного. Это вызывает растяжение легочной ткани, и атмосферный воздух начнет поступать в легкие. Однако давление воздуха внутри бутыли между ее стенками и наружной поверхностью легких все же остается ниже атмосферного, так как упругие свойства легочной ткани препятствуют ее растяжению. Если отпустить резиновое дно бутыли объем ее уменьшится, прекратится действие силы, растягивающей легкие. Благодаря своей эластичности легочная ткань сжимается, давление в легких повышается и воздух из них выходит наружу.
Модель Дондерса доказывает, что непосредственной причиной изменения объема легких при вдохе и выдохе являются изменения размеров грудной клетки и давления в плевральной щели. Воздух же поступает в легкие и выходит из них вследствие колебаний внутрилегочного давления.
Легочные объемы. Легочная вентиляция
Для исследования функционального состояния аппарата внешнего дыхания как в клинической практике, так и в физиологических лабораториях широко используют определение легочных объемов.
Различают четыре основных положения грудной клетки, которым соответствуют четыре основных объема легких: дыхательный, дополнительный, резервный и остаточный.
Дыхательный объем - это количество воздуха, которое человек вдыхает и выдыхает при спокойном дыхании. Его объем составляет 3·10-4-7·10-4 м3 (300-700 мл). Дыхательный объем обеспечивает поддержание определенного уровня парциального давления кислорода и углекислого газа в альвеолярном воздухе, способствуя тем самым нормальному напряжению газов в артериальной крови.
Дополнительный воздух, или резервный объем воздуха, - это количество воздуха которое может быть введено в легкие, если вслед за спокойным вдохом произвести максимальный вдох. Резервный объем вдоха равняется 1,5·10-3-2·10-3 м3 (1500-2000 мл). Он определяет способность легких к добавочному расширению, необходимость в котором имеется при увеличении потребности организма в газообмене.
Резервный воздух, или резервный объем воздуха, - это тот объем воздуха, который удаляется из легких, если вслед за спокойным вдохом и выдохом произвести максимальный выдох. Резервный объем выдоха составляет 1,5·10-3-2·10-3 м3 (1500-2000 мл). Он определяет степень постоянного растяжения легких.
Остаточный объем - это объем воздуха, который остается в легких после максимально глубокого выдоха. Остаточный объем равняется 1·10-3-1,5·10-3 м3 (1000-1500 мл).
Дыхательный объем, резервные объемы вдоха и выдоха составляют так называемую жизненную емкость легких.
Жизненная емкость легких - это самое глубокое дыхание, на которое способен данный человек. Она определяется тем количеством воздуха, которое может быть удалено из легких, если после максимального вдоха сделать максимальный выдох.
Жизненная емкость легких у мужчин молодого возраста составляет 3,5·10-3-4,8·10-3 м3 (3,5-4,8 л), у женщин - 3·10-3-3,5·10-3 м3 (3-3,5 л). Показатели жизненной емкости легких весьма изменчивы. Они зависят от пола, роста, возраста, массы, положения тела, состояния дыхательных мышц, уровня возбудимости дыхательного центра и других факторов. По величине жизненной емкости легких в известной степени можно судить о функциональных возможностях аппарата внешнего дыхания.
Общая емкость легких состоит из жизненной емкости легких и остаточного объема воздуха.
Коллапсный воздух - это минимальное количество воздуха, которое остается в легких после двустороннего открытого пневмоторакса. Наличие коллапсного воздуха в легких доказывается простым опытом. Установлено, что кусочек ткани легкого после пневмоторакса плавает в воде, а легкое мертворожденного, не дышавшего плода тонет.
Частота и глубина дыхания может оказать значительное влияние на циркуляцию воздуха в легких во время дыхания, или на легочную вентиляцию.
Легочная вентиляция - количество воздуха, обмениваемое в 1 мин. За счет легочной вентиляции обновляется альвеолярный воздух и в нем поддерживается парциальное давление кислорода и углекислого газа на таком уровне, который обеспечивает нормальный газообмен. Легочную вентиляцию определяют путем умножения дыхательного объема на количество дыханий в 1 мин (минутный объем дыхания). У взрослого человека в состоянии относительного физиологического покоя легочная вентиляция составляет 6·10-3-8·10-3 м (6-8 л) в 1 мин. Определение минутного объема дыхания имеет диагностическое значение при легочной патологии.
Легочные объемы могут быть определены с помощью специальных приборов спирометра и спирографа. Спирографический метод позволяет регистрировать величины легочных объемов.
Транспорт газов кровью
Мы рассмотрели только одну сторону дыхательного процесса - внешнее дыхание, т. е. обмен газов между организмом и окружающей его средой.
Местом же потребления кислорода и образования углекислого газа являются все клетки организма, где осуществляется тканевое, или внутреннее, дыхание. Вследствие этого, когда речь идет о дыхании в целом, необходимо учитывать пути и условия переноса газов: кислорода от легких к тканям, углекислого газа от тканей к легким. Посредником между клетками и внешней средой является кровь. Она доставляет тканям кислород и уносит от них углекислый газ.
Переход газов из окружающей среды в жидкость и из жидкости в окружающую среду подчиняется определенным физическим закономерностям. Каждый газ переходит в жидкость в зависимости от величины его парциального давления.
Под парциальным давлением понимают ту часть давления, которая приходится на данный газ в смеси газов. При расчете парциального давления газов в альвеолярном воздухе учитывают его насыщенность водяными парами, парциальное давление которых составляет 6,27 кПа (47 мм рт. ст.). В результате на долю остальных газов альвеолярного воздуха приходится 101,3-6,27=95,03 кПа (760-47=713 мм рт. ст.). Зная процентное содержание газов в альвеолярном воздухе, можно рассчитать их парциальное давление. Для кислорода оно будет составлять 13,6 кПа (102 мм рт. ст.), для углекислого газа - 5,33 кПа (40 мм рт. ст.).
Для обозначения давления газов в газовой смеси может быть использован термин "напряжение". Напряжение выражается в миллиметрах ртутного (мм рт. ст.) или водяного (мм вод. ст.) столба.
Движение газов из окружающей среды в жидкость и из жидкости в окружающую среду осуществляется из-за разности их парциального давления. Газ всегда диффундирует из среды, где имеется высокое давление, в среду с меньшим давлением. Это происходит до тех пор, пока не установится динамическое равновесие газов.
Газообмен во всех звеньях дыхательного процесса подчиняется рассмотренным физическим закономерностям.
Проследим движение кислорода из окружающей среды в альвеолярный воздух, затем в капиллярах малого и большого круга кровообращения и к клеткам организма.
Самое высокое парциальное давление кислорода в атмосферном воздухе 21,1 кПа (158 мм рт. ст.), в альвеолярном воздухе 14,4-14,7 кПа (108-110 мм рт. ст.) и в венозной крови, притекающей к легким, 5,33 кПа (40 мм рт. ст.). В артериальной крови капилляров большого круга кровообращения напряжение кислорода составляет 13,6-13,9 кПа (102-104 мм рт. ст.), в межтканевой жидкости - 5,33 кПа (40 мм рт. ст.), в тканях - 2,67 кПа (20 мм рт. ст.) и меньше, в зависимости от функциональной активности клеток.
Таким образом, на всех этапах движения кислорода имеется разность его парциального давления, что способствует диффузии газа.
Движение углекислого газа происходит в противоположном направлении. Самое большое напряжение углекислого газа имеется в тканях, в местах его образования, - 8,00 кПа и более (60 мм рт. ст. и более), в венозной крови 6,13 кПа (46 мм рт. ст.), а альвеолярном воздухе 5,33 кПа (40 мм рт. ст.) и в атмосферном воздухе 0,04 кПа (0,3 мм рт. ст.). Следовательно, разность парциального давления углекислого газа по пути его следования является причиной диффузии газа от тканей в окружающую среду. Схема диффузии газов через стенку альвеол представлена на рис. 25. Однако одними физическими закономерностями объяснить движение газов нельзя. В живом организме равенства парциального давления кислорода и углекислого газа на этапах их движения никогда не наступает. В легких постоянно происходит обмен газов вследствие дыхательных движений грудной клетки, в тканях же разность парциального давления газов поддерживается непрерывным процессом окисления.

Рис. 25. Схема диффузии газов через мембрану альвеолы
Транспорт кислорода кровью. Кислород в крови находится в двух состояниях: физическом растворении и в химической связи с гемоглобином. Из 19 об.% кислорода, извлекаемого из артериальной крови, только 0,3 об.% находятся в растворенном состоянии в плазме, остальная же часть кислорода химически связана с гемоглобином эритроцитов.
Гемоглобин образует с кислородом непрочное, легко диссоциирующее соединение - оксигемоглобин; 1·10-3 кг (1 г) гемоглобина связывает 1,34·10-3 л (1,34 мл) кислорода. Содержание гемоглобина в крови составляет в среднем 140 г/л (14 г%). Отсюда 1·10-1 л (100 мл) крови может связать 1,4·10-2 кг (14 г) × 1,34·10-3 л (1,34 мл) = 1,9·10-2 л (19 мл) кислорода (или 19 об.%), что составляет так называемую кислородную емкость крови. Следовательно, кислородная емкость крови представляет собой максимальное количество кислорода, которое может быть связано 1·10-1 л (100 мл) крови.
Насыщение гемоглобина кислородом колеблется от 96 до 98%. Степень насыщения гемоглобина кислородом и диссоциация оксигемоглобина (образование восстановленного гемоглобина) не находятся в прямой пропорциональной зависимости от напряжения кислорода. Эти два процесса не являются линейными, а совершаются по кривой, которая получила название кривой связывания, или диссоциации, оксигемоглобина.
При нулевом напряжении кислорода оксигемоглобина в крови нет. При низких значениях парциального давления кислорода скорость образования оксигемоглобина невелика. Максимальное количество гемоглобина (45-80%) связывается с кислородом при его напряжении 3,47-6,13 кПа (26-46 мм рт. ст.). Дальнейшее повышение напряжения кислорода приводит к снижению скорости образования оксигемоглобина (рис. 26).
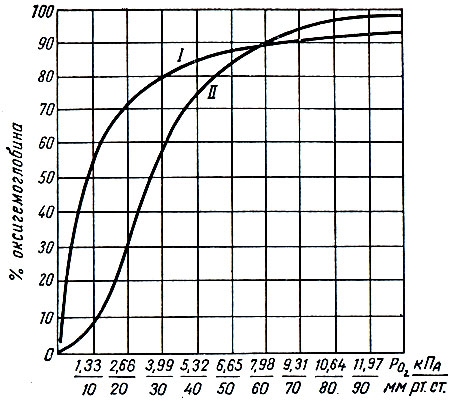
Рис. 26. Кривые диссоциации оксигемоглобина в водном растворе (I) и в крови (II) при напряжении углекислого газа 5,33 кПа (40 мм рт. ст.) (по Баркрофту)
Сродство гемоглобина к кислороду значительно понижается при сдвиге реакции крови в кислую сторону, что наблюдается в тканях и клетках организма вследствие образования углекислого газа. Это свойство гемоглобина имеет важное значение для организма. В капиллярах тканей, где концентрация углекислого газа в крови увеличена, способность гемоглобина удерживать кислород уменьшается, что облегчает его отдачу клеткам. В альвеолах легких, где часть углекислого газа переходит в альвеолярный воздух, способность гемоглобина связывать кислород вновь возрастает.
Переход гемоглобина в оксигемоглобин и из него в восстановленный гемоглобин зависит от температуры. При одном и том же парциальном давлении кислорода в окружающей среде при температуре тела 37-38°С в восстановленную форму переходит наибольшее количество оксигемоглобина.
Таким образом, транспорт кислорода обеспечивается в основном за счет химической связи его с гемоглобином эритроцитов. Насыщение гемоглобина кислородом зависит в первую очередь от парциального давления газа в атмосферном и альвеолярном воздухе. Одной из основных причин, способствующих отдаче кислорода гемоглобином, является сдвиг активной реакции среды в тканях в кислую сторону.
Кровь, проходя по капиллярам большого круга кровообращения, отдает не весь свой кислород. Артериальная кровь содержит около 20 об.% кислорода, венозная - 12 об.%. Следовательно, из 20 об.% ткани получают всего 8 об.%, или 40%, кислорода, содержащегося в крови. Разница в количестве кислорода в артериальной и венозной крови называется артериовенозной разницей. Эта величина характеризует то количество кислорода, которое переходит в ткани из 1·10-1 л (100 мл) крови.
Транспорт углекислого газа кровью. Растворимость углекислого газа в крови гораздо выше, чем растворимость кислорода. Однако только 2,5-3 об.% углекислого газа из общего его количества (55-58 об.%) находится в растворенном состоянии. Большая часть углекислого газа содержится в крови и в эритроцитах в виде солей угольной кислоты (48-51 об.%), около 4-5 об.% - в соединении с гемоглобином, в виде карбгемоглобина, около 2/3 всех соединений углекислого газа находится в плазме и около 1/3 - в эритроцитах.
Угольная кислота образуется в эритроцитах из углекислого газа и воды. И. М. Сеченов впервые высказал мысль о том, что в эритроцитах должен содержаться какой-то фактор типа катализатора, который ускоряет процесс синтеза угольной кислоты. Однако лишь в 1935 г. предположение, высказанное И. М. Сеченовым, было подтверждено. В настоящее время твердо установлено, что в эритроцитах содержится угольная ангидраза (карбоангидраза) - биологический катализатор, фермент, который значительно в (в 300 раз) ускоряет расщепление угольной кислоты в капиллярах легких. В тканевых же капиллярах при участии карбоангидразы происходит синтез угольной кислоты в эритроцитах. Активность карбоангидразы в эритроцитах настолько велика, что синтез угольной кислоты ускоряется в десятки тысяч раз.
Образовавшаяся угольная кислота отнимает основания от восстановленного гемоглобина, в результате чего получаются соли угольной кислоты - бикарбонаты натрия в плазме и бикарбонаты калия в эритроцитах. Кроме того, гемоглобин образует химическое соединение с углекислым газом - карбгемоглобин. Впервые это соединение обнаружено И. М. Сеченовым. Роль карбгемоглобина в транспорте углекислого газа достаточно велика. Около 25-30% углекислого газа, поглащаемого кровью в капиллярах большого круга кровообращения, транспортируется в виде карбгемоглобина. В легких гемоглобин присоединяет кислород и переходит в оксигемоглобин. Оксигемоглобин является более сильной кислотой, чем угольная. Вследствие этого гемоглобин вступает в реакцию с бикарбонатами и вытесняет из них угольную кислоту. Свободная угольная кислота расщепляется карбоангидразой на углекислый газ и воду. Углекислый газ диффундирует через мембрану легочных капилляров и переходит в альвеолярный воздух. Уменьшение напряжения углекислого газа в капиллярах легких способствует расщеплению карбгемоглобина с освобождением углекислого газа.
Таким образом, углекислый газ переносится к легким в форме бикарбонатов и в состоянии химической связи с гемоглобином (карбгемоглобин). Важная роль в сложнейших механизмах транспорта углекислого газа принадлежит карбоангидразе эритроцитов.
Конечной целью дыхания является снабжение всех клеток, органов и тканей кислородом, необходимым для их жизнедеятельности, и удаление из организма углекислого газа. Для осуществления этой цели дыхания необходим ряд условий: 1) нормальная деятельность аппарата внешнего дыхания и достаточная вентиляция легких; 2) нормальный транспорт газов кровью; 3) обеспечение системой кровообращения достаточного кровотока; 4) способность тканей "забирать" из протекающей крови кислород, утилизировать его и отдавать в кровь углекислый газ.
Таким образом, нормальное тканевое дыхание обеспечивается функциональными взаимосвязями между системами дыхания, крови и кровообращения.
При использовании материалов сайта активная ссылка обязательна:
http://anfiz.ru/ 'Анатомия и физиология человека'