Хромосомы и гены
Хромосомы не всегда видны в клеточном ядре, так как они появляются лишь тогда, когда ядра собираются делиться. А деление начинается с ядра. Вот тогда-то, в самом начале деления, рассматривая ядро клетки в микроскоп, можно увидеть прямые или изогнутые палочки. Они хорошо окрашиваются основными красителями. Отсюда и возникло их название (от греческих слов "хромо" - краска, "сома" - тело). В начале деления видно, что каждая хромосома состоит из двух спиралевидных нитей - хроматид. Место соединения обеих хроматид называют центромерой. В хромосоме имеются и другие перетяжки, по расположению и глубине которых можно "установить лицо" каждой отдельной хромосомы. Имеют значение и концевые части хромосом. Длина хромосом у человека равна приблизительно 4-6 мк (рис. 15 и 16).
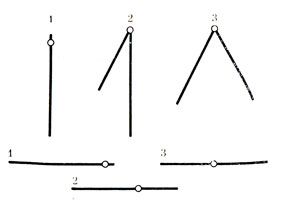
Рис. 15. Различные типы хромосом. 1 - акроцентрические, 2 - субметацентрические, 3 - метацентрические
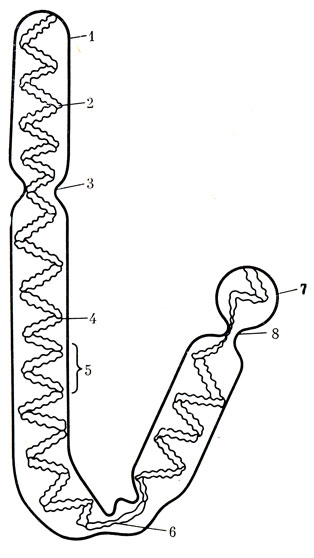
Рис. 16. Схема строения хромосомы. 2 - пеликула, 2 - хроматида, 3 - перетяжка, 4 - малая спираль, 5 - большая спираль, 6 - центромера, 7 - спутник, 8 - вторичная перетяжка
Но сколько же хромосом в клетке? Прежде всего запомним, что половые клетки называют гаметами, а остальные клетки тела - соматическими. В соматической клетке каждая отдельная хромосома имеет своего двойника, ничем от нее не отличающегося. Благодаря наличию таких пар эти клетки содержат двойной или диплоидный набор хромосом, образующийся путем слияния двух половых клеток - гамет (мужской и женской), в которых имеется только одиночный, или гаплоидный, набор хромосом.
Число хромосом в клетках различных организмов сильно колеблется, но для определенного вида строго постоянно. Удивительно, что дольше всего шли споры о том, сколько же хромосом в клетках человека. В конце концов оказалось, что всего 46. Совокупность признаков, характеризующих хромосомный набор клетки, называют кариотипом. Таким образом, в кариотипе человека имеются 22 пары хромосом, одинаковых у мужчин и женщин (рис. 17). Это так называемые аутосомы, которые обычно располагают по размерам и перенумеровывают, причем № 1 дают самой крупной, а № 22 - самой маленькой: 23-я пара - половые хромосомы. Их обозначают символами X и У, причем в клетках тела мужчины только одна X (хромосома, которая образует пару с маленькой У-хромосомой). А у женщин имеются только две одинаковые крупные Х-хромосомы. Как же происходит размножение - деление - клеток? У соматических и половых клеток по-разному. Процесс деления соматических клеток называют митозом, а половых - мейозом.
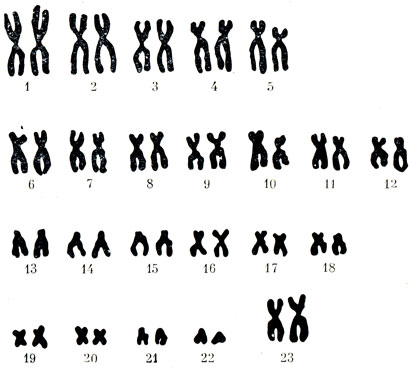
Рис. 17. Нормальный хромосомный набор женщины (по А. А. Прокофьевой-Бельговской)
При митозе из одной материнской клетки образуются две дочерние (рис. 18). Это происходит потому, что каждая из хромосом разделяется во всю свою длину на две совершенно одинаковые половинки. Другими словами, из одной хромосомы образуются две совершенно такие же хромосомы... и тотчас, как бы ссорясь друг с другом, расходятся в противоположные стороны ядра. Этим расхождением как бы управляют центромеры. Затем делится на равные половинки ядро и сама клетка. Так материнская клетка дает жизнь двум дочерним, в каждой из которых сохраняется парное число хромосом. Интересно, что, как только митоз заканчивается, хромосомы снова становятся невидимыми под микроскопом.
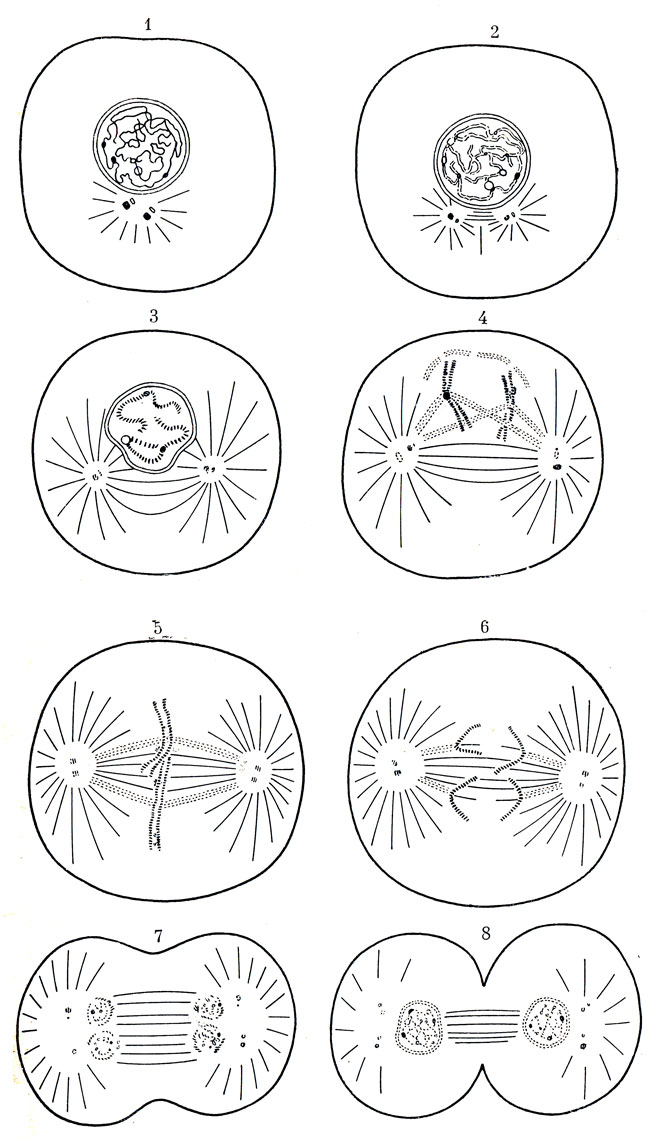
Рис. 18. Схема последовательности стадий митоза (по Мэзия). 1 - период между двумя делениями - интерфаза; хромосомы имеют вид длинных тонких нитей; каждая из двух родительских центриолей находится в паре с маленькой дочерней центриолью; 2 - в какой-то момент между двумя делениями хромосомы удваиваются, центриоли начинают расходиться и начинается образование веретена; 3-4 - профаза. Хромосомы свертываются в спираль и сильно уплотняются; ядерная оболочка и ядрышко распадаются и образуют полюсы, к которым будут перемещаться хромосомы; в то же время образуются связи между центромерами и полюсами; 5 - метафаза; хромосомы располагаются по экватору; 6 - анафаза; после расщепления сестринские хромосомы направляются к полюсам; 7-8 - телофаза; хромосомы раскручиваются, образуются ядерные оболочки и ядрышки дочерних клеток. Каждая центриоль дает новую центриоль. После окончания деления обе клетки вступают в интерфазу (1)
А вот мейоз (процесс образования половых клеток) протекает иначе (рис. 19). Ведь такие клетки в зрелом состоянии содержат только одиночный набор хромосом. Другими словами, яйцеклетка и сперматозоид человека содержат только по 23 хромосомы. При мейозе происходит два последовательных деления. Исходная материнская клетка при первом делении образует две дочерние клетки, каждая из которых получает всего 23 хромосомы. Затем наступает второе деление: каждая из дочерних клеток образует всего четыре дочерних клетки, содержащие по 23 хромосомы. Когда яйцеклетка и сперматозоид сливаются, число хромосом во вновь образованной клетке - зиготе - становится равным 46. Другими словами, половина хромосом получается от матери, а другая половина от отца... со всеми вытекающими из этого последствиями.
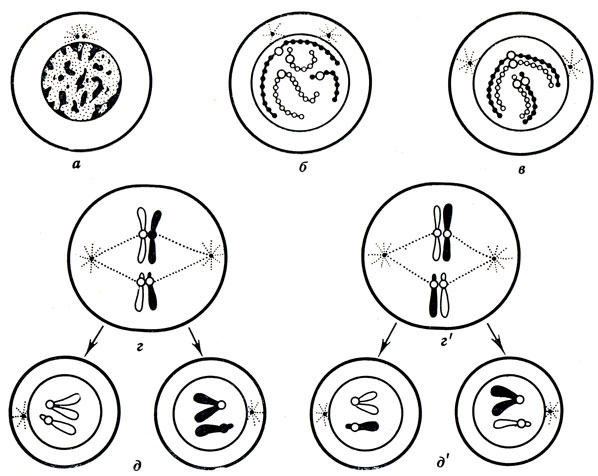
Рис. 19. Схема редукционного деления - мейоза (по А. Мюнтцингу). Число хромосом уменьшается вдвое, и в половые клетки попадает лишь по одной хромосоме из каждой пары: а - покоящееся ядро; б - в ядре видны отдельные нити - хромосомы; в - коньюгация, образование пар, состоящих из двух хромосом каждая (одна хромосома белая, другая - черная, т. е. одна - от матери, другая - от отца); г и г' - метафаза мейоза: ядерная оболочка растворилась, а пары хромосом расположились в экваториальной плоскости клетки; пары хромосом могут быть по-разному расположены по отношению друг к другу, так что образующиеся в следующую фазу ядра получат одну-две черные хромосомы или по одной белой и по одной черной хромосоме; д и д' - пары разъединяются и направляются к разным полюсам клетки, каждая дочерняя клетка получает по одной хромосоме от каждой пары
Ну, а теперь переведем на язык химических понятий то, что здесь происходит. Ведь сущность наследственности заключается в передаче последующим поколениям контроля за видовыми особенностями биохимических реакций. И прежде всего мы встретимся со словом "ген" (от греческого слова "генос" - рождение, происхождение).
Удивительное дело. Обычно химические соединения сначала открывают, и затем дают им название. С геном произошло обратное. Ему сначала дали название и значительно позже дополнили это название химическим содержанием.
Слово "ген" родилось на заре нашего века. Хромосомы были уже известны. Но наследственные свойства организма так многообразны, что все разнообразие этих признаков нельзя было объяснить лишь комбинацией самих хромосом. Возникла нужда в таких мелких единицах наследственности, на которые можно было бы возложить ответственность за развитие того или иного признака. Создалось такое положение, что единицу наследственности - ген - пришлось выдумать. Не всякий верит выдуманному. До недавнего времени были еще биологи, не только сомневавшиеся в существовании гена, но и активно боровшиеся с таким допущением. Они оказались неправы. В этой борьбе победил ген. Победил потому, что природа его была расшифрована до конца. Ему дали конкретное химическое содержание, и в наши дни ученые смогли выделить и синтезировать отдельный ген.
Но что же это такое ген (его называют также "цистрон")? Это участок ДНК, который несет информацию о строении какого-либо конкретного белка. (Но, увы, может, хотя и редко, допускать "опечатки" в этой информации.)
Еще в начале нашего века английский ученый Гэр-род открыл врожденные нарушения обмена и назвал их ошибками природы. Он же предположил, что причиной таких заболеваний может быть наследственно передаваемый недостаток какого-либо фермента. Подобного рода наблюдения умножились, и уже лет 30 назад американские биологи Бидл и Татум предложили гипотезу: "один ген - один фермент". В настоящее время эта формула стала, пожалуй, слишком узкой. Поле деятельности генов распространяется не только на белки-ферменты. Как было сказано выше, белковая молекула может состоять из различных полипептидных цепей. В этом случае строительством каждой цепи может заниматься отдельный ген. Такие гены называют структурными. И на сегодня правильнее пользоваться формулой: "один ген - один полш-пептид".
Итак, ген - мельчайшая материальная единица наследственности? Нет, ученые на этом не остановились. Имеются основания для допущения, что и гены могут делиться на еще более мелкие единицы, на сотни таких единиц, которым уже дали название "мутоны".
Теперь вспомним о том, что генетическая информация в нуклеиновой кислоте записана путем определенной последовательности оснований (аденина, гуанина, урацила, цитозина), которые называют нуклеотидами. Каждому нуклеотиду в одной цепи ДНК всегда строго соответствует определенный нуклеотид в другой цепи двойной спирали ДНК: они даже слегка связаны водородными мостиками друг с другом. Так, цитозину в определенном участке одной цепи всегда противостоит, как бы дополняя его, гуанин в другой цепи спирали. Такую дополнительность называют комплементарностью.
Комплементарность имеет особое значение. Благодаря открытию этого правила раскрыта сущность важнейшего свойства ДНК - ее способность к редупликации, т. е. к построению из одной цепи новой двуспиральной молекулы, абсолютно ничем не отличающейся от старой (рис. 20 и 21).
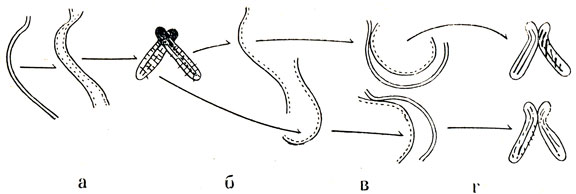
Рис. 20. Репликация хромосомы (по Тейлору). а - удвоение в присутствии Н3-тимидина, б - первая метафаза после включения метки, в - удвоение в отсутствие Н3-тимидина, г - вторая метафаза после включения метки
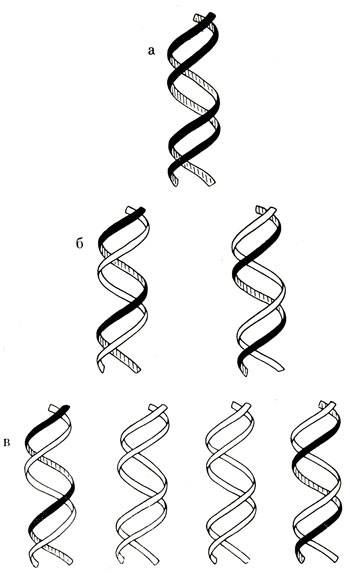
Рис. 21. Модель репликации в ДНК по полуконсервативному механизму. а - исходная родительская молекула, б - первое поколение дочерних молекул, в - второе поколение
Принцип комплементарности соблюдается и тогда, когда ДНК строит другую нуклеиновую кислоту - информационную (и-PHK)*. При этом и-PHK как бы занимается перепиской (транскрипцией) данных о первичной структуре ДНК (к сожалению, вместе с возможными "опечатками" в ее записи).
*(Отметим, что в этом строительстве используются другие нуклеотиды и его ускоряет полимераза РНК (не полимераза ДНК, которую уже выделил американский ученый А. Корнберг).)
Теперь несколько детальнее остановимся на том, как происходит запись информации на молекуле ДНК и как эта информация переносится от гена к белку. Другими словами, как клетка строит по строго заданному плану именно те белки, в первую очередь белки-ферменты, которые ей нужны (рис. 22).
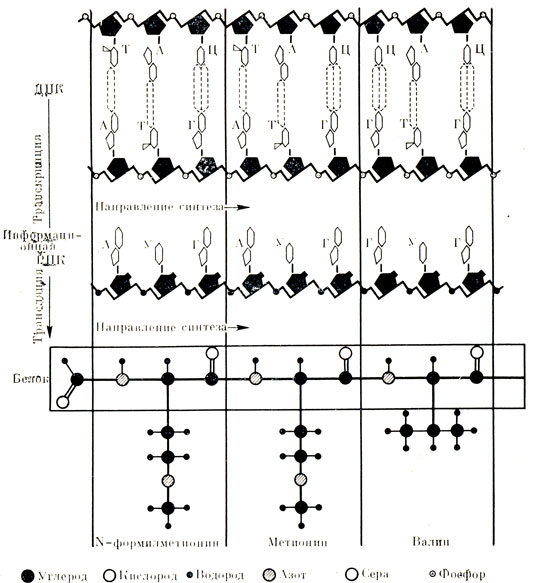
Рис. 22. Передача генетической информации. На первом этапе информация о данном белке, закодированная в линейной последовательности нуклеотидов, переносится с ДНК на РНК. Этот процесс называют транскрипцией. Кодовыми буквами в ДНК служат четыре основания: аденин (А), тимин (Т), гуанин (Г) и цитозин (Ц). Водородные связи (пунктирные линии) между комплементарными основаниями (А - Т и Г - Ц) удерживают две цепи ДНК вместе. Обе цепи состоят из чередующихся звеньев дезоксирибозы (черные пятиугольники) и фосфата, РO3Н. Кодовые буквы в информационной РНК те же, что и в ДНК, с той только разницей, что вместо тимина в РНК стоит урацил (У). Цепь РНК представляет собой точную копию одной из цепей ДНК (с учетом только что упомянутого различия); кроме того, в РНК вместо дезоксирибозы присутствует рибоза. На втором этапе происходит перевод с языка информационной РНК на язык белка - этот процесс называют трансляцией. Кодовые буквы РНК считываются по триплетам кодонам, каждый из которых определяет одну (а иногда и более) аминокислоту, образующую молекулу белка
Химическими кирпичиками, из которых построена молекула белков, являются известные нам аминокислоты. Аминокислотный состав белка и последовательность расположения аминокислотных остатков в полипептидной цепи определяют свойства белка. Нуклеиновая кислота пользуется, как мы уже упоминали выше, кодом, заключающимся в том, что каждую синтезируемую аминокислоту определяют четыре нуклеотида, взятые по три (триплеты или кодоны). Каким образом? Опять-таки последовательностью расположения этих нуклеотидов в самом триплете. А сколько существует таких триплетов? Это легко выяснить. Мы можем сочетать различным образом четыре нуклеотиды в триплете всего 64 раза. Но ведь аминокислот всего 20. Неужели природа проявляет здесь бессмысленную расточительность? Вероятно, нет. Происходит только своеобразная подстраховка, и каждая аминокислота кодируется не одним, а двумя-тремя триплетами (рис. 23). "Безработных" триплетов, очевидно, нет. Они последовательно расположены вдоль цепи нуклеиновой кислоты (рис. 24). Уже сделаны попытки расшифровать генетический код аминокислот для бактерий и вирусов:

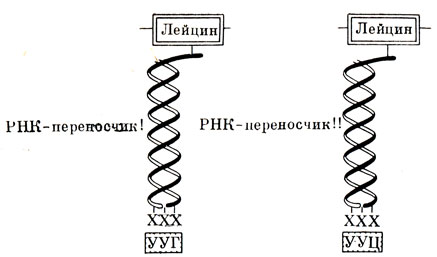
Рис. 23. Двойное кодирование аминокислоты лейцина двумя триплетами УУГ и УУЦ. Одна РНК - переносчик устанавливает лейцин против триплета УУГ, другая - против УУЦ
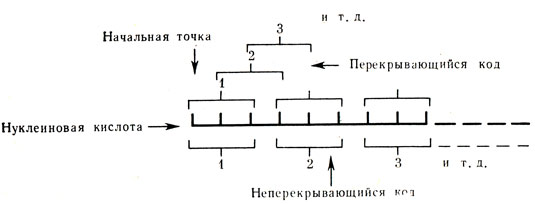
Рис. 24. Перекрывающийся и неперекрывающийся коды. Короткими вертикальными линиями изображены основания нуклеиновых кислот. Схема основана на представлении о триплетном коде (по Крик, Барнетт, Бреннер и Уотте - Тобин)
Можно не сомневаться, что аминокислотный состав синтезируемого белка зависит от последовательности нуклеотидов в молекуле нуклеиновой кислоты, хотя для многоклеточных организмов это еще надо доказать. Вообще не раз отмечалось, что под влиянием различных воздействий (понижения температуры, изменений концентраций некоторых металлов и др.) нормальный механизм кодирования может нарушаться.
Синтез белка требует энергетического обеспечения. Поставщиками энергии в клетке являются соединения фосфорной кислоты, о которых будет сказано в соответствующей главе этой книги. В синтезе белка принимает участие богатая энергией аденозинтрифосфорная кислота - АТФ. На соответствующую аминокислоту воздействует специальный активирующий фермент (его называют синтетазой) и АТФ (рис. 25). Аминокислота образует сложное соединение аминоациладенилат и, таким образом, активируется. Этот процесс сопровождается выделением пи|рофосфорной кислоты. Активированную, как бы заряженную энергией аминокислоту тот же активирующий фермент переносит на транспортную РНК. Конечный нуклеотид (аденин) РНК связывает эту аминокислоту содержащимся в нем углеводом - рибозой.
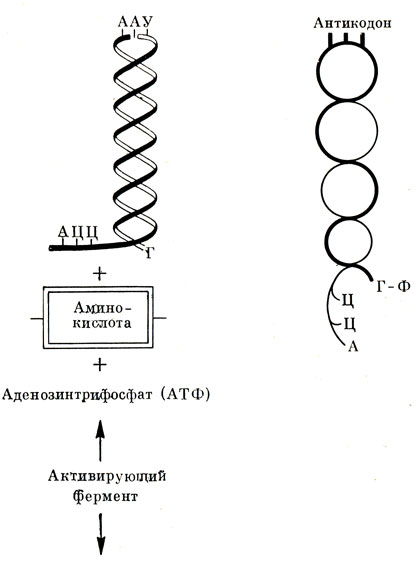
Рис. 25. Структура молекулы РНК-переносчик (т-РНК)
В каждой клетке содержится ровно столько различных транспортных РНК, сколько в ней различных аминокислот (обычно 20). Тут строго соблюдается правило: "Не в свои сани не садись". Каждая активированная аминокислота "узнает" и "садится" только на свою транспортную РНК (а вернее, как бы сцепляется с одним из ее концов). На противоположном конце т-РНК, похожем на петлю, имеется участок, называемый антикодоном. Он состоит из триплета нуклеотидов, различного в разных транспортных РНК. Именно этот участок позволяет аминокислоте "узнать" свою т-РНК (рис. 26). Более того, он заботится и о том, куда поместить аминокислоты в дальнейшем, укладывая их в последовательности, соответствующей расположению нуклеотидов, на своеобразную матрицу, роль которой выполняет находящаяся на рибосомах и-РНК. Это происходит таким образом, что отдельные молекулы т-РНК подходят к рибосоме, к месту синтеза цолипептидной связи, и присоединяются своими антикодонами к соответствующим кодонам и-РНК.
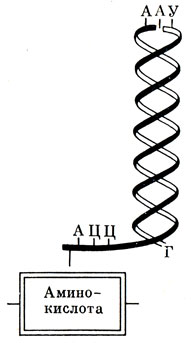
Рис. 26. Схема строения транспортной РНК: Г-Ф - гуанозинфосфат, Ц - цитозин, А - аденин
Как представить себе синтез белка*. Вероятнее всего, передвигаясь вдоль молекулы и-РНК, отдельные рибосомы (вернее, их скопления - полисомы) получают от нее информацию, как бы считывая ее, и последовательно нанизывают одно на другое аминокислотные звенья полипептидной цепи. Эти звенья подаются отдельными молекулами т-РНК, которые присоединяются вдоль и-РНК. Доходя до конца молекулы и-РНК, рибосома оставляет ее и освобождает вновь образующуюся белковую молекулу. В это время к другому концу молекулы и-РНК присоединяется новая рибосомная частица (табл. 1).
*(На синтез одной молекулы белка затрачивается не более одной минуты благодаря тому, что на рибосомах, объединенных в полисому, одновременно идет синтез нескольких молекул белка.)
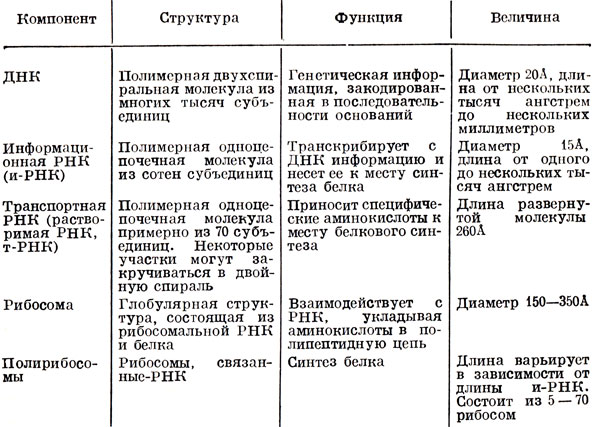
Таблица 1. Клеточные компоненты, участвующие в белковом синтезе
Таким образом структурные гены определяют молекулярное строение белка. Но по современным представлениям существуют еще гены-регуляторы и гены-операторы. Здесь руководит ген-регулятор. Он независим в своих действиях. В роли его помощника выступает особое вещество - репрессор, образующийся в цитоплазме. Репрессор, воздействуя на ген-оператор, может тормозить перенос информации от структурного гена к белку. Значение такой репрессии состоит в том, что она выключает синтез ферментов, потребности в которых в данный момент нет; таким образом сохраняется фонд свободных аминокислот для синтеза других белков (рис. 27).
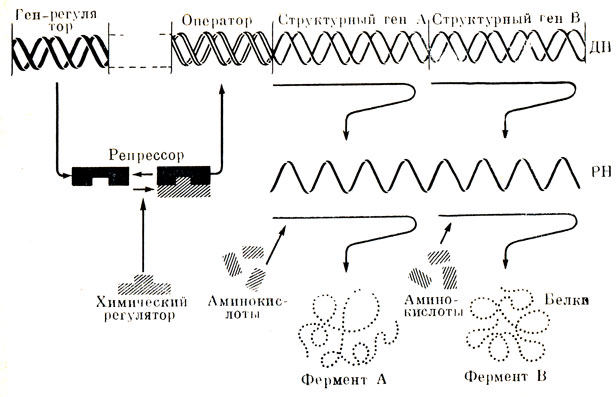
Рис. 27. Схема регуляции белкового синтеза генетическим 'репрессором' (по Жакобу и Moнo). Ген-регулятор контролирует синтез молекулы репрессора, которая связывается с метаболитом, играющим роль регуляторного сигнала. При связывании репрессор либо активируется, либо инактивируется в зависимости от того, какая система имеет место в данном случае - репрессируемая или индуцируемая. В активном состоянии репрессор соединяется с 'геном-оператором', что приводит к выключению структурных генов, под контролем которых протекает синтез данной группы ферментов. Есть указания, что регуляция возможна и на уровне трансляции
Если какое-либо вещество увеличивает скорость синтеза белка-фермента, уже имеющегося в клетке, то это явление называют индукцией, а вещество - индуктором. Один индуктор может вызвать повышение скорости синтеза целой группы ферментов.
Все эти представления нуждаются еще во многих уточнениях. Ясно одно, что только хромосомы могут удваиваться (репликация) при полном сохранении своих специфических особенностей. Другими словами, материнские хромосомы служат шаблоном для дочерних хромосом, которые их копируют. Существует ли внехромосомная передача наследственности? Поскольку хромосомы заключаются в ядре, это равносильно вопросу - может ли цитоплазма передавать наследственность. Ученые до сих пор спорят об этом. Что же заставляет их спорить? Дело в том, что в цитоплазме, и в частности в митохондриях - силовых станциях цитоплазмы, открыли присутствие нуклеиновых кислот. Откуда они взялись и что они там делают? Видный французский ученый Жакоб придерживается мнения, что когда-то, на заре становления жизни клетки, в нее проникли бактерии (содержащие нуклеиновые кислоты). Хозяин и пришельцы "сошлись характерами", стали работать вместе со взаимной выгодой. Возможно, что нуклеиновая кислота играет особую, еще не изученную роль в митохондрии. Во всяком случае, на долю таких кислот приходится менее 1%. А остальные 99% нуклеиновых кислот сосредоточены в ядре, содержащем хромосомы. Следовательно, хромосомная передача наследственности - основной (если не единственный) путь передачи наследственности.
Как будто бы все ясно. И все же это можно сказать только об основных принципах, так как многое в процессе перехода от нуклеиновых кислот к белкам нуждается в изучении.
Не надо забывать, что почти все исследования проводились на бактериях. И если принять, что такие бактерии содержат "всего" две-три тысячи белков, то это соответствует 10 миллионам знаков генетической информации. Предположим, что нам полностью известны правила распределения этих знаков (а это далеко не так). Что дает это для познания генетического шифра человека, содержащего минимум 10 миллиардов знаков! А ведь сотни триллионов клеток нашего тела еще должны согласовывать свои действия с тем, что происходит вокруг них. Как разгадать пути наследственности хотя бы среди 15 миллиардов клеток человеческого мозга, между которыми существуют чрезвычайно сложные взаимоотношения? Как расшифровать возможное кодирование клеток, помогающее им узнавать друг друга, чтобы вступать во взаимодействия? Как организм человека накапливает информацию и использует прошлый опыт? Что обеспечивает постоянство строения организма и его функций (что можно назвать генетической системой памяти)? Каким образом опыт человека и особенности его развития закрепляются в нервной системе через наследственность (нервная система памяти)*? Все это проблемы будущего молекулярной биологии. Об этом хорошо сказано в поэме Д. Апдайка: "Но может кость создать из коллагена одна природа мудрая пока"**.
*(Как известно, нервная система играет огромную роль в жизнедеятельности нашего организма. Но в химической структуре работы нервных клеток пока еще много белых пятен. Некоторые исследователи предполагают, что ДНК ядра этих клеток неполновластный распорядитель структуры части рибонуклеиновых кислот, которую определяют сигналы возбуждения, поступающие извне и запоминающиеся нервной клеткой. К рассмотрению химической основы памяти мы еще вернемся в дальнейшем. К сожалению, веских доказательств правильности подобных предположений пока не имеется.)
**(Нельзя не отметить, что биология с каждым годом накапливает факты, оправдывающие перенос данных, полученных в опытах с микробами, на многоклеточные организмы.)
При использовании материалов сайта активная ссылка обязательна:
http://anfiz.ru/ 'Анатомия и физиология человека'