Проекционные волокна
Проекционные волокна являются восходящими и обратными связями между корой головного мозга, базальными ядрами с нижележащими отделами центральной нервной системы (ствол, мозжечок, спинной мозг). Принимают участие в формировании сложных рефлекторных дуг, которые складываются из чувствительных и двигательного звеньев, представляющих функционально единое целое. Их контакт совершается только в коре головного мозга в так называемых ассоциативных корковых полях. В этих полях происходит затем переключение на моторную зону мозга, где двигательные импульсы распространяются к мышцам и железам. Результатом всей нервной активности является поведение, которое может осуществляться только при посредстве двигательной системы. Чувствительные системы возникли в процессе эволюции для обеспечения лучшей регуляции двигательной системы. Таким образом, чувствительное (афферентное) звено и двигательное (эффекторное) звено - единая цепь нейронов. Для удобства изучения излагаются отдельно чувствительные - восходящие (афферентные) и двигательные - нисходящие (эфферентные) пути.
Восходящие пути разделяются на: 1) экстероцептивные, начинающиеся от кожи и специализированных органов чувств; 2) проприоцептивные, начинающиеся от рецепторов надкостницы, капсул, связок суставов, мышц; 3) интероцептивные, начинающиеся от рецепторов внутренних органов и стенок кровеносных сосудов. Последние излагаются в разделе "Вегетативный отдел нервной системы".
Обычно говорят о пяти чувствах человека: зрение, слух, осязание, вкус и обоняние. В действительности это не совсем верно. У человека и животных имеется шестое чувство - чувство равновесия, включение которого совершается без привлечения к этому сознания. Чувство равновесия содержит две составляющие: в голове имеется орган, перерабатывающий прямолинейные ускорения и вращательные ускорения. Вращательные ускорения функционируют во всех трех измерениях и рефлекторно возникают соответствующие противодействующие движения.
Необходимы некоторые разъяснения и об осязании. Осязание, т. е. кожное "ощущение" (механорецепция), в действительности складывается из: 1) осязания (тактильное чувство), которое в силу быстрой адаптации реагирует только на первые моменты соприкосновения; 2) чувства давления; 3) чувства перемещения. Вторым обобщенным аналогичным понятием ощущений следует различать в системе экстерорецепции терморецепцию, имеющую специфические рецепторы чувства холода и тепла. У некоторых животных, например у гадюки, существуют специальные локальные рецепторы в области головы для принятия инфракрасного излучения. У человека тепловые рецепторы разбросаны по всей поверхности кожи.
Важным ощущением является ноцицепция, или болевое ощущение, возникающее при раздражении нервных окончаний в таламусе.
I. Экстероцептивные пути
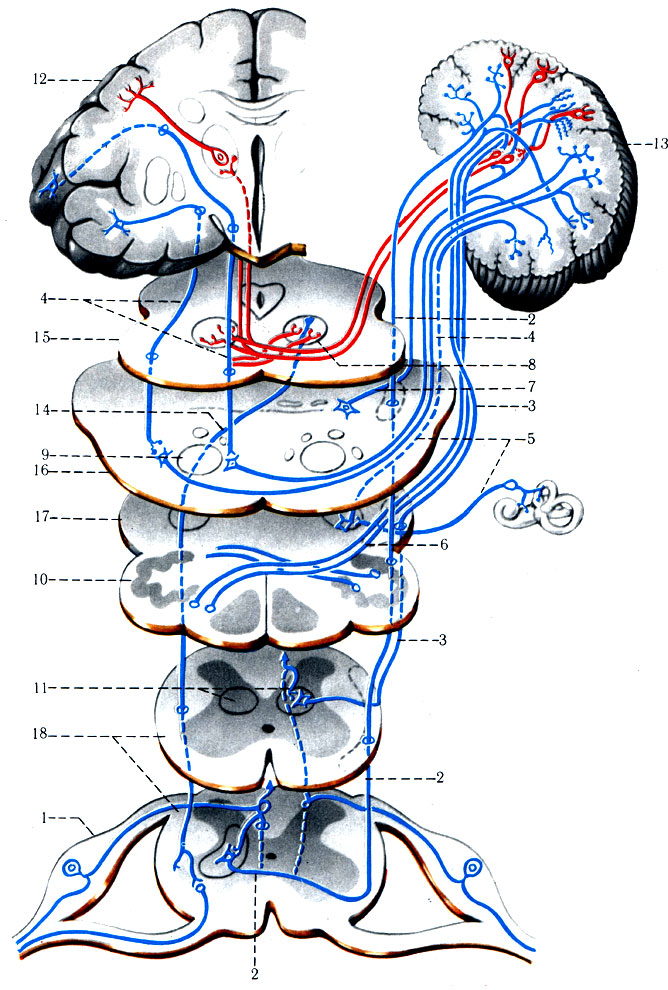
1. Спиноталамический путь разделяется на латеральный и передний (tr. spinothalamici lateralis et anterior) (рис. 496). Содержит волокна, проводящие импульсы от кожных рецепторов, которые воспринимают тактильную, температурную и болевую чувствительность всех частей тела, кроме кожи и органов лицевого отдела головы. От кожи лица подобные раздражения воспринимаются рецепторами тройничного нерва. Первый нейрон спиноталамического пути униполярной формы, располагается в межпозвоночном узле. Его дендриты имеют свободные нервные окончания (воспринимают боль), тельца Краузе (воспринимают холод), тельца Руффини (воспринимают тепло). Аксоны в составе задних чувствительных корешков достигают клеток ядер заднего столба спинного мозга. Аксоны нейронов задних столбов через переднюю спайку проникают на противоположную сторону спинного мозга, поднимаясь на 2-3 сегмента, затем образуют пучок в передней части бокового канатика (передний спиноталамический путь), имея восходящее направление. В продолговатом мозге пучки волокон располагаются дорсальнее оливы, затем проникают в дорсальную часть моста, располагаясь рядом с медиальной петлей кнаружи от красных ядер. Этот нейрон заканчивается синапсом в переднелатеральном ядре таламуса, вступая в синаптическую связь с клетками III нейрона таламокортикального пути (tr. thalamocorticalis). Их отростки проникают через среднюю часть задней ножки внутренней капсулы, направляясь к задней центральной извилине (поля 1-2-3) и верхней теменной дольке (поля 5-7).
Передний спиноталамический путь в основном проводит импульсы от тактильных рецепторов (тельца Мейсснера и Меркеля). Чувство давления воспринимают пластинчатые тельца (Фатера - Пачини).
Центральные отростки I нейрона от клеток спинномозгового узла образуют латеральную группу волокон (латеральный спиноталамический путь), вступающих в спинной мозг. От места вхождения заднего корешка латеральная группа волокон имеет восходящие и нисходящие ветви на 1-2 сегмента (рис. 496). Волокна от ядер задних столбов спинного мозга переходят через переднюю спайку на противоположную сторону в передний канатик белого вещества; только в среднем мозге они присоединяются к волокнам латерального спиноталамического пути, достигая с ним таламуса и постцентральной извилины (корковый центр).
2. Чувствительный путь тройничного нерва начинается от тактильных, болевых и температурных рецепторов кожи лица и головы (кроме затылочной области), органов глазницы и ротовой полости. Импульсы направляются в узел тройничного нерва (гомолог спинномозгового узла) по его дендритам (рис. 496). Аксоны узла образуют чувствительный корешок тройничного нерва, который достигает дорсальной части моста, где разделяется на восходящую и нисходящую ветви. Восходящую ветвь образуют довольно толстые волокна, по которым распространяются импульсы от тактильных и проприоцептивных рецепторов. Тонкие волокна нисходящей ветви проводят к ядру спинномозгового пути тройничного нерва импульсы болевой и температурной чувствительности в соответствующем ядре тройничного нерва (nucl. sensorius n. trigemini). От клеток этого ядра возникают волокна II нейрона тройничного нерва. Они образуют перекрест в области моста, а затем присоединяются к пучку медиальной петли, вместе с которой заканчиваются в ядрах таламуса. Аксоны клеток переднего ядра таламуса проходят в задней трети задней ножки внутренней капсулы и направляются к коре нижйего участка постцентральной извилины. Нисходящие волокна чувствительного корешка тройничного нерва достигают спинного мозга и образуют крупный по объему спинномозговой путь тройничного нерва, в составе которого имеется большое количество тонких безмиелиновых волокон, проводящих болевую и температурную чувствительность. Этот пучок спускается вниз по спинному мозгу до IV шейного сегмента, заканчиваясь в желатинозной субстанции, которая является чувствительным ядром этого нерва (nucl. tr. spinalis n. trigemini). При этом волокна от рецепторов кожи, находящихся выше оральных отделов лица, заканчиваются в самых нижних отделах ядра, а от нижележащей части лица - в верхнем его отделе.
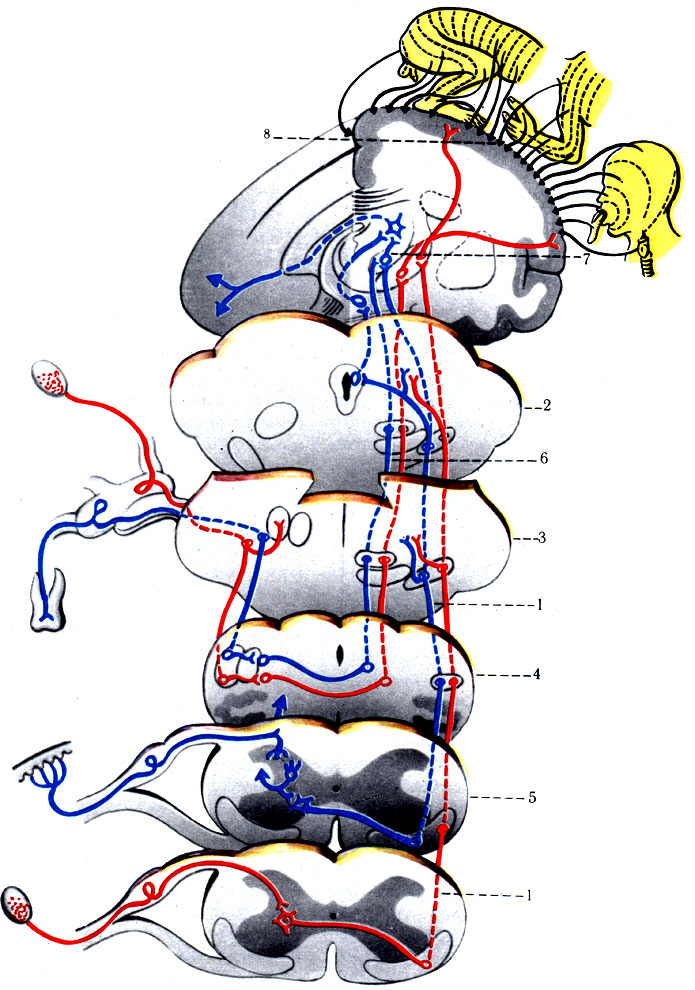
496. Схема проводящих путей общей чувствительности и тройничного нерва (по Сентаготаи). 1 - tr. spinothalamicus anterior; 2 - средний мозг; 3 - мост; 4 - продолговатый мозг; 5 - спинной мозг; 6 - tr. spinalis n. trigemini; 7 - таламус; 8 - gyrus postcentral
Вторыми нейронами пути болевой и температурной чувствительности являются клетки указанного ядра. Они совершают перекрест в мосту, продолговатом и спинном мозге, проходят в ретикулярной формации продолговатого мозга и моста, где, вероятно, имеют связи с ядрами, затем на противоположной стороне вступают также в состав медиальной петли, вместе с которой заканчиваются в переднем ядре таламуса. Третий нейрон этого пути располагается совместно с волокнами таламокортикального пути и достигает задней центральной извилины.
3. Проводящие пути обонятельного анализатора (tr. olfactorius) (рис. 497) имеют сложное строение и излагаются несколько схематизированно. Обонятельные рецепторы слизистой оболочки носовой полости воспринимают изменения химизма воздушной среды и являются самыми чувствительными по сравнению с рецепторами других органов чувств.
Первый нейрон образован биполярными клетками, расположенными в слизистой оболочке верхней носовой раковины и носовой перегородки. Дендриты обонятельных клеток имеют булавовидные утолщения с многочисленными ресничками, воспринимающими химические вещества воздуха; аксоны соединяются в обонятельные нити (fila olfactoria), проникающие через отверстия решетчатой пластинки в полость черепа, и переключаются в обонятельных клубочках обонятельной луковицы (bulbus ofactorius) на II нейрон. Аксоны II нейрона (митральные клетки) формируют обонятельный тракт (tr. ofactorius) и заканчиваются в обонятельном треугольнике (trigonum olfactorium) и в переднем продырявленном веществе (substantia perforata anterior), где находятся клетки III нейрона. Аксоны III нейрона группируются в три пучка - наружный, промежуточный и медиальный, которые направляются к различным структурам мозга. Наружный пучок, обогнув sulcus lateralis большого мозга, достигает коркового центра обоняния, расположенного в крючке (uncus) височной доли. Промежуточный пучок, проходя в гипоталамической области, заканчивается в сосцевидных телах и в среднем мозге (красное ядро). Медиальный пучок разделяется на две части: одна часть волокон, пройдя через gyrus paraterminalis, огибает corpus callosum, вступает в gyrus fornicatus, достигает гиппокампа и крючка; другая часть медиального пучка образует обонятельно-поводковый пучок нервных волокон, проходящих в stria medullaris таламуса своей же стороны. Обонятельно-поводковый пучок заканчивается в ядрах треугольника уздечки надталамической области, где начинается нисходящий путь, подключающий мотонейроны спинного мозга. Ядра треугольной уздечки дублируваны второй системой волокон, приходящих из сосцевидных тел (рис. 497).
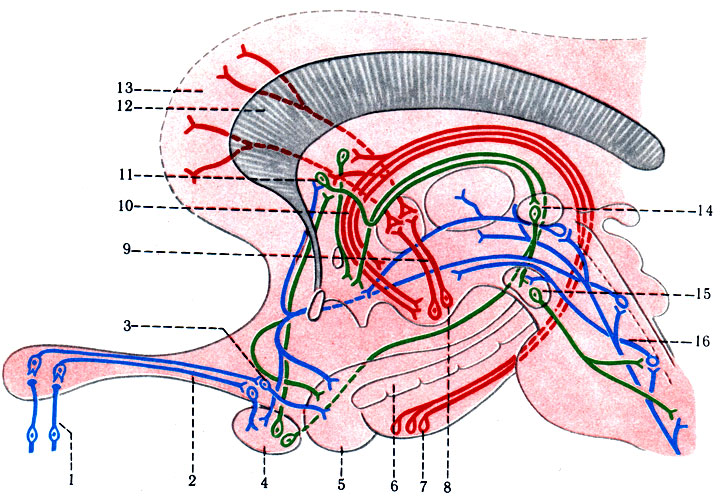
497. Схема проводящего пути обонятельного анализатора. 1 - обонятельные нити; 2 - обонятельный тракт; 3 - обонятельный треугольник; 4 - миндалевидное ядро; 5 - крючок; 6 - зубчатая извилина; 7 - гиппокамп; 8 - сосцевидное тело; 9 - сосцевидно-зрительный пучок; 10 - пути свода; 11 - ядро прозрачной перегородки; 12 - мозолистое тело; 13 - поясная извилина; 14 - ядро поводка; 15 - межножковое ядро; 16 - tr. spinothalamicus
4. Проводящий путь зрительного анализатора (tr. opticus) (рис. 498). Зрительный анализатор осуществляет восприятие, передачу, синтез и анализ световых раздражений, воспринимаемых светочувствительными клетками (палочки и колбочки) со скоростью около 720 м/с. Световые лучи проникают через роговицу, переднюю и заднюю камеры глаза, хрусталик, стекловидное тело на сетчатку. Под действием света родопсин палочек и колбочек распадается, в результате чего образуется энергия, которая воспринимается рецепторами I нейрона, представленного в сетчатке биполярными клетками. Биполярные клетки контактируют с ганглиозными клетками, являющимися II нейроном (рис. 498А). Аксоны ганглиозных клеток радиально собираются к заднему полюсу глаза, образуя зрительный нерв (n. opticus), который выходит из глазницы через зрительное отверстие и направляется к основанию головного мозга. Зрительный нерв состоит из четырех видов волокон: 1) зрительных, начинающихся от височной половины сетчатки; 2) зрительных, идущих от носовой половины сетчатки; 3) папилломакулярных, исходящих из области желтого пятна; 4) световых, идущих в супраоптическое ядро гипоталамуса. Около серого бугра (tuber cinereum) волокна зрительного нерва образуют частичный перекрест (chiasma opticum) за счет медиальных половин. После перекреста формируется зрительный тракт, который, обогнув снаружи ножку мозга, заканчивается в наружном коленчатом теле (corpus geniculatum laterale), в верхнем двухолмии (colliculus superior), а небольшая часть волокон - в подушке таламуса (pulvinar). Верхнее двухолмие является реф-лекторным центром для выполнения автоматических движений, возникающих при включении мотонейронов спинного мозга. Аксоны верхнего двухолмия передают импульсы к парасимпатическим и двигательным ядрам III пары головных нервов (рис. 499), а также включают центры симпатической иннервации спинного мозга, иннервирующие гладкую мускулатуру глаза. Наружное коленчатое тело состоит из семи слоев клеток, чередующихся с прослойками белого вещества. Аксоны III нейрона, лежащие в коленчатом теле, образуют зрительное сияние, волокна которого, обогнув нижний и задний рога бокового желудочка, достигают коры затылочной области, лежащей по бокам шпорной борозды (поля 17, 18, 19).
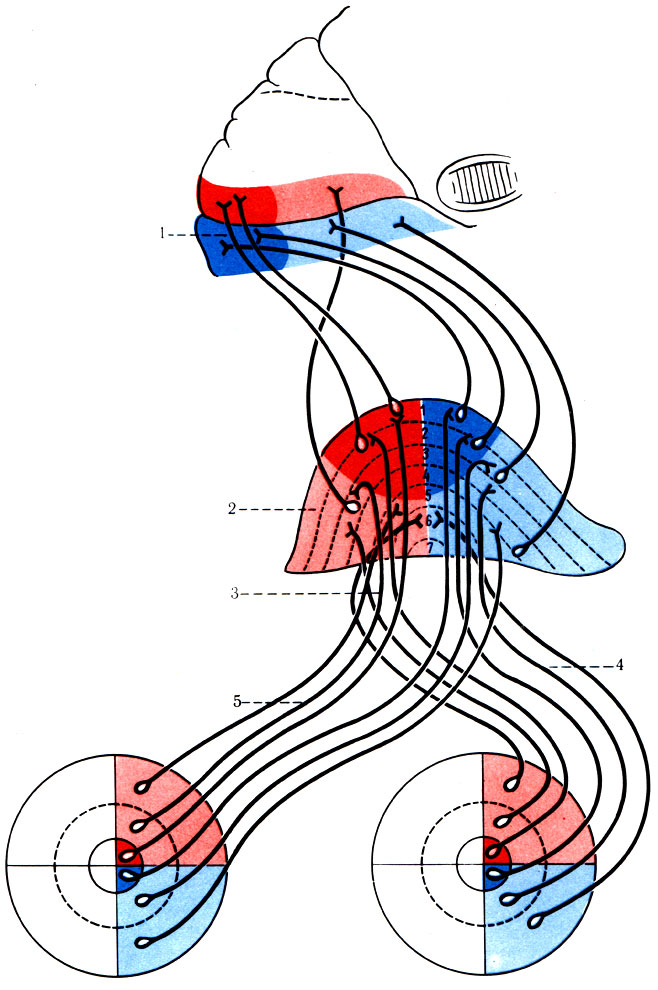
498. Схема проводящего пути зрительного анализатора (левая половина) и соответствие зон сетчатки и зрительного центра (по Сентаготаи). 1 - зрительный корковый центр; 2- латеральное коленчатое тело с ядерными слоями (1-7); 3 - зрительный тракт; 4 - латеральная часть зрительного нерва; 5 - медиальная часть зрительного нерва
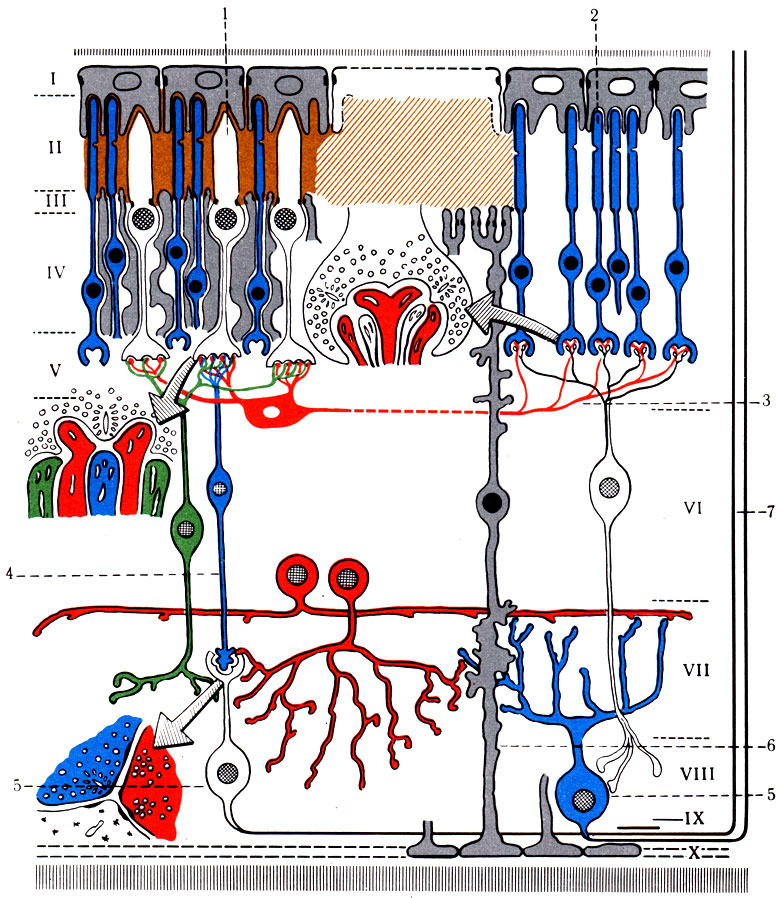
498А. Схема строения и связей сетчатки глаза (по Boycott, Dowling). I-X - слои сетчатки: 1 - наружные членики колбочек; 2- палочки; 3 - горизонтальная клетка; 4 - биполярная клетка; 5 - ганглионарная клетка;. 6 - пигментная клетка; 7- волокна зрительного нерва
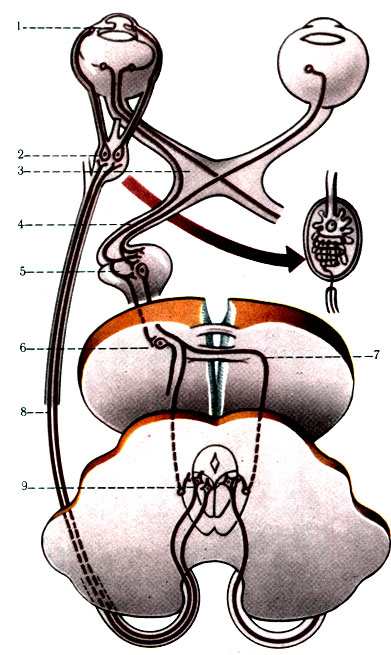
499. Схема проводящего пути зрачкового рефлекса (по Сентаго-таи). 1 - m. sphincter pupillae; 2 - gangl. ciliare; 3 - n. opticus; 4 - tr. noptici; 5 - corpus geniculatum laterale; 6 - ядро двухолмия; 7 - commissura cerebri posterior; 8 - n. oculomotorius; 9 - ядро III пары
Корковый зрительный центр имеет весьма сложную многогранную систему нейронных связей. В ней находятся нейроны, реагирующие только на начало и конец освещения. В зрительном центре совершается не только обработка информации по ограничительным линиям, яркостям и градациям цвета, но и оценка направления движений объекта. В соответствии с этим и число клеток в коре головного мозга больше в 10000 раз, чем в сетчатке. Существенная разница имеется между числом клеточных элементов наружного коленчатого тела и зрительным центром. Один нейрон наружного коленчатого тела соединен с 1000 нейронов зрительного коркового центра, а каждый из этих нейронов в свою очередь образует синаптические контакты с 1000 соседних нейронов.
5. Проводящий путь слухового анализатора (tr. n. cochlearis) (рис. 500). Слуховой анализатор осуществляет восприятие звуков, их анализ и синтез. Первый нейрон находится в спиральном узле (gangl. spirale), расположенном в основании полого улиткового веретена. Дендриты чувствительных клеток спирального узла проходят по каналам костной спиральной пластинки к спиральному органу и оканчиваются у наружных волосковых клеток. Аксоны спирального узла составляют слуховой нерв, вступающий в области мостомозжечкового угла в ствол мозга, где и заканчиваются синапсами с клетками дорсального (nucl. dorsalis) и вентрального (nucl. ventralis) ядер.
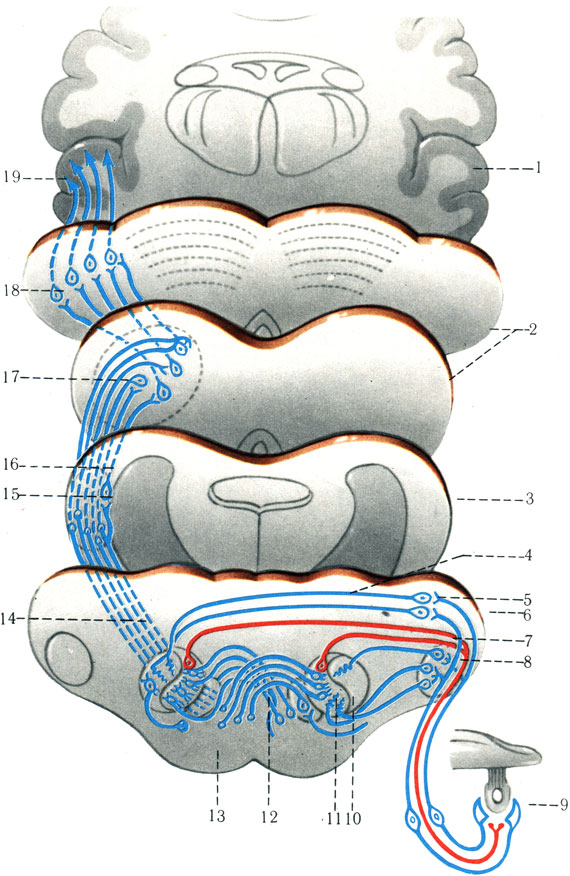
500. Схема пути слухового анализатора (по Сентаготаи). 1 - височная доля; 2 - средний мозг; 3 - перешеек ромбовидного мозга; 4 - продолговатый мозг; 5 - улитка; 6 - вентральное слуховое ядро; 7 - дорсальное слуховое ядро; 8 - слуховые полоски; 9 - оливо-слуховые волокна; 10 - верхняя олива; 11 - ядра трапециевидного тела; 12 - трапециевидное тело; 13 - пирамида; 14 - латеральная петля; 15 - ядро латеральной петли; 16 - треугольник латеральной петли; 17 - нижнее двухолмие; 18 - латеральное коленчатое тело; 19 - корковый центр слуха
Аксоны II нейронов от клеток дорсального ядра образуют мозго-вые полоски (striae medullares ventriculi quarti), находящиеся в ромбовидной ямке на границе моста и продолговатого мозга. Большая часть мозговой полоски переходит на противоположную сторону и около средней линии погружается в вещество мозга, подключаясь к латеральной петле (lemniscus lateralis); меньшая часть мозговой полоски присоединяется к латеральной петле своей же стороны.
Аксоны II нейронов от клеток вентрального ядра участвуют в образовании трапециевидного тела (corpus trapezoideum). Большая часть аксонов переходит на противоположную сторону, переключаясь в верхней оливе и ядрах трапециевидного тела. Другая, меньшая, часть волокон оканчивается на своей же стороне. Аксоны ядер верхней оливы и трапециевидного тела (III нейрон) участвуют в образовании латеральной петли, в которой имеются волокна II и III нейронов. Часть волокон II нейрона прерывается в ядре латеральной петли (nucl. lemnisci proprius lateralis). Волокна II нейрона латеральной петли переключаются на III нейрон в медиальном коленчатом теле (corpus geniculatum mediale). Волокна III нейрона латеральной петли, пройдя мимо медиального коленчатого тела, заканчиваются в нижнем двухолмии, где формируется tr. tectospinalis. Те волокна латеральной петли, которые относятся к нейронам верхней оливы, из моста проникают в верхние ножки мозжечка и затем достигают его ядер, а другая часть аксонов верхней оливы направляется к мотонейронам спинного мозга и далее к поперечнополосатым мышцам.
Аксоны III нейрона, расположенные в медиальном коленчатом теле, пройдя через заднюю часть задней ножки внутренней капсулы, формируют глуховое сияние, которое заканчивается в поперечной извилине Гешля височной доли (поля 41, 42, 20, 21, 22). Низкие звуки воспринимаются клетками передних отделов верхней височной извилины, а высокие звуки - в ее задних отделах. Нижнее двухолмие является рефлекторным двигательным центром, через который подключается tr. tectospinalis. Благодаря этому при раздражении слухового анализатора рефлекторно подключается спинной мозг для выполнения автоматических движений, чему способствует и подключение верхней оливы с моз-жечком; подключается также медиальный продольный пучок (fasc. longitudinalis medialis), объединяющий функции двигательных ядер черепных нервов.
6. Проводящий путь статокинетического аппарата (tr. vestibularis) (рис. 501). Осуществляет передачу импульсов при изменении положения головы и тела, участвуя совместно с Другими анализаторами в ориентировочных реакциях организма относительно окружающего пространства.
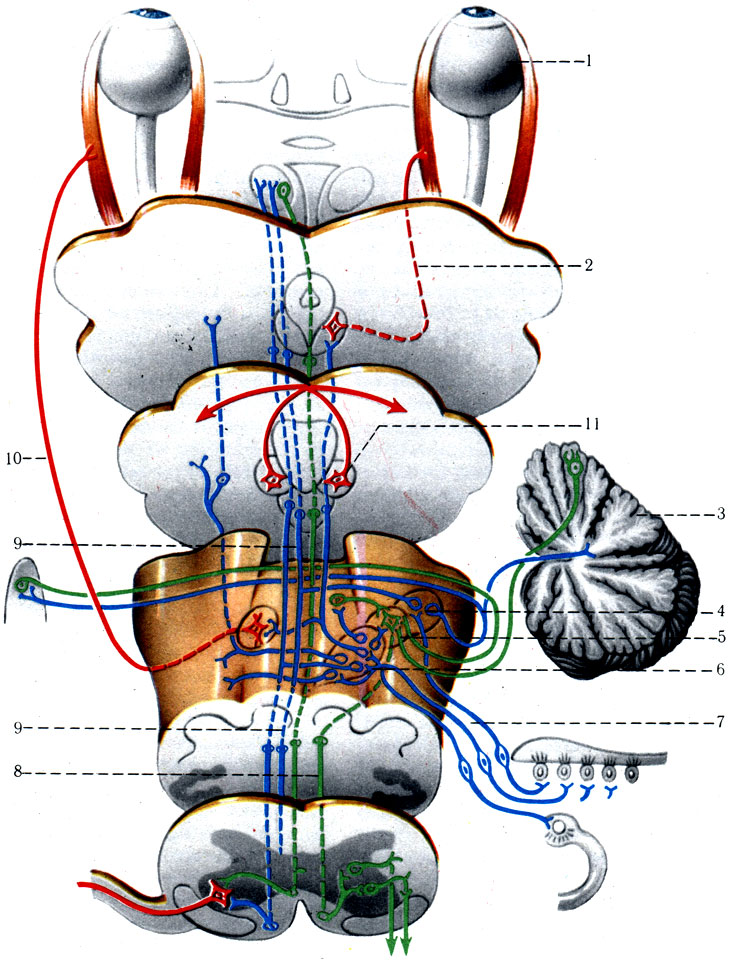
501. Схема проводящих путей статокинетического анализатора. 1 - глаз; 2 - III пара нервов; 3 - мозжечок; 4 - дорсальное преддверное ядро; 5 - латеральное преддверное ядро; 6 - нижнее и медиальное ядра; 7 - преддверный нерв; 8 - преддверно-спинномозговой путь; 9 - медиальный пучок (продольный); 10 - отводящий нерв
Первый нейрон статокинетическо-го аппарата находится в gangl. vestibulare, залегающем на дне внутреннего слухового прохода. Дендриты биполярных клеток преддверного узла формируют преддверный нерв, образованный 6 ветвями: rr. ampullaris superior, ampullaris lateralis, ampullaris inferior, ampullaris posterior, utricularis, saccularis; они контактируют с чувствительными клетками слуховых пятен и гребешков, расположенных в ампулах полукружных каналов, в мешочке и маточке преддверия перепончатого лабиринта.
Чувствительные клетки слуховых пятен и гребешков воспринимают смещение эндолимфы полукружных каналов и преддверия перепончатого лабиринта при малейшем изменении головы, при прямолинейном ускорении и вращении в трех плоскостях. Аксоны, т. е. центральные отростки биполярных клеток преддверного узла, образуют вестибулярную часть VIII пары черепных нервов совместно с n. cochlearis, которая покидает пирамиду височной кости через рorus acusticus internus, в мостомозжечковом углу проникает в вещество дорсальной части моста и продолговатого мозга, достигая верхнего, латерального, медиального и спинального ядер. Незначительное число волокон нейрона, минуя ядра, направляется непосредственно в мозжечок к nodulus, flocculus, uvula, culmen, lingula и nucl. fastigii. Между четырьмя вестибулярными ядрами существуют связи, а также двусторонние связи с ядрами ретикулярной формации.
От преддверного латерального ядра начинается преддверно-спинномозговой путь (tr. vestibulospinalis), который проходит в передней части бокового канатика спинного мозга и заканчивается на мотонейронах передних столбов. Часть аксонов нейронов латерального ядра направляется в медиальный продольный пучок одноименной и противоположной сторон, который объединяет в одно целое функцию III, IV, V, VI пар черепных нервов. В свою очередь от медиального и спинального вестибулярных ядер аксоны направляются к ядрам глазодвигательного нерва противоположной стороны, а от верхнего ядра - к глазодвигательному ядру той же стороны. От медиального ядра аксоны идут к ядру отводящего нерва. Таким образом, аксоны II нейрона четырех вестибулярных ядер образуют связи с мозжечком через tr. vestibulocerebellaris, со спинным мозгом (передние столбы) через tr. vestibulospinalis, с ретикулярной формацией (среднего, заднего и продолговатого мозга) через tr. vestibuloreticularis, с ядрами покрышки среднего мозга через tr. vestibulotectalis, с медиальным продольным пучком через fasc. longitudinalis medialis, непосредственно с ядрами III, IV, VI пар черепных нервов и таламуса.
Аксоны верхнего, латерального, медиального и спинального ядер преддверного нерва, помимо всех описанных нервных связей, образуют внутренние дуговые волокна в продолговатом мозге (fibrae arcuatae internae) и, подсоединившись к пучку медиальной петли, достигают латерального ядра таламуса, где и образуют синаптические контакты с III нейроном. От таламуса волокна направляются в корковые центры равновесия, находящиеся в средней височной извилине, лобной и теменной долях. Вероятнее всего, эти клетки рассеяны по всей коре головного мозга.
7. Проводящий путь вкусового анализатора (tr. gustus). Вкусовой анализатор информирует о химическом составе пищи, употребляемой человеком. Рецепторы вкусового анализатора находятся в физиологической связи с эпителиальными клетками вкусовых луковиц, расположенных в слизистой оболочке языка, губ, мягкого неба и надгортанника. Специализированные рецепторы восприятия соленого расположены преимущественно на кончике языка, кислого - на его краях, сладкого и горького - в основании языка и в мягком небе.
Нервные окончания chorda tympani начинаются от вкусовых луковиц слизистой оболочки ротовой полости, расположенных преимущественно в передних 2/3 языка. Волокна идут в составе n. lingualis, затем покидают его и образуют chorda tympani, которая достигает нейронов, расположенных в узле коленца (gangl. geniculi), находящемся в коленце канала лицевого нерва. Аксоны клеток узла коленца переключаются в ядрах моста - nucl. tr. solitarii VII пары черепных нервов (рис. 529).
Во вкусовых луковицах задней 1/3 языка располагаются вкусовые рецепторы языкоглоточного нерва. Дендриты достигают клеток, лежащих в его нижнем узле. Аксоны переключаются в ядрах продолговатого мозга - nucl. tr. solitarii IX пары черепных нервов.
В слизистой оболочке щек, губ и языка имеются также редко расположенные вкусовые луковицы волокон тройничного нерва. Волокна от рецепторов языка вступают в язычный нерв, от щек и губ - в щечный нерв и достигают клеток тройничного нерва. Аксоны узла переключаются в nucl. tr. solitarii VII пары черепных нервов. Волокна II нейрона от nucl. tr. solitarii VII и IX нервов переходят на противоположную сторону, подсоединяются к волокнам медиальной петли и вместе с ней направляются к вентральному и медиальному ядрам таламуса. Имеются и неперекрещенные волокна II нейрона, которые следуют к таламусу своей же стороны, оканчиваясь в вентральном и медиальном ядрах.
В таламусе располагаются III нейроны, аксоны которых проходят через заднюю ножку внутренней капсулы и оканчиваются в коре поля 43, в гиппокампе и крючке.
II. Проприоцептивные пути
1. Проводящие пути проприоцеп-тивной (глубокой) чувствительности. Состоят из пучков Голля и Бурдаха (рис. 502). С помощью этих путей совершаются движения, которые оцениваются сознанием. Управляемость движений осуществляется за счет афферентных импульсов из мышц и суставов движущихся частей тела. Импульсы достигают постцентральной извилины коры теменной доли. Эта обратная связь обеспечивает постепенность и координацию движений. При повреждении путей проприоцептивной чувствительности больной не может выполнять точные, соразмерные, ловкие движения.
Первые униполярные чувствительные нейроны путей Голля и Бурдаха располагаются в спинномозговых узлах (рис. 502). Их рецепторы - веретенообразные тельца Кюне - начинаются в мышцах, формируя затем периферический нерв. Аксоны образуют задний корешок, который вступает посегментно в белое вещество заднего канатика, объединяясь в тонкий (Голля) и клиновидный (Бурдаха) пучки. Тонкий пучок находится ближе к медиальной борозде и слагается из аксонов копчиковых, крестцовых, поясничных, XII-VII грудных сегментов. Клиновидный пучок располагается латеральнее тонкого пучка и объеди-няет аксоны от VIII-I грудных и VIII-I шейных сегментов.
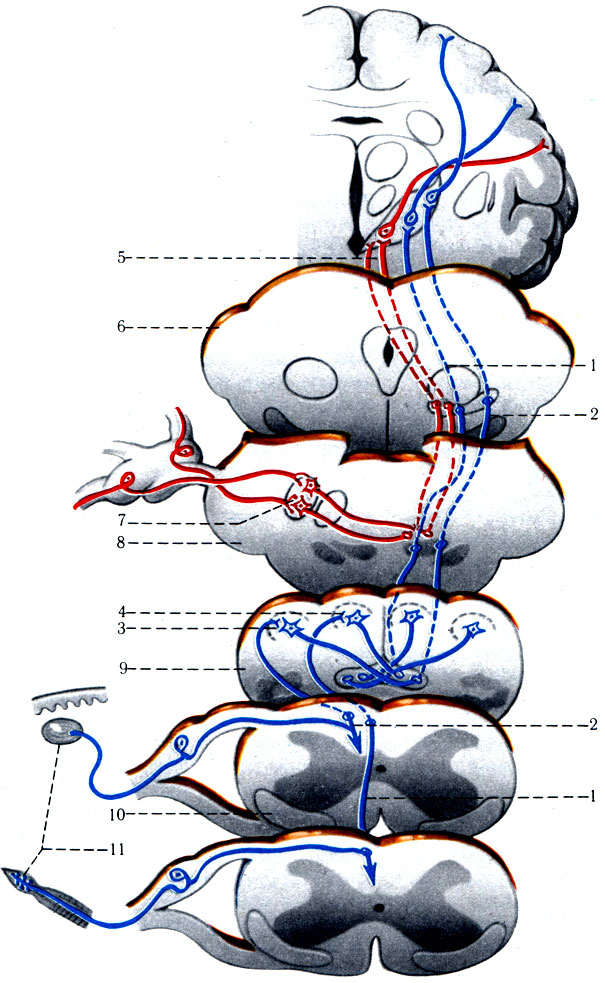
502. Схема проприоцептивных проводящих путей тройничного нерва Голля и Бурдаха (по Сен-таготаи). 1 - путь Голля; 2 - путь Бурдаха; 3 - nucl. cuneatus; 4 - nucl. gracilis; 5 - чувствительный путь тройничного нерва; 6 - средний мозг; 7 - чувствительное ядро V пары; 8 - мост; 9 - продолговатый мозг; 10 - спинной мозг; 11 - проприорецепторы путей Голля и Бурдаха
Тонкий и клиновидный пучки оканчиваются не на ядрах спинного мозга, а в тонком и клиновидных ядрах продолговатого мозга. Аксоны клеток тонкого и клиновидного ядер (II нейрон) на границе с мостом образуют медиальную петлю, которая контактирует с клетками вентролатерального ядра таламуса. С латеральной стороны к медиальной петле присоединяются волокна спиноталамического пути. Аксоны из ядер таламуса (III нейрон), пройдя через заднюю часть внутренней капсулы, оканчиваются в коре верхней теменной дольки (поля 5 и 7) и в передней центральной извилине (поля 4-6).
Часть волокон II нейронов проприоцептивных чувствительных путей направляется в мозжечок через его нижние ножки, участвуя в механизме координации движений.
Существуют проприоцептивные чувствительные пути, которые связывают ядра спинного мозга, продолговатого мозга, моста, подкорковые образования, экстрапирамидную подсистему с мозжечком, участвующие в механизмах автоматической координации движений и тонусе мышц, помимо путей, замыкающихся в коре головного мозга. Эти механизмы, как правило, проявляются при внезапных нарушениях равновесия или выполнении автоматических движений (ходьба, танцы, письмо и др.), вырабатывающихся в процессе упражнений и под влиянием социальных моментов. Безусловнорефлекторные импульсы от всех перечисленных выше образований интегрируются в мозжечке, который координирует и определяет различные по точности движения. Импульсы из мозжечка оказывают регулирующее тормозящее влияние на ядра вестибулярного анализатора и ретикулярной формации. Так как от вестибулярных ядер возникает преддверно-спинальный путь, то по нему и ретикуло-спинальному пути наступает угнетение или облегчение функции альфа- и гамма-мотонейронов передних столбов спинного мозга и мышечных веретен двигательных периферических нервов. Таким образом, благодаря механизмам обратной связи через вестибуло-спинальный и ретикулоспинальный пути мозжечок согласовывает быстрые и медленные сокращения всех мышц. Мозжечок напоминает блок регуляции, основанный на принципе обратной связи. Червь мозжечка координирует движения при ходьбе и стоянии. В полушарии мозжечка расположены механизмы очень точной координации движений, преимущественно для выполнения движений верхней конечностью. Червь подчинен коре мозжечка, а она функционирует под влиянием коры головного мозга.
2. Проводящие пути от спинного мозга к мозжечку. Задний спинно-мозжечковый путь, или путь Флексига (tr. spinocerebellaris dorsalis) (рис. 503). Имеет веретеновидные рецепторы в скелетных мышцах. Его первый специализированный чувствительный нейрон располагается в спинномозговом узле. Аксоны в составе заднего корешка достигают клеток Кларка основания заднего рога, расположенных на уровне от VII шейного до II поясничного сегмента спинного мозга, где вступают в синаптическую связь со II нейронами. Аксоны II нейронов образуют на своей стороне пучок волокон, расположенный в задней части бокового канатика (латеральнее пирамидного пути), через нижние ножки мозжечка вступают в синаптическую связь с клетками коры червя мозжечка. Из коры червя начинается III нейрон, оканчивающийся у клеток зубчатого ядра мозжечка.
3. Передний спинно-мозжечко-вый путь, или путь Говерса (tr. spinocerebellaris ventralis) (рис. 503). Его рецепторы также начинаются в мышцах. Первый специализирован-ный чувствительный нейрон располагается в спинномозговых узлах. После переключения на II нейроны в промежуточно-медиальном ядре спинного мозга их нервные волокна через переднюю серую спайку спинного мозга переходят на противоположную сторону и располагаются в передней части бокового канатика. В мосту они находятся в его дорсальном отделе, медиальнее лицевого нерва. В среднем мозге спинно-мозжечковый вентральный путь достигает нижнего двухолмия и направляется через верхние ножки мозжечка своей же стороны к коре червя. Из червя начинается III нейрон, заканчивающийся в зубчатом ядре мозжечка. Таким образом, при одностороннем включении заднего и переднего спинно-мозжечковых путей расстройства координации возникают на обеих сторонах тела.
503. Схема спинно-мозжечкового переднего и заднего, оливоспинно-мозгового, преддверно-мозжечкового, височно-мостового, мостомозжечкового, ретикуло-мозжечкового, мозжечково-красноядерного, красноядерно-спинномозгового, таламокоркового проводящих путей (по Сентаготаи). 1 - gangl. spinale; 2 - tr. spinocerebellaris anterior; 3 - tr. spinocerebellaris posterior; 4 - tr. frontotemporopontocerebellaris; 5 - tr. vestibulocerebellaris; 6 - tr. olivocere-bellaris; 7 - tr. reticulocerebellaris; 8 - nucl ruber; 9 - tr. pyramidalis; 10 - oliva; 11 - n. dorsalis; 12 - gyrus precentralis; 13 - cerebellum; 14 - tr. rubrospinal; 15 - средний мозг; 16 - мост; 17 - продолговатый мозг; 18 - спинной мозг
4. Проводящие пути от мозгового ствола к мозжечку. Преддверно-мозжечковый путь (tr. vestibulocerebellaris). Начинается преимущественно от ядра Дейтерса (nucl. lateralis) и в составе нижних ножек мозжечка достигает шатрового ядра (n. fastigii) червя мозжечка.
5. Ретикуломозжечковый путь (tr. reticulocerebellaris). Аксоны идут из ядер ретикулярной формации продолговатого мозга через нижние ножки мозжечка к его ядрам, а от ретикулярной формации моста проходят через средние ножки мозжечка к его ядрам.
6. Оливомозжечковый путь (tr. olivocerebellaris). Начинается от оливы продолговатого мозга, после частичного перекреста направляется через нижние ножки мозжечка и оканчивается в его коре.
7. Путь от ядер задних столбов продолговатого мозга к мозжечку. От нежного и клиновидного ядер (nucll. gracilis et cuneatus) продолговатого мозга начинаются волокна, которые формируют наружные дуговые волокна (librae arcuatae externае), неперекрещенные, достигающие через нижние ножки мозжечка коры червя.
Проводящие пути от коры головного мозга к мозжечку.
8. Лобно-мостовой путь (tr. frontopontinus). Начинается в лобной доле. Его волокна погружаются в белое вещество полушарий, подключаясь к системе волокон corona radiata, затем располагаются в передней ножке внутренней капсулы, внутреннего отдела основания ножки мозга, заканчиваясь в ядрах моста своей же стороны.
а) Затылочно-височно-мостовые пути (trr. occipitotemporopontini). Формируются в коре одноименных долей, направляясь в глубину полушарий, где проходят через задние ножки внутренних капсул. В среднем мозге располагаются в латеральном отделе ножек мозга, затем переключаются в ядрах моста своей же стороны.
б) Мостомозжечковые пути (tr. pontinocerebellaris). Начинаются от ядер моста, аксоны которых перекрещиваются и, соединившись в средних ножках мозжечка, достигают коры его полушарий. Поэтому расстройства координации, возникающие при одностороннем поражении лобных и височно-затылочных долей коры головного мозга, выявляются на противоположной стороне очага поражения.
9. Проводящие пути от мозжечка. Мозжечково-красноядерный путь (tr. cerebellorubralis). Аксоны клеток коры мозжечка переключаются в зубчатом ядре. Одна часть волокон через верхние ножки мозжечка после перекреста в среднем мозге достигает красных ядер. Другая часть волокон направляется в таламус. Из таламуса импульсы достигают задней центральной извилины коры. За счет этих волокон осуществляется информация о выполненной жорректировке автоматических импульсов от красного ядра, достигающих спинного мозга по красноядерно-спинномозговому пути.
10. Нисходящие (проекционные) двигательные проводящие пути, идущие от коры головного мозга к спинному и двигательным ядрам черепных нервов, разделяются на пирамидные, экстрапирамидные и волокна, проводящие торможение.
а) Пирамидный путь (tr. pyramidalis) (рис. 504). Хорошо развит у человека, так как по нему передаются импульсы поперечнополосатым мышцам при выполнении целенаправленных, тонко координированных сознательных движений. Пирамидные пути существуют и у многих животных, но функционируют без сознательной корректировки. Двигательные клетки коры не иннервируют отдельно ту или другую мышцу, а осуществляют заданную программу движений для отдельных групп мышц. Пирамидный путь получил свое название от двух клиновидной формы выпуклостей, лежащих на вентральной поверхности продолговатого мозга. Многие годы считалось, что все волокна пирамидного пути начинаются от клеток коры передней центральной извилины. В настоящее время установлено, что только около 40% аксонов, проходящих через пирамиды, начинается от клеток двигательной зоны коры, а 20% аксонов пирамидного пути начинается от клеток задней центральной извилины (соматосенсорная область). Остальные 40% волокон присоединяются к пирамидному пути от клеток различных областей коры полушарий большого мозга.
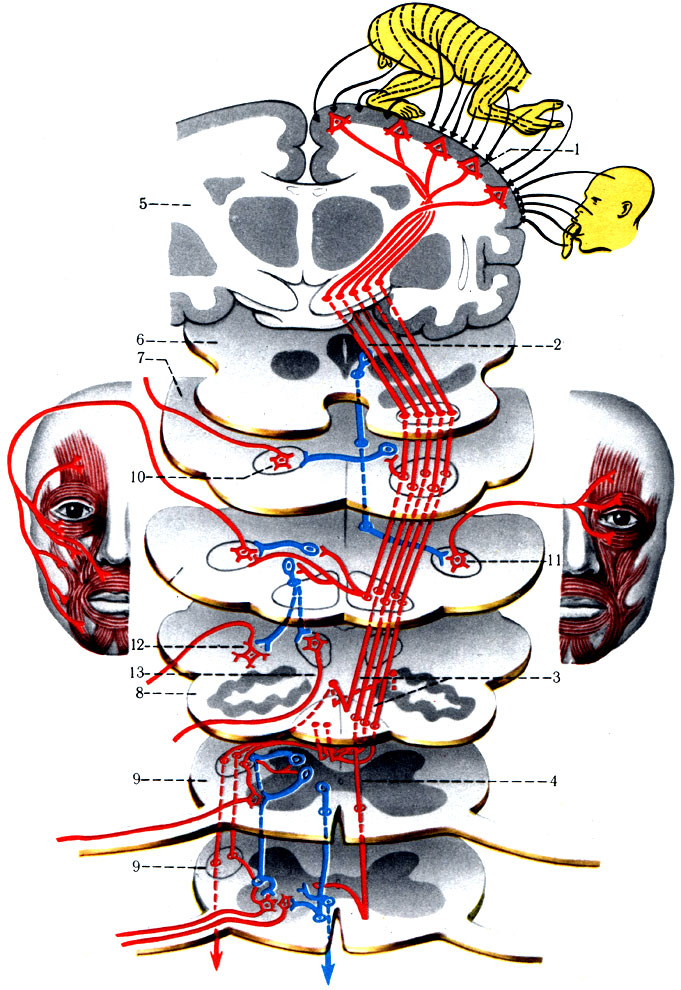
504. Схема пирамидного пути (по Сентаготаи). 1 - gyrus precentralis; 2 - tr. corticonuclearis; 3 - tr. corticospinalis lateralis; 4 - tr. corticospinalis anterior; 5 - полушарие головного мозга; 6 - средний мозг; 7 - мост; 8 - продолговатый мозг; 9 - спинной мозг; 10 - двигательное ядро V пары; 11 - двигательное ядро VII пары; 12 - двигательные ядра IX, X, XI пар; 13 - ядро XII пары
Первые нейроны располагаются в передней центральной извилине, предцентральной и парацентральной дольках (поля 4-6), часть нейронов разбросана в других корковых полях (7-8-9-22-24 и др.). Существенным моментом является то, что все корковые поля пирамидного пути связаны с нейронами, которые своей деятельностью подавляют моторную активность двигательной зоны и находятся в полях 2-4-8-19. Подобная тормозящая система отсутствует у других проводящих путей. Кроме того, в поле 4 есть участок 4S, откуда специальные аксоны достигают ядер ретикулярной формации, которая оказывает тормозящее или возбуждающее влияние на произвольные рефлексы. Дендриты пирамидных клеток имеют связь со вставочными нейронами, подключающими чувствительные клетки всех анализаторов. Эти вставочные нейроны формируют короткие и длинные ассоциативные пути белого вещества.
В передней центральной извилине и парацентральной дольке имеются специализированные участки коры, которые осуществляют программу, заданную определенным группам мышц: мышцы нижних конечностей находятся под контролем клеток верхних отделов (ближе к сагиттальной борозде большого мозга) передней центральной извилины и пара-центральной дольки, мышцы верхних конечностей - клеток среднего отдела центральной извилины, мышцы лица и органов головы - клеток нижнего отдела.
Пирамидный путь включает три пучка: а) корково-ядерный путь (tr. corticonuclearis), осуществляющий центральное кодирование программы движений в двигательных ядрах черепных нервов (III, IV, V, VI, VII, IX, X, XI, XII пары); б) передний корково-спинальный путь (tr. corticospinalis anterior); в) боковой корково-спинальный путь (tr. corticospinalis lateralis). Оба последних пучка проводят импульсы программы движений к мотонейронам спинного мозга.
Первые нейроны пирамидного пути находятся в различных областях коры полушарий головного мозга. В V слое коры головного мозга имеются пирамидные клетки Беца, аксоны которых принимают участие в образовании лучистого венца белого вещества полушарий мозга. Эти волок-на конвергируют вниз, проходя в колене и в 2/3 задней ножки внутренней капсулы. Пирамидные клетки имеют длинные аксоны и большое число коллатералей, которые подключают несколько двигательных клеток II нейронов.
Волокна пирамидного пути, пройдя внутреннюю капсулу, располагаются в основании ножки мозга, где от них отделяются перекрещенные волокна к ядрам глазодвигательного нерва (иннервирующего, верхнюю, нижнюю, медиальную прямые, нижнюю косую мышцы глазного яблока и мышцу, поднимающую верхнее веко), к ядру блокового нерва (иннервирующего верхнюю косую мышцу глазного яблока) и к ядру отводящего нерва (иннервирующего латеральную прямую мышцу глазного яблока).
Из основания ножки мозга пирамидный путь спускается в вентральную часть моста, на уровне которого отделяются перекрещенные волокна для контакта с двигательным ядром тройничного нерва (иннервирующего жевательные мышцы), с двигательным ядром лицевого нерва (иннервирующего мимическую мускулатуру); некоторые волокна отдают коллатерали в ретикулярную формацию. Пучок пирамидного пути расположен в мосту некомпактно, через него поперечно проходят волокна корково-мостомозжечкового пути (описан в разделе "Проприоцептивные пути"). В продолговатом мозге волокна пирамидного пути объединяются в компактный пучок и на вентральной поверхности продолговатого мозга образуют пирамиды. В каждом из двух трактов пирамидных путей содержится около 1 млн. волокон, главным образом тонких и скудно миелинизированных; около 3% волокон имеет большой диаметр и покрыто толстой миелиновой оболочкой; они являются аксонами клеток Беца. В продолговатом мозге двигательные ядра языкоглоточного (IX пара), блуждающего (X пара), добавочного (XI пара), подъязычного (XII пара) нервов также контактируют с волокнами пирамидного пути. Волокна пирамидного пути, направляющиеся к ядрам двигательных черепных нервов, перекрещиваются. Эти ядра получают иннервацию от волокон своей и противоположной сторон. Поэтому при центральном одностороннем поражении коры полушарий мозга или проводящих путей не бывает полного паралича мышц, иннервируемых III, IV, V, VI, VII, IХ, X, XI парами черепных нервов. В области пирамид продолговатого мозга небольшая часть волокон пирамидного пути, огибая нижнюю оливу через нижнюю или среднюю ножку мозжечка, входит в него.
В нижней части продолговатого мозга пирамидный путь разделяется на два пучка. Один большой пучок (около 80% волокон) перекрещивается (decussatio pyramidum) и переходит в латеральный канатик спинного мозга, образуя боковой корково-спинномозговой путь (tr. corticos-pinalis lateralis). Волокна этого пути оканчиваются возле дендритов вставочных клеток (II нейрон), расположенных в задних столбах спинного мозга. Аксоны этих клеток передают импульсы вставочным клеткам (III нейрон) переднего столба, а последние - большим альфа-нейронам (IV нейрон) переднего столба, от которых импульсы направляются к малым альфа-нейронам (V нейрон), а также к мышцам конечностей и туловища.
Меньшая часть пирамидного пути в продолговатом мозге не перекрещивается и спускается в переднем канатике под названием переднего корково-спинномозгового пути (tr. corticospinalis anterior). В каждом сегменте спинного мозга его аксоны переходят на противоположную сторону, переключаясь в передних столбах одной частью на вставочные нейроны (II нейрон) и другой - на мотонейроны (II нейрон). Аксоны вставочных нейронов подключаются к малым альфа-нейронам (III нейрон), аксоны которых достигают мышц туловища и конечностей (рис. 505). Волокна вставочных нейронов прослеживаются в шейных и верхних грудных сегментах спинного мозга. Часть волокон переднего корково-спинномозгового пути переключается в мотонейронных пулах своей стороны.
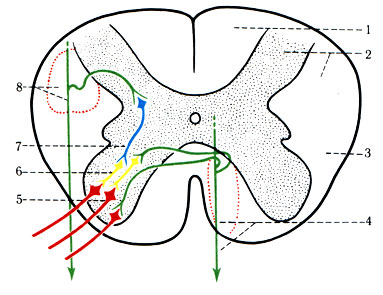
505. Схема переключения кортико-спинального пути (пирамидного) в спинном мозге. 1 - задний канатик; 2 - задний столб; 3 - боковой канатик; 4 - передний кортико-спинальный путь; 5 - крупные мотонейроны переднего столба; 6 - вставочные нейроны переднего столба; 7 - вставочные нейроны заднего столба; 8 - латеральный корково-спинальный путь
Аксоны периферического спинномозгового нерва, являющиеся отростками крупных мотонейронов передних столбов серого вещества спинного мозга, иннервируют экстрафузальные мышечные волокна поперечнополосатых мышц. Каждое волокно имеет химически чувствительную область - концевую пластинку, где оканчивается двигательный аксон; она эквивалентна постсинаптической мембране нейрона. При возбуждении аксон мотонейрона выделяет ацетилхолин, воздействующий на концевую пластинку при этом наблюдается деполяризация мышечного волокна и генерация электрического импульса, который распространяется в обе стороны к концам мышечного волокна, вызывая кратковременное его сокращение.
Следовательно, пирамидный путь осуществляет главным образом перекрестную иннервацию. Поражение бокового корково-спинномозгового пути вызывает расстройство движений конечностей на противоположной стороне и почти не нарушает функции мышц туловища вследствие сохранения иннервации за счет переднего кортико-спинального пучка. Такой односторонней иннервацией обладают не все мышечные группы. Большая часть мышц, а именно мышцы глазного яблока, жевательные, мимические мышцы верхней части лица, глотки, гортани, шеи, туловища и промежности, имеет двустороннюю иннервацию за счет волокон перекрестных и своей стороны. Односторонне иннервируются мышцы конечностей, языка, мимические мышцы ниже ротовой щели. Поражение соответствующих клеток коры вызывает полный паралич.
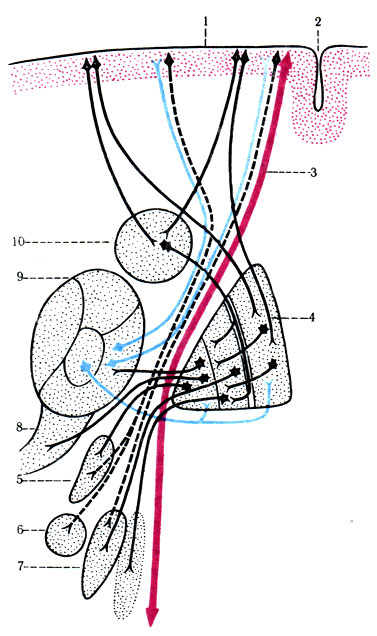
506. Связь коры головного мозга с базальными ядрами, таламусами, ретикулярной формацией и ядрами подталамической области. 1 - корковые поля; 2 - центральная борозда; 3 - волокна пирамидного пути; 4 - чечевицеобразное тело; 5 - луизово тело; 6 - черная субстанция; 7 - рекикулярная формация; 8 - подталамическое ядро; 9 - зрительный бугор; 10 - хвостатое тело
б) Экстрапирамидные пути. Подразделение двигательной системы на пирамидную и экстрапирамидную является источником путаницы и ошибок, так как вначале пирамидную систему представляли единственной двигательной системой. Вероятно, поэтому все двигательные пути, описанные позднее, были объединены в экстрапирамидную систему. Провести четкую грань в анатомическом строении и функциональном значении между пирамидной и экстрапирамидной системами практически невозможно, так как они функционируют в полном взаимодействии. С помощью экстрапирамидной системы тело поддерживается в нужном положении и выполняются содружественные движения с регуляцией мышечного тонуса при выполнении безусловнорефлекторных двигательных актов.
К экстрапирамидной системе относятся многие рассеянные двигательные клетки среди чувствительной зоны, базальные ядра (бледный шар, скорлупа, хвостатое ядро, ограда), передненижнее ядро таламуса, субталамические ядра, мозжечок, красное ядро, черная субстанция и ядра ретикулярной формации.
Экстрапирамидная проводящая система начинается от разбросанных по всей коре головного мозга двигательных клеток, аксоны которых поступают через заднюю ножку внутренней капсулы к хвостатому ядру и скорлупе, а затем после переключения идут к бледному шару. От наружного членика бледного шара аксоны направляются в черное вещество, красное ядро, медиальное вестибулярное ядро и ретикулярную формацию.
От ядер ретикулярной формации берет начало ретикулярно-спинно-мозговой путь (tr. reticulospinalis), вступающий в контакт со вставочными нейронами и малыми альфа-нейронами передних столбов спинного мозга. От красных ядер формируется tr. rubrospinalis, входящий в боковой канатик спинного мозга и контактирующий со вставочными нейронами заднего и переднего столбов спинного мозга, которые переключаются на малые альфа- и гамма-нейроны.
В экстрапирамидной системе имеется замкнутая петля проводящих путей; она начинается в бледном шаре и проходит к таламусу, где есть центр "активации", затем к скорлупе и вновь возвращается к бледному шару. Бледный шар связан с моторными клетками коры, которые и активируются под действием возвратных импульсов, посылая прямые сигналы к спинному мозгу через пирамидные пути. Допускается, что полосатые тела (хвостатое ядро, скорлупа, бледный шар) являются первичными инициаторами двигательной активности после получения сигналов из сенсорных областей. Кора не доминирует над полосатым телом, но в процессе формирования движений повышает способность экстрапирамидной системы к включению в рефлекторный процесс и увеличивает ее гибкость в процессе перестройки.
в) Волокна, проводящие торможение разделяются на центральные и периферические.
Центральное торможение формируется специализированными нейронами ядер стволовой части мозга и коры полушарий. При централь-ном торможении импульсы идут от высших центров по эфферентным нервным волокнам к сенсорным органам. Среди избытка информации тормозные импульсы играют большую роль в отборе воспринимаемых импульсов головным мозгом. В этом случае при переключении внимания на какой-нибудь стимул импульсы других сенсорных систем угнетаются импульсами торможения, направляя необходимые стимулы в определенный "канал". За счет торможения производится фильтрация импульсов и их суммация, что обеспечивает локальность возбуждения и его целенаправленность. Импульсы торможения создают гиперполяризацию рецепторной мембраны, тем самым инактивируют рецептор, что нарушает восприятие раздражений.
Периферическое торможение возникает при взаимодействии сенсорных клеток, действуя на сами сенсорные клетки или синапсы. Периферическое торможение осуществляется за счет латеральных нервных волокон, соединяющих соседние рецепторы и клетки. Например, при нагрузке на мышцу сокращение развивается равномерно во всех экстрафузальных мышечных волокнах. Это распределение осуществляется за счет латеральных нервных волокон, при помощи которых возбужденный мотонейрон тормозит себя и другие нейроны соответствующего двигательного пула, возбуждая тормозные клетки Реншоу. Аксоны клеток Реншоу в свою очередь также заканчиваются на мотонейронах. Петля обратной связи тормозит повторное быстрое возбуждение мотонейронов передних столбов спинного мозга.
В действительности иннервация поперечнополосатых мышц значительно сложнее, чем это можно изложить при описании проводящих путей.
Каждая мышца человека одинаково хорошо снабжена чувствительными и двигательными нервами. Один мотонейрон спинного мозга иннервирует одновременно большое число мышечных волокон, объединяя их в двигательную единицу. Естественно, что в иннервации мышц принимают участие несколько двигательных единиц. Мотонейроны, обеспечивающие иннервацией одну мышцу, не представлены единым компактным ядром, а рассеяны по нескольким сегментам спинного мозга и функционально объединяются в двигательный пул. В пределах каждого двигательного пула имеется сложная система вставочных нейронов, объединяющая в одно целое альфа-и гамма-мотонейроны спинного мозга. Этот комплекс вставочных нейронов частично описан при изложении бокового кортико-спинального пути и рефлекторной дуги.
Чтобы понять принцип иннервации мышц, необходимо кратко познакомиться со строением мышечной ткани. Каждая поперечнополосатая мышца имеет неоднородные экстрафузальные мышечные волокна, иннервируемые концевыми пластинками двигательных нервов. Кроме того, в меньшем числе встречаются тонкие интрафузальные мышечные волокна, образующие мышечные веретена. В каждом мышечном веретене может быть по 2-10 интрафузальных мышечных волокон, лежащих в концевых частях веретена. Задачей интрафузальных мышечных волокон является натяжение мышечной трубки, где находится первичный чувствительный рецептор. Эта трубка расположена между двумя сократительными концами веретена (в них находятся интрафузальные мышечные волокна) и чрезвычайно чувствительна к малейшему сокращению интрафузальных волокон или растяжению мышцы. Концы мышечных веретен прикреплены к оболочке экстрафузальных волокон, так что при малейшем изменении их длины меняется натяжение мышечных веретен и соответственно их мышечных трубок. При изотоническом сокращении мышечное волокно укорачивается и натяжение на первичном рецепторе уменьшается. Когда мышца расслабляется, ее веретена растягиваются и первичные рецепторы возбуждаются. Следовательно, первичный рецептор может растягиваться в 3 случаях: при расслаблении экстрафузальных волокон, сокращении интрафузальных волокон и растяжении мышц за счет внешней нагрузки. Аксон, идущий от первичного рецептора мышечного веретена, возвращается в соответствующий двигательный пул, из которого выходит нерв к экстрафузальным мышечным волокнам. При растяжении мышечной трубки в ней генерируются импульсы, которые поступают в задние столбы спинного мозга.
В чем заключается действие этой обратной связи? При выполнении движения значение ее очень велико, так как только с помощью обратной связи возможна четкая последовательность сокращения и расслабления сгибателей и разгибателей, приводящих и отводящих мышц, супинаторов и являющихся антагонистами пронаторов. Пирамидная или экстрапирамидная система может генерировать сигналы или передавать их с сенсорных систем, достигающих двигательного пула спинного мозга и подключающих почти исключительно мелкие мотонейроны, которые, возбуждаясь, вызывают сокращение интрафузальных волокон мышечных веретен. Это приводит к растяжению и возбуждению первичного рецептора, находящегося на мышечной трубке мышечного веретена. Импульсы от него достигают спинного мозга и возбуждают крупные мотонейроны. Аксоны крупных мотонейронов формируют двигательный нерв, по которому передается импульс экстрафузальным мышечным волокнам, вызывающий сокращение мышцы. В процессе сокращения мышц первичные рецепторы мышечной трубки угнетаются и постепенно импульсы с мотонейронов затухают. Действие обратной связи мышечного веретена наглядно проявляется, например, при сухожильном рефлексе. Удар по сухожилию расслабленной мышцы приводит к сокращению интрафузальных волокон, которые посылают крупным мотонейронам залп импульсов.
После короткой задержки возникает внезапное сокращение четырехглавой мышцы. При длительном изотоническом сокращении мышцы крупные мотонейроны спинного мозга должны постоянно генерировать импульсы для сокращения экстрафузальных мышечных волокон. В этом случае поступающие даже слабые импульсы из моторной зоны коры головного мозга возбуждают только мелкие мотонейроны, которые вызывают сокращение интрафузальных мышечных волокон. Оно приводит к натяжению мышечных трубок и генерации импульсов для возбуждения крупных мотонейронов всего двигательного пула спинного мозга данной мышцы. Таким образом автоматически поддерживается изотоническое сокращение.
Механизм обратной связи мышечного веретена является весьма эффективным в случаях запредельных сокращений. Теоретически допустимо, что вполне возможно повреждение сухожилия и даже костей при максимальном сокращении, когда возбуждаются достаточно сильно крупные мотонейроны двигательного пула спинного мозга. Подобных запредельных сокращений в здоровом организме не происходит, так как в сухожилиях мышц имеются рецепторы Гольджи. Эти рецепторы выдают тормозные залпы, вызывающие угнетение мотонейронов спинного мозга.
При использовании материалов сайта активная ссылка обязательна:
http://anfiz.ru/ 'Анатомия и физиология человека'