Место, занимаемое человеком в системе животных. М. А. Мензбир
Человек по своей организации принадлежит к обширной группе позвоночных животных, которые составляют одно из трех подразделений (подтипов) типа хордальных. Все хордальные характеризуются присутствием у них или в течение всей жизни, или в течение ранних стадий развития особого органа, спинной струны, или хорды (chorda dorsalis), которая у огромного большинства хордальных может быть названа осевым скелетом. Кроме того, за исключением одной группы, у всех хордальных есть так называемые глоточные щели, соединяющие полость глотки с наружной средой, и нервная система с полостью внутри.
Однако эти три основные особенности хордальных выражены совершенно ясно только у позвоночных, характеристика которых является в таком виде. В основе своего строения позвоночные представляют собою двусторонне симметричных животных, т. е. таких, у которых правая половина относится к левой (или обратно), как предмет к своему изображению в зеркале. В теле всех позвоночных имеются две полости, видимые как на продольном сагиттальном, так и на поперечном разрезе. Та из них, которая лежит на дорзальной стороне, содержит центральную нервную систему и называется спинным каналом; та, которая лежит на вентральной стороне, содержит разные внутренние органы и называется общей полостью тела (coelom) (рис. 1). Между этими двумя полостями в разделяющей их стенке лежит спинная струна, представляющая собою стержень или шнур из вакуолизированных клеток, одетый особыми оболочками. Самая наружная из этих оболочек продолжается отростками справа и слева вверх, очерчивая ими спинной канал, где помещается центральная нервная система, и соответствующими отростками вниз, в стенки общей полости тела. Центральная нервная система сохраняется всю жизнь в виде лежащей в спинном канале нервной трубки с продольным каналом внутри. У большинства позвоночных передний конец спинного канала расширяется в черепную полость, соответственно чему центральная нервная система делится на шнуровидный спинной мозг, суживающийся кзади, и расширенный передний конец - головной мозг. Та часть спинного канала, в которой лежит спинной мозг, называется спинномозговым каналом; головной мозг находится в полости черепа. У огромного большинства позвоночных во взрослом состоянии спинная струна заменяется так называемым позвоночным столбом, расчлененным на сегменты, называемые позвонками. В черепе позвонков нет. Как дополнительная часть черепа у всех позвоночных существует так называемый висцеральный скелет, состоящий из парных висцеральных дуг, из которых первая служит в качестве челюстной, вторая - подъязычной, а остальные, в числе от семи до трех в разных группах, - в качестве жаберных. Эти дуги залегают в боковых стенках глотки, разделяя жаберные щели, которые у позвоночных, живущих в воде, служат для соединения глотки с наружной средой и при помощи сидящих на их краях жаберных лепестков играют роль органов водного дыхания. У рептилий, птиц и млекопитающих эти щели существуют лишь в виде углублений на боковых стенках глотки и не прорезываются.
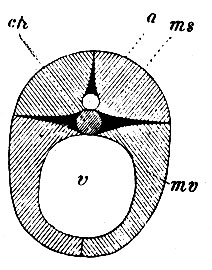
Рис. 1. Схематичный поперечный разрез тела позвоночного. А - спинномозговой канал; ch - хорда; ms - спинные мышцы; mv - брюшные мышцы; v - общая полость тела (coelom)
Кроме того, у всех позвоночных существует очень важный отдел кровеносной системы, называемый воротной системой печени, т. е. кровь, идущая от стенок кишечного канала к сердцу, не прямо попадает в сердце, а сначала проходит через особую железу - печень, которая развивается как вырост кишечного канала. Особенность воротной системы заключается в том, что она начинается и оканчивается капиллярами.
У всех позвоночных, за очень немногими исключениями, имеются парные конечности, которых никогда не бывает более двух пар. У ланцетника и миног их неизвестно ни в каком возрасте; у других позвоночных, лишенных конечностей, они отсутствуют вследствие недоразвития. У позвоночных конечности развиваются из парных почковидных выростов по бокам тела.
Отношение челюстей к конечностям составляет весьма важный отличительный признак от беспозвоночных. Тогда как у позвоночных челюсти развиваются из первой висцеральной дуги, у членистоногих они представляют собою видоизмененные конечности.
Общая полость тела (coelom) образуется у всех позвоночных, кроме ланцетника, путем расщепления мезодермы (см. ниже Главные черты развития позвоночных), следовательно, очерчивается ее наружной пластинкой, так называемой соматоплеврой, и внутренней, или спланхноплеврой. Наружная одета снаружи эктодермическим эпителием и образует стенку тела; внутренняя облекает в виде трубки железистую выстилку кишечного канала и составляет вместе с нею стенку кишечного канала. Мышцы, развивающиеся из мезодермы, сегментированы, и эта сегментация на известных стадиях развития продолжается в задний отдел головы, оканчиваясь в области глазного яблока. Сюда же заходит и передний конец хорды, оканчивающийся дойдя до области воронки и гипофиза, т. е. до третьего желудочка. Следовательно, задний отдел головы сегментирован, как и туловище, но эту сегментацию не должно переносить на череп, который, как выше сказано, ни на какой стадии развития не бывает сегментирован.
Общая полость тела выстлана собственной оболочкой, которая по своим особым свойствам называется серозной. У низших позвоночных общая полость тела является сплошной, у высших, и в том числе у человека, от нее отделяется часть, заключающая в себе легкие (плевроперитонеальная полость), и часть, заключающая в себе сердце (перикардиальная полость).
Органы выделения представлены у позвоночных выделительными канальцами, которые распадаются на три группы, называемые передними, или головными, почками, средними, или туловищными, и задними. У низших позвоночных развиваются только передние, которые у многих во взрослом состоянии заменяются средними. У высших передние имеют быстро преходящее значение и в зародышевом состоянии уже заменяются средними, которые у человека обыкновенно известны под названием вольфовых тел. Во взрослом состоянии вольфовы тела заменяются задними или окончательными почками.
У всех позвоночных, кроме ланцетника, центральный орган кровообращения составляет сильно мускулистый орган - сердце, лежащее на вентральной стороне. У ланцетника сердца нет, и его отсутствие заменяется пульсацией всех крупных сосудов.
При описании строения тела позвоночных обыкновенно употребляют следующие термины: продольная ось, идущая от переднего конца тела к заднему и лежащая горизонтально; две другие оси лежат как друг к другу, так и к продольной оси под прямым углом, одна из них идет справа налево и называется поперечной, являясь в то же время лежащей в горизонтальной плоскости, другая идет перпендикулярно к ним и называется спинно-брюшной, так как соединяет спинную (дорзальную) сторону с брюшной (вентральной). Плоскость, проходящая через продольную и спинно-брюшную ось, называется медиальной; она делит тело животного на две равные и симметричные половины. Плоскость, проходящая через поперечную и спинно-брюшную ось, называется поперечной. Различают еще фронтальную плоскость, параллельную фронтальной поверхности черепа, и сагиттальную, к ней перпендикулярную.
При помощи этих осей и плоскостей точно определяется положение органов в теле любого позвоночного.
Однако надо помнить, что у большинства позвоночных во взрослом состоянии происходит более или менее значительное нарушение двубоковой симметрии, особенно это касается непарных органов, находящихся в общей полости тела. Особенно нарушается положение кишечного канала вследствие его чрезвычайного удлинения, неодинаково развиты правое и левое легкое, у некоторых позвоночных правая и левая ночка, сердце у человека, например, значительно сдвигается влево и т. д.
Затем в приложении к человеку в частности вышеприведенная номенклатура должна быть изменена вследствие вертикального положения его тела. Так, продольная ось становится для него вертикальной, вентральная сторона - передней, дорзальная - задней, передние парные конечности - верхними, задние - нижними и т. д. Все эти изменения понятны сами собой.
Позвоночные развиваются только из яиц. Каждое яйцо представляет собой клетку, но яйца разных животных по виду и по величине разнятся между собой очень сильно. Это различие обусловливается присутствием образований, имеющих значение или при процессах дальнейшего развития (питательный желток), или для защиты яйца от внешних влияний (различные оболочки). Наиболее простое строение имеют яйца Amphioxus и высших позвоночных - млекопитающих, исключая Monotremata. Последнее объясняется тем, что яйцо всех млекопитающих, кроме Monotremata, проходит цикл своего развития в полости матки и, следовательно, не требует сложных оболочек для своей защиты. Кроме того, кровеносная система зародыша млекопитающих рано вступает в особую связь, с кровеносной системой матери, на счет которой зародыш питается, следовательно, яйцо млекопитающих не требует такого большого запаса питательного желтка и потому оно мало (0,2 мм). Яйцо млекопитающих состоит из протоплазмы, в которой заключены в небольшом количестве жироподобные тела питательного желтка (deutoplasma) и крупное ядро (vesicula germinativa) с ядрышком (macula germinativa). Оболочка зрелой яйцеклетки довольно толста и пронизана радиальными канальцами (zona pellucida seu radiata). Эта оболочка отлагается фолликулярным эпителием, на счет которого яйцеклетка питается в течение времени своего созревания. Яйца птиц наиболее крупны, так как содержат много желтка и покрыты очень сложными защитными оболочками. Зрелое яйцо птицы, готовое покинуть фолликул яичника, образует тело, которое в общежитии называется желтком. Оно представляет собой единичную клетку, сильно увеличенную содержанием питательного желтка, и имеет следующее строение. На поверхности желточной сферы находится в определенном месте беловатое чечевицеобразное тело (discus proligerus seu cicatricula); оно имеет от 3 до 4 мм в диаметре, состоит из мелкозернистой протоплазмы, содержащей ядро яйцеклетки и особый вид шариков питательного желтка, называемых тельцами "белого желтка". В cicatricula происходят начальные процессы развития. Тельца белого желтка, включенные в протоплазму cicatricula, придают ей беловатый вид. Тонкий беловатый слой, отходящий от краев cicatricula, продолжается к центру желтка, где заканчивается колбообразным вздутием (latebra). Весьма вероятно, что периферический слой белого желтка, равно как и центральное образование (latebra), содержит, кроме телец желтка, и протоплазму, распространяющуюся от cicatricula между тельцами желтка в виде сети. Так называемый желтый желток имеет слоистое строение, тельца, его слагающие, более крупны, чем тельца белого желтка, и несколько отличаются строением своего содержимого. Вероятно, пласмодическая сеть, отходящая от краев cicatricula, заходит и в промежутки между тельцами желтого желтка. Вся желточная сфера с cicatricula одета очень тонкой эластической оболочкой (membrana vitellina seu tunica adventitia), которая соответствует zona pellucida яйца млекопитающих.
Описанное строение имеет яйцо птицы, готовое начать свое путешествие по яйцеводу. В этом виде оно оплодотворяется. Во время движения по яйцеводу яйцо одевается слоем белка и системой оболочек, выделяемых железистыми аппаратами стенок яйцевода. В снесенном яйце толстый слой жидкого белка, окружающий желток (яйцеклетку), облекается двуслойной, волокнистой, но не состоящей из клеток оболочкой (membrana testae). У тупого полюса яйца, между обеими пластинками membrana testae, находится пространство, наполненное воздухом. Снаружи яйцо одето пористой известковой скорлупой (testa).
Процессы развития начинаются в яйце после соединения его с половым продуктом мужской, особи - сперматозоидом (оплодотворение). Сперматозоид позвоночных животных есть, так же как и яйцо, клетка, но приноровленная к активному передвижению. Элементы ядра (хроматин) заключены в головке сперматозоида. Большая часть протоплазмы вытянута в жгутик, служащий для движения сперматозоида и представляющий большую аналогию с ресничками клеток мерцательного эпителия. Процесс оплодотворения сводится к процессу конъюгации двух клеток, яйца и сперматозоида. Элементы ядра обеих клеток вступают в тесное соединение друг с другом и образуют одно ядро оплодотворенной яйцеклетки.
Развитие зародыша начинается процессом деления яйцеклетки на 2, 4, 8 и т. д. шаров дробления; по мере хода процесса шары дробления становятся меньше, в результате чего получается скопление мелких, так называемых эмбриональных клеток.
При процессах дробления яйца наблюдаются те же морфологические изменения ядра, что и при процессах деления клеток любой ткани, и весь процесс дробления сводится в сущности к процессу размножения клеток. При начальных стадиях дробления некоторых беспозвоночных животных, несомненно, констатировано, что элементы ядер шаров дробления образуются как из элементов ядра яйцеклетки, так и из элементов, внесенных в яйцеклетку ядром сперматозоида,- и те, и другие в одинаковом количестве. Из этого следует, что каждая клетка строящегося организма, по крайней мере в начальных стадиях развития, а вероятно, и в более поздних, содержит материальные элементы как женского, так и мужского половых продуктов. Этим замечательным фактом в значительной мере освещаются явления наследственности.
В зависимости от количества и способа распределения питательного желтка в яйцеклетке процессы дробления различных позвоночных представляют более или менее значительные различия. Если количество питательного желтка невелико и он распределяется равномерно в образующихся шарах дробления, то борозды, отделяющие шары друг от друга, распространяются сквозь всю толщу желтка. Яйца с таким полным дроблением называются голобластическими. Голобластические яйца имеют миноги, хрящевые ганоиды, амфибии и все млекопитающие, кроме Monotremata. Если яйцеклетка содержит много питательного желтка и он распределяется в образующихся шарах дробления очень неравномерно, то дробление происходит только у одного полюса яйцеклетки, называемого анимальным (животным). Противоположный полюс, вегетативный (растительный), не участвует в дроблении. Только в последующем развитии он обрастает тканями, надвигающимися из области анимального полюса. Яйца с таким неполным дроблением называются меробластическими. Меробластические яйца имеют акулы, пресмыкающиеся, птицы и самые низшие млекопитающие, Моnotremata, яйцо которых имеет поразительное сходство с яйцом пресмыкающихся и птиц. У некоторых отдельно стоящих форм наблюдаются переходные состояния между голо- и меробластическими яйцами (Lepidosteus).
Выше мы сказали, что в конечном результате процесса дробления получается скопление эмбриональных клеток. Следующий процесс развития состоит в группировке эмбриональных клеток в два слоя, которые называются первичными эмбриональными листами. Наружный слой называется эктобласт или эктодерма, внутренний - эндобласт или эндодерма. Способы образования первичных листов у различных позвоночных весьма различны, но у всех позвоночных из эктобласта развиваются эпидермис, кожные железы, центральная нервная система и важнейшие составные части органов чувств, словом, так называемые органы животной жизни. Из эндобласта развивается кишечник с его железами (печень, pancreas и т. д.), а также и легкие кроме того, самый существенный отдел кровеносной системы - эндотелий сердца. Словом, из эндобласта развиваются органы растительной жизни. Так как группы перечисленных систем органов совершенно отличны по своему физиологическому значению, то эктобласт и эндобласт называются первичными эмбриональными листами; они представляют собою первое выражение физиологической специализации в однородном скоплении эмбриональных клеток.
Между эктобластом и эндобластом появляется третий слой клеток, так называемый средний лист, или мезобласт, или мезодерма. Существенным в его образовании является следующее: во-первых, мезобласт появляется после экто- и эндобласта и, следовательно, представляет собой вторичную диференцировку зародыша. Во-вторых, он развивается как из экто-, так и из эндобласта. Участие первичных листов в образовании главной осевой части мезобласта у различных позвоночных весьма различно (первичная борозда, край бластопора, так называемые дивертикулы coelom и т. д.). В-третьих, развитие некоторых отделов мезобласта продолжается в течение начального развития долгое время, причем происхождение этих отделов нельзя приурочить к определенному первичному листу (эктобластическое происхождение так называемой мезенхимы, эндобластическая мезенхима головного отдела кишечника). На основании всего этого мезобласт называется вторичным эмбриональным листом и морфологически несравним с двумя первыми. Из мезобласта развиваются стенки общей полости тела (coelom), мускулатура, chorda (Goette), весь скелет, половые железы и важнейшие отделы органов выделения.
Главные процессы дальнейшего развития эмбриональных листов состоят в следующем. Соответственно продольной оси зародыша эктобласт утолщается. Утолщенная полоса верхнего листа называется медуллярной пластинкой и представляет зачаток всей центральной нервной системы. У всех позвоночных, кроме ланцетника, передний отдел медуллярной пластинки расширен, задний более узок. Передний отдел называемся головной пластинкой и представляет зачаток головного мозга, задний есть зачаток спинного мозга. Далее боковые края медуллярной пластинки приподнимаются и загибаются навстречу друг другу, вследствие чего пластинка превращается в продольный желобок (медуллярный желобок). В конце процесса края медуллярной пластинки сливаются друг с другом по линии, называемой спинным швом. Таким образом медуллярная пластинка превращается в спинномозговую трубку. Спинномозговая трубка отделяется по линии спинного шва от дорзально лежащего эктобласта и уже в этих ранних стадиях представляет следующие отделы: задний, более узкий, есть спинной мозг; передний, более широкий, - головной мозг. Головной мозг, образовавшийся заворотом головной пластинки, представляет три расширенные отдела, более или менее ясно отделенные друг от друга поперечно идущими перетяжками. Эти отделы отчасти уже были намечены при раскрытом состоянии медуллярной пластинки, они называются первичными мозговыми пузырями. Самый передний называется первичным передним мозгом или межуточным мозгом (thalamencephalon). Полость этого отдела называется третьим желудочком. Средний отдел шире других, называется средним мозгом (mesencephalon). Полость его, вследствие утолщения стенок пузыря, превращается впоследствии в узкий канал, называемый сильвиевым водопроводом (aquaeductus Sylvii). Задний отдел, постепенно суживаясь, прямо переходит в спинной мозг, вследствие чего его задняя граница не ясна. Этот отдел называется задним мозгом (myelencephalon), а полость его - четвертым желудочком.
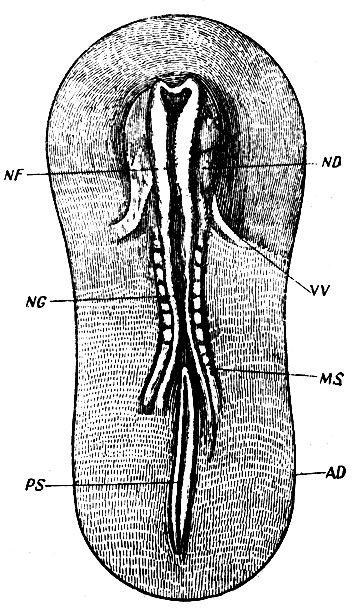
Рис. 2. Зародышевый диск курицы (AD). NG - медуллярная борозда, ограниченная медуллярными складками (NF); MS - сомиты; PS - первичная борозда; HD - головные утолщения; VV - желточные вены
Первичные мозговые пузыри лежат на прямом продолжении оси спинного мозга, и, следовательно, их можно рассматривать как передние расширенные отделы спинномозговой трубки. Три первичные мозговые пузыря гомодинамичны* некоторому отделу спинного мозга и отличаются от него более сильным развитием.
* (Гомодинамичными называются органы, гомологичные в пределах тела одного и того же животного.)
В дальнейшем развитии часть боковых стенок первичного переднего мозга выпячивается в виде двусторонне-симметричных пузырей - так называемых первичных глазных пузырей, и весь thalamencephalon загибается вентрально почти под прямым углом к оси заднего отдела спинномозговой трубки. Вследствие этого загибания дорзальная стенка первичного переднего мозга меняет свое горизонтальное положение на вертикальное и ложится почти параллельно плоскостям поперечных сечений зародыша. Позже дорзальная стенка thalamencephali выпячивается спереди в виде парных выростов. Эти выросты называются вторичным передним мозгом (prosencephalon) и представляют собой закладку полушарий головного мозга. Полости полушарий называются первым и вторым боковыми желудочками головного мозга, а проход, ведущий из thalamencephalon в боковые желудочки, называется монроевым отверстием (foramen Monroi). Передняя стенка foramen Monroi образована вертикально стоящей дорзальной стенкой thalamencephali, лежащей между полушариями, и называется концевой пластинкой (lamina terminalis).
В ранних стадиях у foramen Monroi собственно нет задней стенки, так как боковые желудочки прямо переходят в третий желудочек, но в более поздних стадиях боковые стенки thalamencephali сильно утолщаются. Эти сильно утолщенные отделы, или зрительные бугры (thalami optici), значительно суживают полость третьего желудочка в особенности спереди, вследствие чего в поздних стадиях foramen Monroi ограничивается сзади передними отделами thalami optici, между которыми остается вертикальная щель, ведущая из монроева прохода в третий желудочек. Вторичный передний мозг как парный вырост только дорзальной стенки thalamencephali не может быть сравним (гомологизирован) с отделами спинного мозга, так как представляет собою своеобразную и только местную диференцировку дорзальной стенки спинномозговой трубки.
В самых ранних стадиях средний эмбриональный лист распадается на серединный и два боковых отдела. Серединный отдел есть стержень хорды: он лежит непосредственно вентрально от медуллярной пластинки, соответственно осевой линии зародыша, и доходит спереди до передних отделов дна среднего мозгового пузыря. Следовательно, хорда продолжается и в головную область зародыша. Боковые отделы мезобласта состоят из парных пластинок. Серединные отделы этих пластинок, обращенные к хорде, более толсты и называются туловищными отделами; к бокам они постепенно утончаются и переходят в так называемые париетальные отделы мезобласта. Мезобластические пластинки непрерывно продолжаются до самых передних отделов головной области зародыша.
В несколько более поздних стадиях париетальные отделы мезобласта расщепляются на две пластинки. Дорзальная пластинка, прилегающая к эктобласту, называется кожноволокнистым листом (somatopleura); вентральная пластинка, прилегающая к эндодерме, называется кишечно-волокнистым листом (splanchnopleura). Полость, получившаяся путем расщепления париетальных отделов мезобласта, есть общая полость тела (coelom). Расщепления простираются медиально и в утолщенные туловищные отделы мезобласта, но не доходят до дорзомедиальных углов этих отделов. Общая полость тела (coelom) простирается непрерывно в головной отдел зародыша и доходит включительно до того места, где впоследствии образуется нижняя челюсть, т. е. приблизительно до задних отделов среднего мозга. Продолжение полости тела далее кпереди не вполне достоверно констатировано. При дальнейшем развитии туловища отделы мезобласта подвергаются сегментации, т. е. расщеплению на поперечные, метамерно расположенные отделы, называемые первичными сомитами. Полость, заключенная в каждом сомите, сообщается несколько суженным канальцем с широкой полостью, образовавшейся между splanchno- и somatopleura (coelom). Эти канальцы мы называем канальцами сомитов (Urwirbelcommunication Rabl, Nephrotom Rückert, Mesomer v. Wijhe). Канальцы сомитов остаются недолго; уже в ранних стадиях сомиты отделяются от плевральных пластинок, причем известные отделы канальцев сомитов преобразуются в канальцы органов выделения первичных почек (mesonephros). Другие отделы этих канальцев распадаются в скелетобластическую ткань. К этим процессам мы еще вернемся впоследствии. После полного отделения плевральных пластинок от сомитов вершина полости тела заканчивается несколько дорзальнее свода кишечника в месте серединного перегиба somato- в splanchnopleura.
При рассматривании зародыша сверху сомиты представляются в виде четырехугольных, более темных участков, лежащих по бокам хорды и отделенных друг от друга поперечными светлыми пограничными полосами. Первичные сомиты имеют поверхностное сходство с позвонками, вследствие чего они весьма неудачно и назывались прежде "первичными позвонками". С позвонками эти образования не имеют ничего общего; они представляют собою метамерные зачатки мышц, причем серединные отделы их при дальнейшем развитии распадаются в индиферентную, скелетообразующую ткань, не расчлененную на поперечные сегменты. Эта ткань окружает сплошной массой хорду и спинномозговую трубку, давая материал для их оболочек. Только в значительно позднейших стадиях дорзально и вентрально от хорды появляются в этой ткани парные, метамерно расположенные центры охрящевения (у Ichtyopsida). Дорзальные представляют зачатки верхних дуг позвонков, вентральные - нижних дуг. Значительно позже дуг образуются тела позвонков. Здесь важно также отметить, что метамерия дуг не совпадает с метамерией первичных сомитов, так как зачатки дуг появляются в местах, соответствующих границам между двумя следующими друг за другом сомитами. Каждый первичный сомит захватывает, таким образом, область двух скелетных сегментов. На взрослом животном несоответствие мускульной и скелетной метамерии выражается в том, что у рыб соединительнотканные перегородки между сегментально расположенными мышцами (миокоммы) прикрепляются к середине тела позвонка. Следовательно, каждая сегментальная мышца соответствует заднему отделу предшествующего и переднему кзади следующего позвонка. То же самое мы находим и в эмбриональных сомитах.
Эти факты выясняют также значение эмбриональной метамерии и в головной области позвоночных животных. Позже, чем в туловище, появляются сомиты в мезобласте головной области. Яснее всего они выражены у акул; относительно числа их существует большое различие во взглядах, но число сомитов не имеет тут особенно важного значения. Существенными являются следующие факты: 1) расчленение мезобласта головы на сомиты простирается на всю область, занимаемую впоследствии висцеральными дугами черепа, включая сюда и область челюстной дуги; 2) образование сомитов не стоит в связи с образованием жаберных мешков или щелей, так как линии, делящие сомиты друг от друга, начинаются с дорзомедиальных частей головного мезобласта, т. е. с области, в которую не простираются дорзальные концы жаберных щелей (Marschall); 3) продолжение общей полости тела в головную область зародыша и, еще более, факт расчленения мезобласта этой области на некоторое число сомитов указывают на примитивное состояние, при котором известный отдел головной области является менее отличным от отдела туловищной области, чем у самого примитивного взрослого животного, имеющего череп (т. е. низшего из Craniata); 4) в головной области зародыша прямо обозначаются два существенно отличные отдела: задний, в котором находится общая полость тела, сомиты, в котором происходит закладка, висцеральных дуг и в котором, как выше указано, на известном протяжении из туловищной области продолжается хорда, имеет много общего с туловищным отделом; передний отдел, в который полость тела не простирается, не имеет общих черт с туловищем. В этом переднем отделе лежат закладки органов зрения: и обоняния; 5) в головной области так же, как мы выше видели в туловищной, присутствие сомитов может указывать только на метамерную закладку мускулатуры, но никак не на метамерную закладку осевого скелета, т. е. черепа. И действительно, сомиты головной области дают прямо закладку мышц глазного яблока и мышц висцеральных дуг. Косвенно часть ткани этих сомитов участвует в образовании самих метамерно расположенных висцеральных дуг (челюстной, подъязычной и жаберных).
Черепная коробка возникает из нерасчлененных образований, и развитие ее существенно отличается от выше описанного развития позвоночного столба.
В общих чертах развитие черепа совершается следующим образом. Медиальные отделы сомитов распадаются на индиферентную эмбриональную ткань, которая сливается с нерасчлененным на сомиты мезобластом передней части головы. Эта ткань в виде сплошной нерасчлененной массы облекает основание мозга и простирается более или менее на его боковые и дорзальные отделы. В области среднего мозга эта ткань окружает передний (черепной) отдел хорды. Первые следы хряща возникают в этой ткани в виде двух пар удлиненных пластинок, лежащих у основания черепу. Задние пластинки лежат по бокам хорды и называются parachordalia. Они помещаются непосредственно под слуховыми пузырями, которые образовались путем врастания эктобласта. Передние пластинки называются trabeculae cranii и лежат на продолжении parachordalia. Они обхватывают самый передний конец хорды и проходят под глазными пузырями, доходя до закладок обонятельных органов, которые также образуются путем парных врастаний эктобласта в передне-вентральной части головного отдела зародыша. Следующая стадия развития состоит в том, что задние концы trabeculae cranii срастаются с parachordalia, и путем медиальных сращений из двусторонне-симметричных хрящей образуется одно нерасчлененное хрящевое образование - закладка основания черепной коробки, или так называемая базальная пластинка. Нетрудно убедиться, что базальная пластинка представляет несколько отделов, на которых резко выражено применение роста хрящей к трем парным органам чувств головы. Parachordalia и задние отделы trabeculae, сливаясь медиально, облекают хорду и растут дорзально, окружая дно слуховых пузырей. Таким образом получается лабиринтовый отдел базальной пластинки. Задние отделы trabeculae очерчивают под глазными пузырями овальное пространство, в котором помещается мозговой придаток (hypophysis cerebri). От боковых частей этого отдела начинается обрастание хрящом глазных пузырей и частей головного мозга, лежащих в области этих пузырей. Описанный отдел базальной пластинки называется орбитальным. Кпереди от орбитального отдела trabeculae cranii сливаются медиально и образуют так называемую этмоидальную пластинку, лежащую между органами обоняния.
Разращением боковых частей этой пластинки образуется этмоидальный отдел черепной коробки, включающей капсулы обонятельных органов. На сравнительно ранней стадии обрастание обонятельных органов хрящом выражается в развитии хрящевых отростков этмоидальнои пластинки, очерчивающих эти органы спереди и сзади. Передние отростки очень постоянны и называются соrnua trabecule.
Из этого краткого очерка развития черепа важно отметить следующее:
1. Никаких следов метамерии в начальных закладках черепной коробки мы не находим. Этим закладка черепной коробки существенно отличается от закладки хрящевых дуг позвоночного столба, которые с самого начала закладываются метамерно, соответственно числу будущих позвонков.
2. В начальной закладке черепа резко выражается применение этой закладки к трем парным органам чувств головы. Применение это вызывает с самых ранних стадий появление лабиринтового, орбитального и этмоидального отделов базальной пластинки. Те же отделы мы встречаем и в хрящевом черепе низших позвоночных животных, который тем примитивнее, чем яснее сохраняется в его общей форме первоначальная применяемость стенок к включенным в нее аппаратам органов чувств.
У менее примитивных форм применение стенок черепа к придаточным аппаратам (система висцеральных дуг и их мускулатура), а также и развитие различных отделов головного мозга изменяют примитивную архитектуру черепа. Таким образом примитивные отделы черепной коробки становятся менее ясными.
В связи со средним листом, именно с кишечно-волокнистой пластинкой этого листа, развивается сердце, причем, однако, так называемый эндотелий сердца, или эндокардий, получается из эндобласта. Мышечная стенка сердца образуется из кишечно-волокнистого листа. Полость околосердечной сумки есть вначале часть общей полости тела и отделяется от последней только в дальнейшем развитии. У круглоротых и многих Plagiostomi околосердечная сумка находится в прямом сообщении с полостью тела. У этих форм в течение всей жизни сохраняется отношение околосердечной сумки, общее зародышам всех позвоночных животных. Кровеносные сосуды развиваются в среднем листе и возникают как сливающиеся друг с другом лакуны или щели между клетками мезобласта. Часть клеток, выстилающих эти лакуны, превращается в форменные элементы крови, другая часть превращается в эндотелий, выстилающий внутреннюю поверхность сосудов. Часть форменных элементов крови, повидимому, образуется из эндобласта, выстилающего боковые отделы желтка. Описание кровеносной системы зародыша удобнее дать после описания главнейших процессов развития эндобласта.
В начале развития эндобласт плотно прилегает к желтку и в более поздних стадиях облекает всю его поверхность. При дальнейшем развитии головная и хвостовая области зародыша вместе с подлежащим эндобластом отделяются, от поверхности желтка, вследствие чего в головной и хвостовой области зародыша образуются два слепых, мешковидных выступа, внутренняя поверхность которых выстлана эндобластом. С боков зародыш тоже несколько поднимается над желтком, так что эндобласт образует соответственно продольной оси зародыша продольный жолоб. Этот жолоб есть дорзальный свод средней части кишечника. Все описанные отделы эмбрионального кишечника вентрально сообщаются прямо с эндобластическцм мешком, заключающим в себе желток, или, как у млекопитающих, с выстланной эндобластом полостью зародышевого пузыря. Последний вполне гомологичен желточному мешку других позвоночных.
Широкий канал, ведущий из полости эмбрионального кишечника в полость желточного пузыря, называется ductus omphalo-entericus. При дальнейшем развитии эмбриональный кишечник все более и более отшнуровывается от желточного пузыря, вследствие чего ductus omphalo-entericus становится постепенно уже.
В головной части кишечника у всех позвоночных развиваются глоточные, или жаберные, мешки. Эти мешки представляют собой слепые выросты эндобласта, растущие по направлению к вентральному эктобласту головы, с которым они, наконец, сливаются. У большинства позвоночных глоточные мешки в местах своего сращения с эктобластом вскрываются наружу так называемыми висцеральными щелями. У млекопитающих эктобластические пластинки, замыкающие жаберные мешки, не прорываются; следовательно, у них дело не доходит до образования глоточных, или висцеральных щелей. У всех позвоночных между глоточными мешками образуются метамерно расположенные мезобластические закладки в виде дуг из уплотненной ткани, называемые эмбриональными висцеральными дугами. У рыб образуется 6 глоточных мешков и 7 эмбриональных висцеральных дуг, расположенных следующим образом: передняя дуга лежит кпереди от первой глоточной, или висцеральной, щели (так называемое брызгальце, spiraculum) и называется челюстной дугой, вторая называется подъязычной (гиоидной) и лежит кзади от брызгальца и кпереди от 1-й жаберной щели (2-я глоточная щель), остальные дуги называются жаберными и считаются назад от 1-й, как 1-я -5-я, Пятая дуга лежит позади 5-й жаберной щели (6-й глоточный мешок). В каждой из эмбриональных висцеральных дуг рыб, кроме мышц, сосудов и нервов, развиваются еще скелетные части - висцеральные дуги черепа: 1-я, или челюстная, 2-я, или подъязычная, и 5 жаберных. Сравнительная анатомия рыб указывает, что у огромного большинства современных рыб число висцеральных дуг редуцировано на две дуги. Дело в том, что у акул Hexanchus и Chlamydoselachus имеются не 5, а 6 жаберных дуг, и у акулы Heptanchus даже 7. По своему строению, эти акулы являются низшими среди современных.
У зародыша человека закладывается с каждой стороны только 5 эмбриональных висцеральных дуг: челюстная, подъязычная и 3 жаберные дужки. Задняя жаберная дужка при рассматривании зародыша снаружи не видна, но может быть прослежена на разрезах в виде поперечного валика, вдающегося в полость глотки. Так как для этого валика существует особая дуга аорты, то очевидно, что это образование представляет собой третью жаберную дугу. Таким образом у зародыша высшего млекопитающего сравнительно с высшими рыбами произошла редукция еще двух задних эмбриональных висцеральных дуг. Скелетные части, развивающиеся в висцеральных дугах млекопитающих, следующие: из 1-й образуются небно-квадратный хрящ (зачаток верхней челюсти), меккелев хрящ (зачаток нижней челюсти) и позднее, за их счет, две слуховые косточки - наковальня и молоточек. Во 2-й развивается 3-я слуховая косточка - стремя, шиловидный отросток (processus styloideus) и соединенный с ним связкой малый рог подъязычной кости. В 3-й дуге развивается большой рог подъязычной кости, тогда как ее тело ( образуется из особого хряща, связывающего висцеральные дуги вентрально. Остатки 4-й и: 5-й висцеральных дуг идут на развитие щитовидного хряща.
Теперь возвратимся к развитию кровелосной системы.
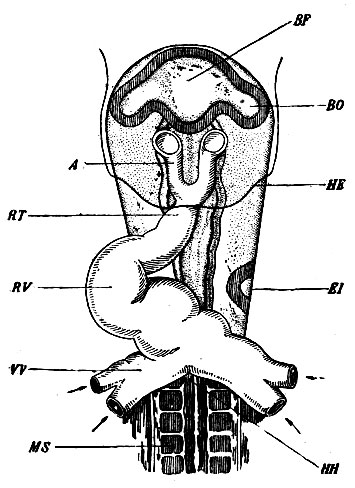
Рис. 3. Закладка сердца. RV - желудочковая часть сердца; RT - передний придаток сердца с корнями аорты (A); VV - желточные сосуды; MS - сомиты; BF - передний первичный пузырь головного мозга; ВО - зачаток глаза; El - зачаток уха; НН - спинной мозг
В начальных стадиях развития сердце у зародыша человека представлено двумя желобками, лежащими по бокам зачатка пищеварительной полости. Каждый желобок представляет собой выпячивание эндобласта, охваченное соответственным выпячиванием спланхнической пластинки мезобласта. Когда зачаток кишечника свертывается в трубку, парный зачаток сердца, представленный двумя развившимися из желобков трубочками, ложится вентрально, трубочки срастаются в одну, и теперь сердце представлено искривленной известным образом широкой трубкой, которая лежит под головной частью кишечника и сильно выпячивается наружу. Сердце начинает сокращаться тотчас после его образования и ранее, чем в нем появляется уловимая для нас гистологическая диференцировка на мышечную и нервную ткань. Передний конец сердечной трубки продолжается в непарный сосуд - truncus arteriosus, на высоте задней висцеральной дужки. Truncus arteriosus распадается на две симметричные ветви, которые идут кпереди, затем дугой охватывают боковые части кишечника и поднимаются к его дорзальной стороне, где и дают кпереди пару ветвей. Эти ветви несут кровь к головному мозгу и называются сонными артериями (a. carotides). Главные стволы загибаются назад и идут вентрально от хорды к хвостовой части зародыша. Дугообразный отдел этих сосудов, огибающий головной отдел кишечника, называется первой дугой аорты; их прямое продолжение - парные сосуды, идущие к хвостовой части зародыша, называются первичными аортами. Вначале аорты парны по всей длине зародыша, но вскоре те отделы их, которые лежат в срединной части туловища зародыша, сливаются в один непарный сосуд. Отделы, остающиеся парными, дают вправо и влево артерии, несущие кровь к стенкам желточного мешка, где они распадаются в сильно развитые сети лакун. Эти важные сосуды называются желточными артериями (arteriae omphalo-mesentericae). Конечные отделы первичных аорт несут кровь к хвостовой части кишечника и называются пупочными артериями (a. umbilicales s. allantoideae).
В головной части кишечника по мере развития висцеральных дуг возникают новые дугообразные сосуды, лежащие кзади от первой дуги аорты; они соединяют начальные артерии с первичными аортами и идут вдоль висцеральных дуг. Эти сосуды называются 2-й, 3-й и т. д. дугами аорты. Число их соответствует числу закладывающихся эмбриональных висцеральных дуг. У низших позвоночных две передние пары рано исчезают. У высших их закладывается 5 пар, из которых 5-я соответствует по положению 6-й, а собственно 5-я не развивается. Постоянная аорта развивается у них за счет 4-й пары.
Задний отдел сердечной трубки называется венозной пазухой (sinus venosus). Он образуется слиянием двух больших парных вен, несущих кровь из желточного пузыря (venae omphalo-mesentericae). Ближе кпереди в него впадают справа и слева два коротких поперечно идущих венозных ствола, которые называются правым и левым протоком Кювье (ductus Cuvieri). Протоки Кювье несут венозную кровь собственно из тела зародыша. Каждый проток слагается из слияния двух вен: передней кардинальной (v. cardinalis anterior, или первичная v. jugularis) и задней (v. cardinalis posterior), несущей кровь из туловищной и хвостовой области. У человека из передних развиваются яремные вены, из задних - непарная (v. azygos) и полунепарная (v. hemiazygos), а сама венозная пазуха ассимилируется правым предсердием.
Придатки и оболочки зародыша. У большинства рыб и амфибий оплодотворение яиц происходит или после их отложения, или незадолго до их отложения. Такие формы называются яйцеродными (Ovipara). Яйца рыб и амфибий развиваются в воде и, кроме студенистых оболочек и zona radiata, т. е. собственно оболочек яйца, других оболочек не имеют (Anamnia).
Единственный зародышевый придаток Anamnia представлен желточным пузырем, который сильно развит у рыб с меробластическими яйцами и слабо у форм с голобластическими яйцами. У последних он сводится на ступень раздутого так называемыми желточными клетками среднего отдела кишечника.
В течение всего развития зародыш поглощает кислород окружающей среды и выделяет углекислоту, т. е. дышит. Кроме того, он питается, поглощая постепенно желток, заключенный в желточном пузыре. У Anamnia дыхание совершается всей поверхностью тела зародыша, причем газы диффундируют через яйцевые оболочки. В более поздних стадиях дыхание, а также и питание, совершается посредством сильно развитой сети сосудов стенок желточного мешка (vasa omphalo-mesentericae).
У птиц оплодотворенное яйцо откладывается в очень ранних стадиях развития. У многих пресмыкающихся зародыш проходит почти полное развитие до отложения яйца - такие формы называются яйцеживородными (Ovoviviраrа).
У птиц и пресмыкающихся из тканей зародышу образуется система оболочек придатков, которых нет у Anamnia. На ранних стадиях развития эктобласт в сопровождении листка somatopleura начинает подниматься вокруг зародыша в виде складок (головная складка, две боковые и хвостовая). Последние растут навстречу друг другу к одному центру и срастаются своими вершинами над зародышем. Вследствие этого весь зародыш, за исключением желточного пузыря, заключается в оболочку, полость которой наполняется серозной жидкостью. Эта оболочка называется amnion. Как следствие развития амниона прямо получается общая внешняя, оболочка (serosa), которая состоит из соединившихся над амниотическим швом наружных пластинок амниотических складок. Вскоре амнион совершенно отделяется по линии шва от серозной оболочки. Последняя покрывает зародыш, одетый амнионом, и продолжается на желточный мешок. Само собой понятно, что так как в образовании складок амниона участвовали эктобласт и соматоплевра, пространство, покрытое serosa, прямо переходит в общую полость тела (coelom) зародыша. Физиологическое значение амниона сводится, как кажется, к защите зародыша от внешних воздействий (толчки, передающиеся через слой жидкости, ослабляются; другими словами, амнион можно сравнить с подушкой, окружающей зародыш).
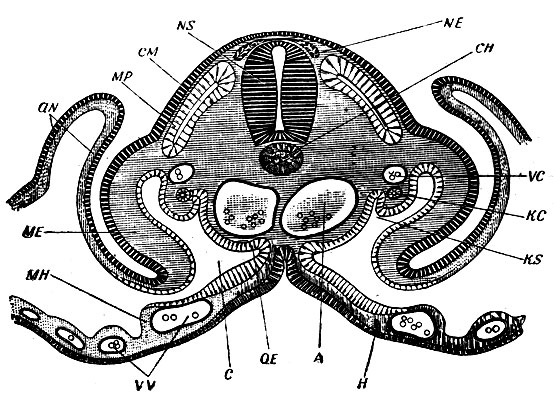
Рис. 4. Поперечный разрез через зародыш курицы, когда закладывается амнион (aN). NS - спинной мозг; NE - зачатки нервов; СМ - сомиты; СН - хорда; А - аорта; КС, KS - зачатки почек; С - общая полость тела; ME - соматоплевра; МН - спланхноплевра; Н - эндодерма
После образования амниона в хвостовой части кишечника появляется вентрально направленный дивертикул или вырост, который изнутри одет эндобла стом, а снаружи кишечноволокнистым листом мезобласта (splanchnopleura). Этот дивертикул растет в пространство, лежащее между амнионом, serosa и желточным пузырем, т. е. в полость тела, лежащую вне пределов собственно зародыша. Вершина дивертикула скоро превращается в пузырь, который называется аллантоисом (allantois), а его стебель, ведущий в полость хвостовой части кишечника, - urachus. Пупочные артерии (аа. umbilicales) разветвляются в стенках аллантоиса и дают сильно развитые капиллярные сети, из которых кровь собирается в две пупочные вены (vv. umbilicales), впадающие одним общим стволом в левую желточную вену (v. omphalo-mesenterica). В дальнейшем развитии аллантоис сплющивается и, сильно разрастаясь, вместе с пластинкой serosa плотно прилегает к пористым оболочкам яйца. Ткань serosa, которая по устарелой терминологии называется также субзональной оболочкой, на местах сращения с аллантоисом редуцируется. У пресмыкающихся и птиц аллантоис составляет исключительно орган дыхания зародыша. Диффузия газов совершается сквозь тонкие стенки капиллярных сосудов аллантоиса и пористые оболочки яйца. В яйцах, покрытых лаком или сильно загрязненных, зародыш погибает от асфиксии.
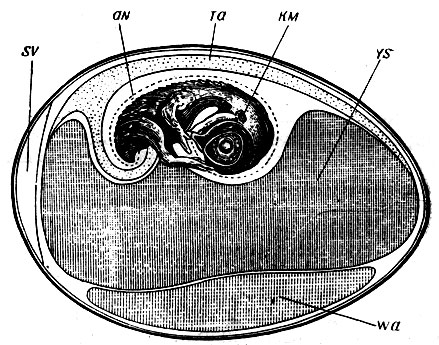
Рис. 5. Продольный разрез куриного яйца. Зародыш с амнионом (aN), аллантоисом (Та) и желточным мешком (VS). Wa - остатки белка; КМ - зачаток уха
Как мы выше видели, яйца высших млекопитающих почти лишены питательного желтка. Дробление - голобластическое. Начальные стадии развития ведут к образованию объемистого пузыря (зародышевый пузырь), состоящего из эктобласта с скоплением клеток у одного полюса, откуда постепенно распространяется эндобласт. Последний листок обрастает пузырь (под эктобластом) в течение хода развития зародыша. Из только что упомянутого клеточного скопления на одном из полюсов пузыря (area embryonalis) происходит развитие зародыша. В главных чертах оно происходит так же, как это описано для птиц. По мере развития, зародышевый пузырь, как сказано, сильно увеличивается в объеме и при этом наполняется серозной жидкостью, транссудирующей сквозь его стенки из полости матки. Процесс образования зародышевого пузыря млекопитающих сводится к процессу развития желточного пузыря, причем недостающий у млекопитающих питательный желток заменяется для ранних стадий развития жидкостью, транссудирующей из полости матки. У млекопитающих вся функция питания зародыша выпадает в начальном развитии на слизистую оболочку матки, а впоследствии на кровеносную систему матери.
Придатки зародыша в различных отрядах млекопитающих представляют значительные различия во взаимных отношениях, но эти различия сводятся к частностям. У всех млекопитающих есть serosa, амнион, желточный мешочек (зародышевый пузырь или желточный пузырь) и аллантоис. Развитие этих придатков, кроме указанных различий в образовании желточного мешочка, совершается в общих чертах так же, как у птиц. Существенными являются различия в отношении аллантоиса к слизистой оболочке матки в различных группах млекопитающих.
У двуутробок после образования амниона на поверхности serosa образуются складки, которые плотно внедряются в складки слизистой оболочки матки, но не срастаются с ними. Сосуды желточного мешочка получают сильное развитие; аллантоис очень мал и не срастается с поверхностью serosa; следовательно, хотя его сосуды и развиты, он не может играть важной роли в процессах дыхания зародыша.
У высших млекопитающих питание и дыхание зародыша совершается через посредство аллантоиса и сосредоточивается в тех участках этого зародышевого придатка, где он, срастаясь с serosa, представляет сильно развитые ворсины, внедряющиеся в слизистую оболочку матки. Поверхность аллантоиса, занятая этими ворсинами, называется зародышевой плацентой (placenta foetalis), а более или менее сильно измененная слизистая оболочка матки, включающая ворсины аллантоиса, - плацентой маточной (placenta uterina). Оба образования, состоящие друг с другом в тесной связи, вообще называются последом (placenta).
Еще до полного развития аллантоиса поверхность serosa покрывается небольшими ворсинами. В этом виде серозная оболочка зародыша известна под названием chorion frondosum. В последующем развитии на поверхности хориона обозначаются два различных отдела. На том из них, с которым срастается аллантоис, развитие ворсин идет прогрессивно далее, и они вступают в связь с слизистой оболочкой матки; на остальной периферии хориона ворсины постепенно атрофируются. Этот отдел называется cborion laeve. По мере развития аллантоиса и его связей со стенками матки, желточный мешочек (зародышевый пузырь) сильно уменьшается в объеме, и желточный проток (ductus omphalo-entericus) превращается в длинный тонкий стебель. Этот стебель плотно прилегает к стеблю аллантоиса (urachus), и оба образования, вследствие замыкания стенки полости зародыша, обхватываются корнем амниотического пузыря. Таким образом, получается тот орган, который называется пуповиной (funiculus umbilicalis), на котором зародыш как бы подвешен к плаценте. Пуповина состоит, следовательно, из следующих образований: снаружи она облекается корнем амниона, внутри включает желточный проток с резорбированным желточным пузырем и редуцированными периферическими отделами желточных сосудов. Кроме того, пуповина содержит стебель аллантоиса с сильно развитыми пупочными артериями и венами, через которые и совершается плацентарное кровообращение зародыша.
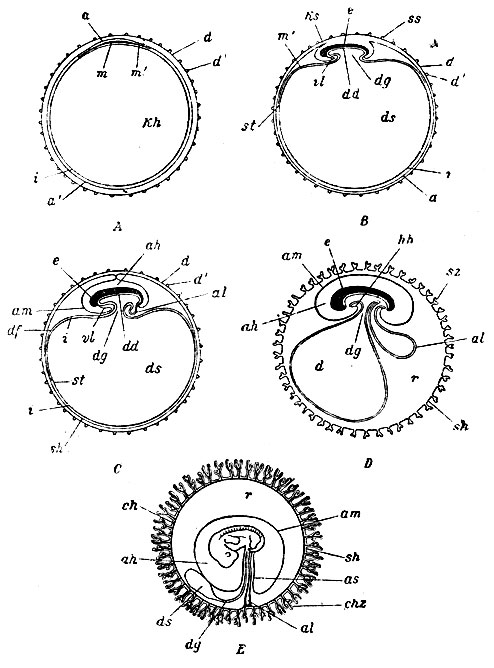
Рис. 6. Пять схематичных разрезов, иллюстрирующих образование зародышевых придатков млекопитающего. На рисунках зародыш представлен в продольном разрезе. А - яйцо с zona pellucida, бластодермическим пузырьком и зародышевой областью; В - яйцо при начале образования желточного мешочка и амниона; С - яйцо с почти замкнувшимся амнионом; D - яйцо с сосочками на оболочке и большим аллантоисом; рот и заднепроходное отверстие образовались; Е - яйцо, в котором мезодерма аллантоиса распространилась по всей внутренней поверхности оболочки яйца, образовав вместе с нею хорион; полость аллантоиса исчезла, а - эктодерма зародыша; а' - эктодерма внезародышевой части бластодермического пузырька; ah - полость амниона; al - аллантоис; am - амнион; ch - хорион; chz - его сосочки; d - zona radiata; d' - ее отростки; dd - эктодерма зародыша; df - сосудистая область; dg - стебелек желточного мешочка; ds - его полость; е - зародыш; hh - околосердечная полость; i - внезародышевая эндодерма; Kh - полость бластодермического пузырька; Ks - головная складка амниона; m - зародышевая мезодерма; m' - внезародышевая мезодерма; r - промежуток между хорионом и амнионом; sh - серозная оболочка; ss - хвостовая складка амниона; st - sinus terminalis; vl - брюшная стенка тела
Ворсины chorion frondosum могут быть рассеяны на большой поверхности serosa без определенного порядка. В таком случае места, лишенные ворсин (chorion laeve), обыкновенно находятся у полюсов оболочки. Такое распределение ворсин обусловливает форму плаценты, которая называется плацентой рассеянной (placenta diffusa). Другая форма плаценты происходит вследствие скопления ворсин в виде разбросанных дисковидных групп, называемых котиледонами. Такая плацента называется плацентой котиледонарной (рlacenta cotyledonaria). Число котиледонов вариирует от нескольких до сотни. При плаценте диффузной и котиледонарной связь слизистой оболочки матки с ворсинами хориона довольно слаба. Ворсины входят в углубления гипертрофированной слизистой оболочки и могут быть легко вытянуты из них, что и происходит, действительно, при родах. Стенки углублений слизистой оболочки матки выстланы цилиндрическим эпителием, который выделяет особую жидкость, похожую на молоко. Это маточное молоко всасывается ворсинами chorion frondosum и через сосуды аллантоиса приносится к зародышу. При таких формах плаценты слизистая оболочка матки не отпадает при родах (точнее, после родов), и млекопитающие с таким устройством плаценты называются млекопитающими без отпадающей оболочки (Nondeciduata).
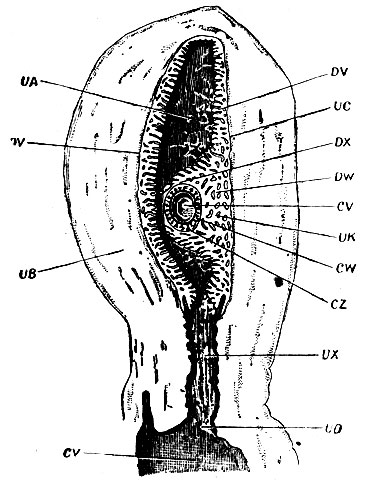
Рис. 7. Матка человека с яйцом, в продольном разрезе, приблизительно на тринадцатый день. CV - полость бластодермического пузыря; CW - стенка его; CZ - ворсинка хориона; DV - decidua vera; DW - d. serotina; DX - d. reflexa; UA - полость матки; UX - шейка матки
У всех других млекопитающих слизистая оболочка матки столь сильно изменена и ее связь с зародышевой плацентой столь тесна, что большая или меньшая часть ее выделяется при родах с зародышем,- другими словами, поверхность матки при родах обнажается. Млекопитающие с такими формами плаценты называются млекопитающими с отпадающей оболочкой (Deciduata).
У млекопитающих с отпадающей оболочкой плацента является в трех главных формах. У одних (хищные, слоны) плацента окружает серозный пузырь в виде пояса и оставляет свободными его полюсы, это - поясовидная плацента (placenta zonaria). Другая главная форма - плацента дисковидная (placenta discoidea), в которой chorion frondosum сосредоточивается на известном дисковидном участке серозной оболочки, как это существует у обезьян и человека. Наконец, у ленивцев и лемуров имеется колоколовидная плацента (placenta a cloche), когда ворсины образуют кольцо, окружающее некоторое пространство, к центру которого постепенно исчезают.
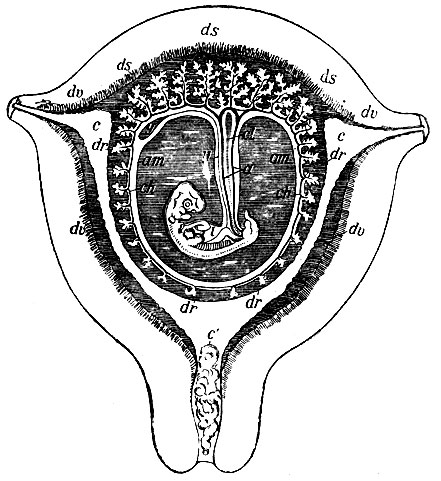
Рис. 8. Схематический разрез беременной матки человека на 7-8-й неделе. at - стебель аллантоиса; am - амнион; с - полость матки; с' - слизистая пробка в шейке матки; ch - хорион; dr - decidua reflexa; ds - d. serotina; du - d. vera; i - кишка зародыша; u - пупочные артерии; у - желточный мешок; у' - его стебель
При плаценте дисковидной, как и при поясовидной, в слизистой оболочке матки происходят сильные изменения. При первой изменения эти состоят в следующем: серозный пузырь прилегает к известному месту слизистой оболочки матки, которая называется decidua serotina; здесь и развивается материнская (или маточная) плацента. Остальные части слизистой оболочки сильно гипертрофируются и получают название decidua vera. Кроме того, края decidua serotina обрастают со всех сторон серозный пузырь и получают название decidua reflexa. В слоях decidua serotina, прилегающих к chorion frondosum, т.е. к зародышевой плаценте, сосуды слизистой оболочки превращаются в расширенные кровеносные полости, или лакуны, выстланные эндотелием. По направлению к этим лакунам растут ворсины зародышевой плаценты, одновременно с чем эпителий, покрывающий эти ворсины, атрофируется. Ворсины врастают в лакунарные пространства и таким образом окружаются кровью стенок матки. Следовательно, капиллярная сеть chorii frondosi, т.е. разветвления сосудов аллонтоиса, отделяется тонкой эндотелиальной стенкой от крови, циркулирующей в лакунах маточной плаценты.
Таким образом, в плаценте устанавливаются самые благоприятные условия для обмена питательных веществ и газов между кровеносной системой зародыша (плода) и матери.
При использовании материалов сайта активная ссылка обязательна:
http://anfiz.ru/ 'Анатомия и физиология человека'