Исходная гипотеза организации клеточных механизмов замыкания
Известно, что многократное предъявление на фоне анодной поляризации поверхности двигательной коры звуковых стимулов ведет к установлению на клеточном уровне относительно устойчивой связи между следами поляризующего тока и сенсорного раздражения, которая в последействии поляризации воспроизводится одной звуковой стимуляцией (Morrell, 1961b). По нашим данным, такая связь может быть выработана как при суммарной поляризации поверхности двигательной коры, так и при локальной поляризации отдельной клетки через регистрирующий микроэлектрод. В том и другом случае значительная часть нейронов после выключения поляризующего тока была способна дифференцировать звуковой раздражитель, многократно предъявленный на фоне поляризации, и раздражитель той же модальности, но другой частоты, который в аналогичных условиях столь часто не предъявлялся, на протяжении до 14-30 мин (М. Я. Рабинович, Ф. В. Копытова, 1969; Ф. В. Копытова, М. Я. Рабинович, 1970). Феномен носит в себе черты простых сенсо-сенсорных ассоциаций, но в отчетливой форме имитирует механизмы "сцепления" следов двух гетерогенных стимулов и в известной степени процесс афферентной специализации условной реакции.
Не требуется, вероятно, дополнительных пояснений, что понятие "сцепление" следов условного и безусловного стимулов предполагает какие-то специфические ультраструктурные и химические преобразования в цитоплазме нейрона, как это постулирует концепция конвергентного замыкания П. К. Анохина (1968). По-видимому, именно гетерохимической организацией синаптических входов для сигнального (сенсорного) и подкрепляющего (биологически значимого) стимулов и особыми химическими свойствами нейромедиатора (или медиаторов) каждого из входов можно сейчас удовлетворительно объяснить столь различные роли условной и безусловной афферентации в становлении устойчивых связей между ними.
Вероятная гипотетическая схема результатов взаимодействия синаптических входов с различными свойствами нейромедиаторов для сигналов сенсорной и биологической модальностей представлена на рис. 1. Сочетания стимулов двух сенсорных модальностей, например звука и света (обозначенных б1 и б2), приводят к образованию весьма кратковременной и легко распадающейся связи типа ранее упомянутых сенсо-сенсорных ассоциаций (см. рис. 1), хотя при этом, что важно отметить, также имеет место воспроизведение первым в сочетании стимулом некоторых особенностей второго. Сочетания каждого из сенсорных стимулов (б1 или б2) с биологически значимым подкреплением (а), нейромедиация которого, по-видимому, опосредована активностью структур и путей мозга (Т. А. Леонтович, 1968: Shute, Lewis, 1966; Г. Джаспер, 1967; Ungerstedt, 1971), причастных к мотивационно-эмоциональным факторам поведения (П. К. Анохин, 1968; П. В. Симонов, 1970; К. В. Судаков, 1971; Ю. А. Макаренко, 1972), приводит к выработке устойчивой связи со всеми свойствами условного рефлекса (см. рис. 1, 2).
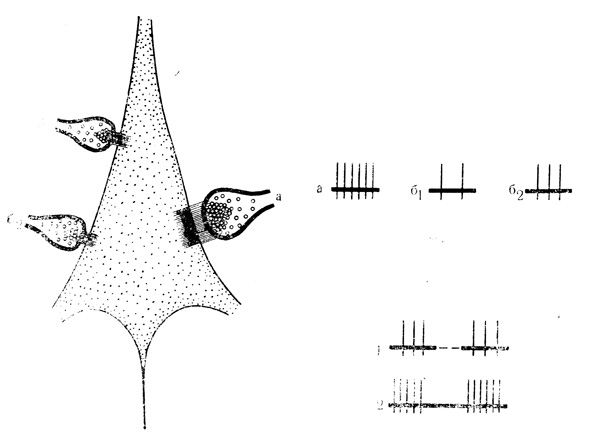
Рис. 1. Вероятная схема взаимодействия синаптических входов с гетерохимической организацией для стимулов биологической (а) и сенсорных (б1 и б2) модальностей. Различные свойства входов для сенсорных и биологически значимых сигналов условно изображены в виде размеров синапса и его 'активных зон'. Дополнительные пояснения в тексте. В основу положена схема последовательных стадий возбуждения синапса по С. А. Саркисову и Н. Н. Боголепову (1967)
В настоящее время известно достаточно много физиологически активных веществ, которым с большим или меньшим основанием приписывается роль медиаторов и модуляторов синаптической передачи и действие которых на субсинаптическую мембрану нейрона оценивается главным образом по возбудительной или тормозной реакции клетки (Bloom, 1969; Hubbart, 1970; Euler, 1971; Krnjevic, 1971). Между тем для понимания особых свойств медиации подкрепляющего действия на нейрон необходимы новые подходы и прежде всего идентификация нейромедиаторов по их способности фиксировать связи между стимулами и длительно сохранять эти связи.
Вместе с тем на данном этапе наших знаний о природе условного рефлекса важно представить себе возможную организацию клеточных механизмов замыкания, которые в конечном счете реализуют устойчивые связи между следами сочетаемых стимулов, скорее всего сложившиеся в результате молекулярных преобразований в пределах цитоплазмы нейрона.
При выработке условных связей на уровне нейронов механизмы "сцепления" следов условного и безусловного раздражителей проявляют себя при пропусках в отдельных сочетаниях подкрепляющего стимула (Л. Л. Воронин, Р. Г. Кожедуб, 19716; Е. Л. Полонская, М. Я. Рабинович, 1972) или при пропусках сочетания в целом в условном рефлексе на время, в той его модификации, которая принята в нашей лаборатории (Ф. В. Копытова, М. Я. Рабинович, 1967; Ф. В. Копытова, Л. К. Куликова, 1970; Ф. В. Копытова, Ю. С. Медникова, 1972).
Характерная структура следовых разрядов на подкрепление при его пропуске в сочетании, воспроизводящая особенности ответа на реальный раздражитель, интенсивность таких разрядов, часто превышающая ответ на реальный стимул, появление их, как правило, в пределах интервалов отставления подкрепляющего стимула в предшествующих сочетаниях, позволяют рассматривать такую следовую активность, как явление опосредованное синаптическим действием, т. е. вызванное выбросом медиатора из пресинаитических терминалей (см. рис. 42, А). Еще лучше это видно при внутриклеточной регистрации, когда следовая реакция на подкрепление ничем по существу не отличается от ответа клетки на реальный безусловный стимул: такого же характера возбуждающие постсинаптические потенциалы, генерирующие импульсные разряды у нейронов с активизирующими условными реакциями (см. рис. 42, Б), и аналогичные деполяризационные колебания с прекращением генерации потенциалов действия у нейронов с тормозными условными реакциями (см. рис. 26).
Особенно демонстративно пресинаптические механизмы следовых реакций нейронов проявляются в серии последовательных пропусков сочетаний в целом при выработке условных рефлексов на время, когда в фиксированных этими сочетаниями временных интервалах выступают чисто следовые реакции на условный и безусловный стимулы, воспроизводящие структуру установившейся между ними связи (см. главу V, с. 166) Внутриклеточные исследования нейронов двигательной коры при выработке аналогов условного рефлекса на время, проведенные Л. Л. Ворониным (19716), показали, что следовые деполяризационные колебания, генерирующие потенциалы действия в ритме, предшествовавшего электрокожного раздражения конечности, по своей длительности, временному течению и реакциям на действие поляризующего тока не отличаются от постсинаптических потенциалов, вызванных реальной стимуляцией (см. главу V, рис. 51).
Предварительное, весьма сжатое изложение относящихся сюда фактов дает основание полагать, что регистрируемые па клеточном уровне условные связи опосредованы синаптическим действием. Необходима, следовательно, нейрофизиологически приемлемая схема, которая позволила бы объяснить, какими механизмами могут быть воспроизведены нейроном оба компонента этой связи действием одного сигнального стимула при пропусках подкрепления или вообще без какой-либо внешней стимуляции (т. е. действием пока неизвестных факторов) при воспроизведении условных реакций на время.
Такая гипотетическая схема, представленная на рис. 2, предусматривает, что синаптические входы для условного и безусловного стимулов расположены на пресинаптической клетке в цепи нейронов (ее удобнее называть "входной" клеткой; рис. 2, I), в пределах которой оба стимула взаимодействуют и где по мере сочетаний формируются молекулярные механизмы их "сцепления". Действием условного и безусловного стимулов сложившаяся ассоциация передается на постсинаптический ("выходной"; см. рис. 2, II) нейрон, но уже по одному общему пути и для сигнального, и для подкрепляющего раздражителей. Общий для всей ассоциации в целом синаптический вход дублируется и модулируется системами вставочных нейронов возбуждающего или тормозного действия в зависимости от активирующей или тормозной формы вырабатываемой условной реакции. В качестве примера на рис. 2 приведена схема формирования и передачи активирующей условной связи.
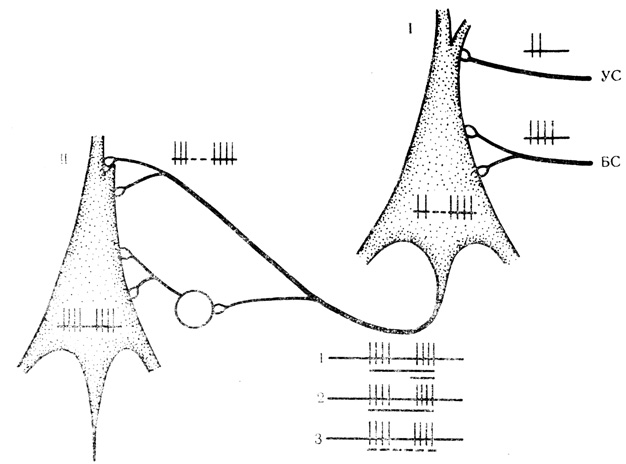
Рис. 2. Гипотетическая схема передачи условной связи, сложившейся в процессе сочетаний, с 'входного' нейрона (I) на 'выходной' нейрон (II) по одному общему пути. Клетка круглой формы - вставочный нейрон возбуждающего действия. На осциллограммах - различные формы условной активности, которые возможно регистрировать с 'выходного' нейрона: 1 - обычная реакция на условный (УС) и безусловный (БС) стимулы, обозначенные длинной и короткой линией под осциллограммой; 2 - воспроизведение следовых разрядов на БС при его пропуске в сочетании; 3 - воспроизведение следовых разрядов на УС и БС при пропуске всего сочетания (обозначено пунктиром) в условных реакциях на время. Дополнительные пояснения в тексте
Особо следует подчеркнуть, что деление нейронов на "входные" и "выходные" отнюдь не означает новой классификации. Эти обозначения относятся только к двум клеткам, искусственно вычлененным из сложных цепей нейронов, составляющих отдельные звенья условного рефлекса. В действительности каждый "входной" нейрон может служить "выходным" для каждого из предыдущих в цепи клеток и соответственно проявлять все формы реализации условных связей при регистрации его активности. В то же время каждая "выходная" клетка в свою очередь выполняет обязанности "входной" по отношению к последующим в цепи нейронам, если она располагает эффективно функционирующими входами для сочетаемых стимулов.
Более того, синаптические входы у "выходных" нейронов, организованных принципиально так же, как и клетки с "входными" функциями, способствуют дополнительной фиксации и совершенствованию условной связи, оба компонента которой были переданы сюда по общему для них пути. После такой дополнительной "обработки" в пределах "выходного" нейрона упроченная связь может быть передана на эффекторные звенья условного рефлекса одним сигнальным раздражителем, действие которого воспроизводит всю структуру поведенческого акта.
При этом химическая организация синаптических аппаратов общего пути может оказаться иной, чем у синаптических входов для сигнального и подкрепляющего стимулов. Эти различия могут проявить себя в латентных периодах реакции клетки, длительности эффектов последействия, которые установлены для разных нейромедиаторов при их микроионофоретической аппликации (В. В. Шерстнев, 1972; Krnjevic, 1971) и даже в механизмах деполяризующего действия, которые оказались иными, например, у ацетилхолина по сравнению с другими возбуждающими нейромедиаторами (Krnjevic е. а., 1971).
Таким образом, экспериментатор, вырабатывая условную связь на клеточном уровне, в конечном счете всегда регистрирует ее у нейронов с преобладанием "выходных" функций, как это вытекает из предложенной гипотезы формирования и передачи сложившихся интеграции в пределах популяции клеток. Такая схема удовлетворительно объясняет возможность наряду с обычной реакцией на сочетаемые стимулы воспроизвести у того же нейрона следовой разряд на пропущенное подкрепление или оба компонента (сигнальный и подкрепляющий) условной связи при пропусках всего сочетания в целом в условных реакциях на время (см. рис. 2, осциллограммы 1-3).
Сходные предположения можно усмотреть в обзоре Doty (1969), который детально рассмотрел вероятные пути и механизмы условных двигательных рефлексов, выработанных сочетаниями электростимуляции сенсорных и моторных зон мозга. По мнению Doty, такой "бессодержательный" стимул, как электроток, не мог бы вызвать сложный поведенческий акт, если бы реализующая его популяция нейронов не содержала в себе сложившуюся пространственно-временную структуру этого акта и не находилась по меньшей мере на одно синаптическое переключение дальше по отношению к месту приложения сигнального электростимула.
Потребность в постулировании механизмов, подобных описанному, возникает уже на этапе конвергенции на клетке стимулов различной афферентной природы. Об этом свидетельствуют внутриклеточные исследования Л. Л. Воронина и Л. И. Таненгольца (1969), результаты которых послужили авторам основанием предположить, что в определенной ситуации (хлоралозовый наркоз) первичное взаимодействие таких раздражителей, как звук, свет и электрокожное раздражение конечности, осуществляется на каких-то предшествующих клетках, после чего они приходят к исследуемым нейронам двигательной коры по одному общему для всех сигналов пути.
При использовании материалов сайта активная ссылка обязательна:
http://anfiz.ru/ 'Анатомия и физиология человека'