7. Негроидно-австралоидная раса
Большинство рас и групп антропологических типов тропической зоны образует экваториальную, или негроидно-австралоидную, большую расу, которая, как мы видели, естественно распадается на две малые расы: африканскую, или негроидную, и океанийскую, или австралоидную (рис. 48 и 49).

Рис. 48. Меланезиец с острова Новая Каледония. (Меланезийская группа типов австралоидной малой расы.)

Рис. 49. Меланезиец с Соломоновых островов. (Меланезийская группа типов австралоидной малой расы.)
Если бы мы пожелали сравнить африканских негров с австралоидами, то, наряду со многими чертами иногда поразительного сходства, мы заметили бы и заметные различия. Прежде всего волосяной покров на теле у негров слабый, нередко почти отсутствует, а у австралийцев, большинства меланезийцев и папуасов он очень заметно развит. На голове негров волосы завиваются сильнее, чем у папуасов или меланезийцев, у которых дети родятся с волнистыми волосами; у австралийцев волнистые волосы и у взрослых.
Лоб у негров прямой, с хорошо выраженными лобными буграми, а у индонезийских австралоидов средненаклонный или, как у австралийцев, даже довольно покатый, причём и надглазничные дуги у них обычно сильно развиты, а у негров - слабо. По форме лба негры, по-видимому, уклонились дальше от первоначального типа, чем океанийские австралоиды. В обратном смысле может быть истолкована разница в форме носа, который у африканских негров, как правило, уплощённый, в то время как у восточных негроидов он характеризуется выпуклой или прямой спинкой, у некоторых же меланезийцев - вогнутой.
Как видим, расовые различия между негроидами и австралоидами касаются преимущественно волосяного покрова, формы лба, надглазничных дуг и спинки носа. Перед подавляющим общим сходством эти различия, в конце концов, не так уже велики. Они объясняются, по-видимому, несколько иными путями развития обеих негроидно-австралоидных малых рас в заметно отличающихся и удалённых друг от друга областях земного шара.
Очевидно, надо предположить, что где-то в южной Азии, в Индостане или Индокитае, а может быть и западнее, в начале верхнего палеолита существовала исходная группа австралоидно-негроидного характера, впоследствии разделившаяся на западную и восточную ветви, которые позже утеряли территориальный контакт между собой.
Если допустить существование в южной Азии подобной предковой экваториальной расы в отдалённом прошлом, около 50 тыс. лет назад или даже раньше, то тогда легче представить себе и расселение дифференцировавшихся расовых групп преимущественно в юго-восточном, или океанийском, направлении в западном, а позже и в юго-западном, или африканском, направлениях. Распространение шло параллельно с преобразованиями сложившихся расовых типов и дифференциацией новых. При этом у негроидов узкая волнистость волос на голове превратилась в спиральную завитость; произошло ослабление третичного волосяного покрова; лоб стал прямым, надглазничные дуги уменьшились; спинка носа в некоторых группах приобрела прямизну. Это был, надо полагать, очень сложный процесс, и мы его ещё не можем очертить детально за недостатком антропологических и археологических материалов. Сейчас необходимо перенести наше внимание исключительно на западных (африканских) и восточных (океанийских) негроидо-австралоидов. Ещё раз повторяем, что сходство в их расовых признаках свидетельствует о родстве и общем происхождении.
В пользу самостоятельного, автохтонного развития негроидов в Африке обычно приводится два довода.
Первый заключается в наличии большого разрыва между территориями, которые заняты западными и восточными негроидами.
Однако значение этого факта в известной мере ослабляется существованием эфиопской расы в восточной Африке, а также дравидов и веддов (рис. 50) в Индии, территориально менее сильно отдалённых от эфиопов. Территориальный разрыв между западными и восточными негроидами не может служить непререкаемым свидетельством в пользу генетического разрыва между этими двумя близкими группировками темнокожих рас.

Рис. 50. Ведды - мужчина (слева) и женщина (справа). (Цейлоно-зондская группа типов австралоидной малой расы.)
Второй довод в пользу мнения об автохтонном появлении негров в Африке основывают на палеантропологических источниках, приписывая чрезмерную древность и примитивность, вместе с негроидными чертами, костным остаткам ископаемых людей на территории материка.
Костные остатки древних негров были обнаружены не так давно. В 1927 г. в глубине пустыни Сахары, близ военного поста Асселяр* в отложениях плейстоценовой древности был найден почти полный окаменелый скелет негроидного типа (рис. 51). Однако древность его относится к поздней (мадленской) эпохе верхнего палеолита. Судя по скелету, длина тела его обладателя была не менее 170 см; вместимость мозговой коробки около 1500 куб. см; черепной указатель - 70,9 (долихокрания).
* (В 400 км к северо-востоку от г. Тимбукту и около 200 км к юго-востоку от г. Эль-Мабрук.)
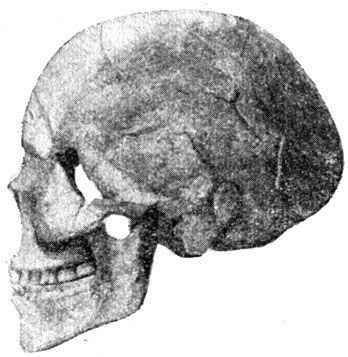
Рис. 51. Череп негроидного типа из Асселяра, Сахара (1927)
Другая интересная находка негроидного черепа была сделана в 1939 г. близ Найваши (восточная Африка). Всё же и он не обладает такой древностью, чтобы считаться свидетельством особой стадии развития негрской расы: по расовым особенностям он сходен с современными негрскими черепами.
Больше надежд защитники полицентризма обычно возлагают на два более примитивные и, может быть, более древние, черепа из Брокен-Хилла и Ньярасы (Эяси). Что касается первого черепа (рис. 52), найденного в Северной Родезии (южная Африка) в 1921 г., то он совершенно непохож на негрские черепа и должен быть снят со счетов. К тому же и геологическая датировка его совершенно неопределённая. Череп этот сходен с неандертальскими: у него нависающий мощный надглазничный валик, очень покатый лоб, сильный наружный рельеф. Большое затылочное отверстие располагается почти горизонтально, ближе к середине основания черепа, как и у современного человека. Объём мозговой коробки - около 1200 см. Рост родезийца, судя по приписываемой ему большой бердовой кости, был около 180 см. Находка в Брокен-Хилле может обозначать, что была какая-то более древняя группа гоминид, которые проникали в Африку, возможно, из Азии, далее заметно не эволюционировали и вымерли, не оставив потомков.
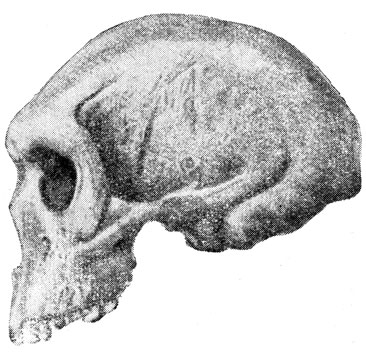
Рис. 52. Череп человека из Брокен-Хилла, Северная Родезия (1921)
Не обладает негроидными особенностями и череп, найденный близ озера Ньяраса (восточная Африка) в 1935 г.
Таким образом, в Африке неандертальских черепов с негроидными чертами не обнаружено.
В пользу проникновения протонегроидов в Африку из Азии говорят другие находки, а именно восточноафриканские негроидные скелеты верхнепалеолитического возраста из Олдовая и Гэмбля (восточная Африка). Эти древние негроиды, отличающиеся своим сравнительно высоким лицом, ближе всего, по-видимому, стоят к эфиопской группе антропологических типов. Местоположение указанных находок свидетельствует скорее о миграции южноазиатской древней негроидной расы на запад и юго-запад" из Аравии в Сомали.
Ещё один вероятный след такой миграции южноазиатских негроидов на запад остался в виде десятков скелетов, найденных в пещерах у Шукбы и на горе Кармел. Эти скелеты принадлежат ископаемым людям верхнего палеолита (или мезолита). Скелет из Асселяра, судя по чертам строения, свидетельствует в пользу родства между восточными и западными негроидо-австралоидами. В населении зоны, идущей из северо-восточной Африки через Переднюю Азию, Индию и Индонезию к Австралии, происходит, начиная с древних времён и до наших дней, своеобразное переплетение признаков негроидов и австралоидов, тянутся подчас малозаметные, но определённые нити родства между африканской и океанийской малыми расами. Этот факт тоже свидетельствует о родстве между негроидами и австралоидами.
Одной из характерных особенностей экваториальной расы является наличие в её составе пигмеев - карликовых антропологических типов, которые неизвестны в европеоидной и монголоидной больших расах. Африканских пигмеев называют негриллями, а океанийских-негритосами (оба термина уменьшительные от слова "негр").
Проблема возникновения пигмеев представляет большой интерес для расоведения, изучающего процесс расогенеза, и для учения об антропогенезе.
Вокруг проблемы пигмеев между прогрессивными и реакционными антропологами давно идёт спор. В пигмеях реакционные зарубежные учёные хотят видеть древнейшую, "низшую из низших", чуть ли не обезьяноподобную расу, которая обречена на полное вымирание.
В советской антропологии эта точка зрения подверглась специальному анализу, показавшему её полную научную несостоятельность и реакционную сущность. Западная и восточная пигмейские группы антропологических типов обладают большой жизненной стойкостью, не обнаруживают признаков деградации, биологически совершенно равноценны любой другой группе антропологических типов, способны к полному и быстрому культурному развитию.
Одновременно советская антропология подвергла основательной критике гипотезу зарубежных учёных о пигмеях, как предках всего человечества. Действительно, древнейшие люди обладали более высоким ростом, чем пигмеи (синантроп 152-163 см, питекантроп около 170 см). Неандертальцы точно так же выше пигмеев (155-160 см). Таким образом, пигмеи не являются реликтом первой или второй стадии эволюции человека. Из этого следует, что карликовость в современном человечестве есть явление вторичное, притом частное и местное, так как встречается лишь в одной, а не во всех больших расах человечества.
Другими словами, пигмеи, как и высокорослые люди, произошли от среднерослых представителей своих рас или групп антропологических типов.
Начнём с негриллей, которых объединяют в центральноафриканскую, или пигмейскую, группу антропологических типов*.
* (Негрилли живут в центральной зоне экваториальной Африки, в глухих тропических лесах. Восточные негрилли (бамбути) обитают в области Итури, центральные (батуа)- в Бельгийском Конго, а западные (бабинга)- во Французской экваториальной Африке, включая Камерун.)
Средний рост негриллей не превышает 150 см. В некоторых племенах рост мужчин падает до 140 см, а женщин до 130-125 см. Борода встречается не у всех; третичный волосяной покров на теле у представителей одних племён слабо развит, у других - умеренный. Голова относительно крупная, мезокефальная; лицо низкое, но с круглыми, высокими глазницами. Глаза карие; губы средней толщины или даже тонкие; нос уплощённый, переносье низкое или среднее. Туловище довольно длинное по сравнению с более короткими ногами; скелет руки тонкий, грацильный. В общем негрилли очень похожи на своих соседей негров, так как кожа у них большей частью темнопигментированная, волосы на голове спирально закрученные, нос очень широкий, лоб выпуклый (рис. 53).

Рис. 53. Африканский пигмей неггрилль бабинга из Санги, Французское Конго. (Центральноафриканская группа типов негроидной малой расы.)
Рассмотрим теперь антропологические особенности негритосов, которые обитают на острове Новая Гвинея, на Ново-Гебридских островах и ближайших к ним.
Одна группа негритосов Новой Гвинеи более похожа на меланезийцев, например на обитателей островов Новая Каледония.
Рост их достигает 150-152 см. Другая группа негритосов Новой Гвинеи больше похожа на папуасов, но у них более широкий нос; им свойственна также мезокефалия, в то время как папуасы отличаются долихокефалией. Рост этих негритосов ещё меньше и опускается у мужчин до 144 см; их можно рассматривать как местный вариант папуасской группы.
С негритосами Новой Гвинеи очень сходны ещё несколько океанийских антропологических типов; таковы андаманцы с Андаманских островов, аэта с Филиппинского острова Люцон и семанги с полуострова Малакка (рис. 54). Все эти пигмеи объединяются некоторыми учеными в одну группу антропологических типов. Однако происхождение указанных типов различно, и они занимают очень разобщённые ареалы. Поэтому вряд ли всех этих пигмеев можно соединить в одну группу.

Рис. 54. Азиатские пигмеи негритосы. Аэта с Филиппинских островов (слева) и семанг, мальчик, с полуострова Малакка (справа). (Андаманская, или негритосская, группа типов австралоидной малой расы.)
В отношении новогвинейских пигмеев достаточно обосновано мнение об их происхождении от меланезийской группы антропологических типов. Это следует хотя бы из того факта, что пигмеи тапиро Новой Гвинеи незаметными переходами связаны с низкорослым племенем аруп (рост 160 см), обитающим на севере острова. По-видимому, и другие низкорослые племена тоже происходят от соседних групп либо представляют собой изменённые, сохранившиеся в лесах и горных областях остатки тех групп антропологических типов, которые некогда переселились из Индокитая или южного Китая через ближайшие острова Малайского архипелага на юго-восток, где постепенно приостановили свой путь и прочно осели.
Негрилли и негритосы занимают ареалы, удалённые один от другого на 10-15 тыс. км. Как понять такую разорванность ареалов, если допустить, что они имели общее происхождение от гипотетической предковой карликовой расы, занимавшей какой-то промежуточный ареал в южной Азии, откуда протопигмеоиды могли бы расселиться на юго-восток и на юго-запад? Подобное предположение, в частности, неверно и потому, что в южной Азии ископаемые костные остатки карликовых людей не обнаружены.
Из западных негроидов к пигмеям по росту довольно близки бушмены (рис. 55), входящие в южноафриканскую, или бушменскую, группу типов. Эти очень низкорослые люди (152-155 см) почти полностью истреблены англичанами-колонизаторами и лишь в ничтожном количестве ещё сохранились в самых скрытых участках южноафриканской пустынно-степной области Калахари и в пустыне Намиб.
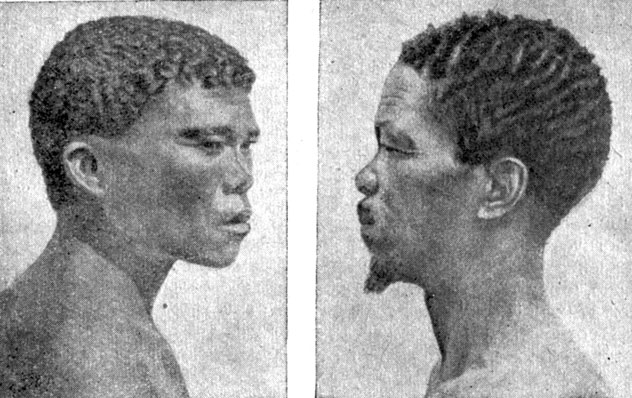
Рис. 55. Молодой (слева) и взрослый (справа) бушмены из пустыни Калахари. (Южноафриканская группа типов негроидной малой расы.)
Кроме низкого роста, у бушменов отмечаются и другие черты близости к пигмеям: короткие (относительно туловища) ноги, довольно крупная голова, плоское и очень низкое лицо, прямой низкий лоб, слаборазвитые надглазничные дуги, выступающие скулы, низкое переносье, широкие крылья носа, слаборазвитый подбородочный выступ.
Из других характерных особенностей бушменов отметим следующие: желтоватый цвет кожи (у женщин более светлый); морщинистость кожи лица; волосы на голове чёрные и спирально закручены сильнее, чем у негров; третичный волосяной покров на теле и лице почти отсутствует; глаза карие, складки нижнего и верхнего век сильно развиты, но эпикантус, как правило, отсутствует; губы утолщённые, верхняя губа выступает вперёд (прохейлична)*.
* (От греч. про - вперёд, хейле - губа (мкож. хейлиа).)
По цвету кожи, складке верхнего века и некоторой уплощённости лица бушмены напоминают монголоидов, с которыми, однако, у них нет особенно близкого родства. Складка века бушменов по своему строению отлична от монголоидной. Сходство с последней чисто внешнее и, возможно, вызвано влиянием природной среды пустыни.
Большинство расовых особенностей роднит бушменов с суданской, или собственно негрской, группой типов. Бушмены, очевидно, являются одним из её слабо пигментированных вариантов. Не противоречит мнению о негроидной природе бушменов и сильное развитие жировых скоплений в области ягодиц, так как подобное явление наблюдается и у некоторых других антропологических типов Африки, например у племён полуострова Сомали. Наибольших размеров эти жировые отложения достигают у соседей бушменов - готтентотов.
Антропологические данные о родственных связях бушменов с неграми можно дополнить археологическими. Так, древние рисованные, а также некоторые резные изображения животных и людей, встречающиеся на скалах почти по всей южной, а местами и в восточной Африке, очень сходны с бушменскими. Это говорит о широком распространении бушменов по Африке в прошлом. Возможно, что они являются на материке одной из древнейших групп населения.
О родстве между бушменами и негроидо-австралоидами свидетельствуют и палеантропологические находки нескольких черепов, вроде найденного в Кэп Флетс (южная Африка).
Таким образом, бушменская группа типов, несомненно, принадлежит к африканской негроидной малой расе. Если её положение и кажется несколько обособленным, то это лишь свидетельствует о том, что филогенетическое родство не всегда выявляется по одним только внешним признакам.
Это в особенности следует иметь в виду при рассмотрении групп антропологических типов австралоидной расы, в которой австралийская группа типов является наиболее характерной. Коренные жители Австралии окончательно сформировались в результате очень длительной географической изоляции на сравнительно небольшом материке, бедном природой и удалённом от континента Азии.
Совокупность расовых черт австралийцев говорит о наибольшей близости к типу негроидов, хотя волнистые волосы, сильно развитый третичный волосяной покров на лице и теле и некоторые другие особенности заставляют подумать о каком-то отдалённом родстве австралийцев и с европеоидами.
Австралийцы (рис. 56) не стоят изолированно среди других групп австралоидов. Более близкое сходство они имеют с некоторыми меланезийцами, например с новокаледонцами, у которых заметно развит волосяной покров на теле, а волосы по форме ближе к волнистым. Однако родственные связи австралийцев можно проследить и дальше на северо-запад, вплоть до Индии и Цейлона, где локализуются во многом сходные с ними антропологические типы веддов и дравидов. Интересно, что последние многими чертами сближаются с эфиопской группой типов. Таким образом, генетическая связь, вероятно, очень древняя, тянется от европеоидов не только к африканским негроидам, но и через Индию к океанийским австралоидам.
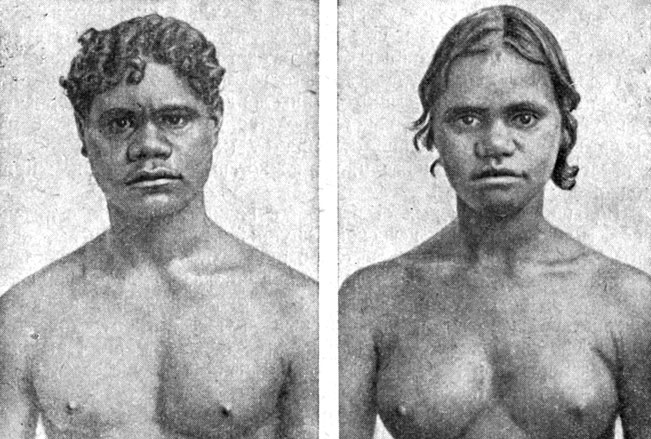
Рис. 56. Австралийцы: юноша (слева) и девушка (справа) из Квинсленда. (Австралийская группа типов австралоидной малой расы.)
Возможно, что в юго-восточной Азии в самые последние эпохи каменного века формировались группы типов, близкие к типу веддов Цейлона. Это отчасти подтверждается находками костных остатков в Индокитае, а также в Индонезии.
Прародиной группы австралийских типов, как и меланезийцев, был, по-видимому, юго-восточный угол материка Азии. Из Индокитая предки австралийцев, вероятно, в эпоху верхнего палеолита, переселились в Австралию через острова Молуккские, Церам и Новую Гвинею, или, южнее, через острова Большие Зондские, Филиппинские, Целебес, Тимор.
Распространяясь по более плодородным прибрежным районам, австралийцы, очевидно, встретили в восточной части Австралии более древних пришельцев - представителей тасманийского антропологического типа, другая часть которых до того уже успела перебраться через Бассов пролив на остров Тасмания.
В некоторой степени это предположение подтверждается находками ископаемых форм. В юго-восточном углу Австралии обнаружены три неполных черепа: в 1884 г. в Талгае, в 1925 г. в Когуне и в 1940 г. в Кэйлоре.
Рис. 59. Метисы европейцев с тасманийками.
Череп из Талгая (рис. 57) принадлежит юноше 14-16 лет, а череп из Когуны - взрослому индивидууму. Их геологическая датировка - примерно конец ледниковой эпохи. Оба сходны с австралийскими черепами не только по форме и строению, но и по ёмкости мозговой коробки, которая у современных австралийцев мужчин в среднем равна 1300 куб. см.
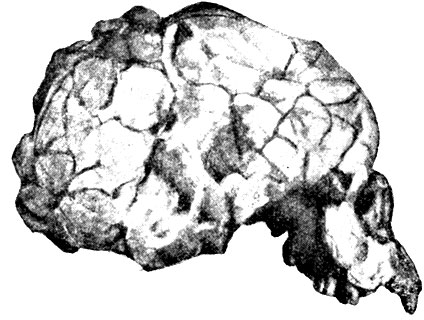
Рис. 57. Череп из Талгая, Австралия (1925)
В Кэйлоре был найден более полный череп, который принадлежит взрослому индивидууму. Его геологическая древность близка к эпохе последнего оледенения. По форме и большой вместимости мозговой коробки (1590 куб. см) кэйлорский череп заметно отличается от предыдущих и очень напоминает вадьякский. В 1889 г. на острове Ява у Вадьяка голландским учёным Дюбуа (который открыл позже и питекантропа) были найдены остатки двух черепов. Вместимость мозговой коробки более полно сохранившегося вадьякского черепа составляет 1650 куб. см.
Черепа из Вадьяка подтверждают давность появления океанийской расы и на материке Австралии: очевидно это были предки тасманийцев (до юго-восточной его оконечности австралийцы добрались позже, чем до других прибрежных районов).
Тасманийская группа антропологических типов представляет интерес не меньший, чем австралийцы. К сожалению, в настоящее время ни одного тасманийца уже не осталось в живых. Сто лет назад английские войска вместе с колонистами произвели поголовное истребление населения Тасмании, которое тогда определялось примерно в 5 тыс. человек. Всего же в момент открытия этого острова на нём было около 15 тыс. населения. Случайно оставшиеся в живых 150 тасманийцев были увезены, англичанами на маленький островок, где они и вымерли все до одного. Последней скончалась тасманийка Труганина в 1876 г. (рис. 58). Несколько тасманийцев были завезены на южный берег Австралии, где они смешались с австралийцами, а также с европейцами (рис. 59).
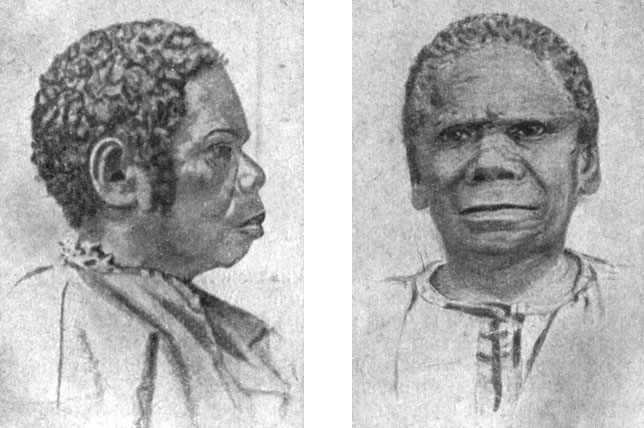
Рис. 58. Тасманийки: слева - Труганина и справа - Патти-о-Коонеана. (Меланезийская группа типов австралоидной малой расы.)

Рис. 59. Метисы европейцев с тасманийками
При использовании материалов сайта активная ссылка обязательна:
http://anfiz.ru/ 'Анатомия и физиология человека'