Глава XIII. Физиология органов чувств (анализаторы)
Рецепторы и их характеристика
Центральная нервная система постоянно получает информацию о внешнем мире и внутреннем состоянии организма от рецепторов, которые представляют собой чувствительные нервные окончания, специализированные к воздействию различных раздражений.
Рецепторы отличаются друг от друга по своей структуре и выполняемым функциям. В зависимости от характера воздействующего раздражителя рецепторы делят на экстерорецепторы, интерорецепторы и проприорецепторы.
Экстерорецепторы воспринимают раздражения из внешней среды. К ним относят зрительные рецепторы глаза (фоторецепторы), слуховые рецепторы (фонорецепторы), обонятельные рецепторы слизистой оболочки носа, вкусовые рецепторы, расположенные в слизистой оболочке языка, температурные, болевые и тактильные рецепторы кожи и слизистых оболочек.
Интерорецепторы расположены во внутренних органах, в стенке сосудов. Они возбуждаются изменениями состояния и деятельности органов и внутренней среды организма.
Проприорецепторы локализованы в мышцах, сухожилиях, связках, суставных поверхностях костей. Они возбуждаются в результате растяжения мышц и изменения положения конечностей и других частей тела по отношению друг к другу и в пространстве.
Все рецепторы можно разделить на две большие группы: дистантные и контактные. Дистантные рецепторы способны воспринимать раздражения от предметов, находящихся на значительном расстоянии от организма (зрительные, слуховые, обонятельные рецепторы). Контактные рецепторы воспринимают раздражения только от предметов, которые непосредственно к ним приложены, т. е. находятся в близком соприкосновении с рецепторным аппаратом. К ним относят тактильные, температурные, вкусовые рецепторы.
Рецепторы трансформируют энергию раздражения в энергию нервного импульса. Причиной возникновения возбуждения в рецепторе является деполяризация его мембраны в результате воздействия раздражителя. Эту деполяризацию называют рецепторным, или регенераторным, потенциалом. Образование рецепторного потенциала связано с повышением проницаемости мембраны для ионов натрия. Когда рецепторный потенциал достигает определенной - критической - величины, он вызывает разряд афферентных импульсов в нервном волокне, связанном с рецептором.
Одним из свойств рецепторов является адаптация, т. е. приспособление к силе раздражителя. Способностью к адаптации обладают в большей или в меньшей степени почти все рецепторы. Исключение составляют проприорецепторы.
Явление адаптации заключается в том, что происходит снижение чувствительности рецепторов к постоянно действующему раздражителю. Внешне адаптация проявляется в привыкании к постоянно действующему раздражителю - запаху, шуму, давлению одежды и т. д.
Как только постоянное действие раздражителя заканчивается, возникшая под его влиянием адаптация постепенно исчезает. Чувствительность рецепторов при этом повышается.
Орган зрения
Органом зрения является глаз. Расположено глазное яблоко в полости глазницы. Его стенку образуют три оболочки. Переднюю часть наружной оболочки глаза называют роговицей, которая в дальнейшем переходит в склеру, или белковую оболочку. Следующей оболочкой глаза является сосудистая. Внутренняя оболочка - сетчатка, на которой расположены фоторецепторы.
В состав глаза входят рецепторный аппарат, находящийся в сетчатке, и оптическая система. Значение последней заключается в том, что она собирает световые лучи и обеспечивает четкое действительное изображение предметов на сетчатке, но в уменьшенном и обратном виде.
Оптическая система глаза представлена передней и задней поверхностью роговой оболочки, хрусталиком и стекловидным телом. Поступающие в глаз световые лучи проходят через оптическую систему глаза и попадают на сетчатку. Ход лучей зависит от показателей преломления и радиуса кривизны поверхности роговой оболочки, хрусталика и стекловидного тела. Преломляющую силу оптической системы глаза выражают в диоптриях. Одна диоптрия (дптр) - преломляющая сила линзы, имеющей фокусное расстояние 100 см. При увеличении преломляющей силы фокусное расстояние уменьшается. Преломляющая сила оптической системы глаза при рассматривании далеких предметов составляет около 59 дптр, при рассматривании близких предметов - 70,5 дптр.
Для ясного видения предмета необходимо, чтобы лучи от всех его точек падали на сетчатку. Человек с нормальным зрением может хорошо рассмотреть как близко, так и далеко расположенные предметы. Приспособление газа ясному видению разноудаленных предметов называют аккомодацией. Аккомодация осуществляется путем изменения кривизны хрусталика, что приводит к сдвигам в его преломляющей способности. При рассматривании близких предметов хрусталик делается более выпуклым, благодарящему расходящиеся лучи от предмета сходятся на сетчатке глаза.
Механизм аккомодации глаза связан с сокращением ресничных мышц, которые изменяют выпуклость хрусталика.
Хрусталик заключен в капсулу, переходящую по краям в волокна цинновой связки, прикрепленной к ресничному телу (рис. 91). Цинновые связки всегда натянуты, и их натяжение передается капсуле, сжимающей и уплотняющей хрусталик. В ресничном теле находятся гладкие мышечные волокна. При их сокращении наступает ослабление тяги цинновых связок, а значит, уменьшение давления на хрусталик, который вследствие своей эластичности принимает более выпуклую форму.
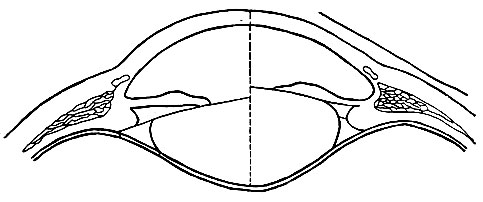
Рис. 91. Механизм аккомодации (по Гельмгольцу). Левая половина глаза - хрусталик при рассматривании далекого предмета, правая - близкого предмета. Видна большая выпуклость хрусталика справа
Существуют две главные аномалии преломления лучей в глазу - дальнозоркость и близорукость. Они связаны, как правило, не с недостаточностью преломляющих сред, а с ненормальной длиной глазного яблока (рис. 92).
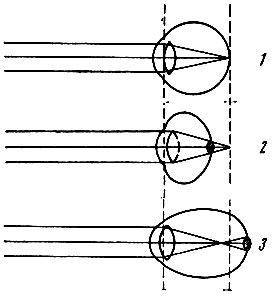
Рис. 92. Схема рефракции в норме (1), при дальнозоркости (2) и при близорукости (3)
Дальнозоркость. В дальнозорком глазу продольная ось глаза короткая, поэтому параллельные лучи, идущие от далеких предметов, собираются сзади сетчатки. На сетчатке же получается круг светорассеяния, т. е. неясное, расплывчатое изображение предмета. Этот недостаток рефракции может быть исправлен путем применения двояковыпуклых стекол, усиливающих преломление лучей.
Близорукость. Если продольная ось глаза слишком длинная, то главный фокус будет находиться не на сетчатке, а перед ней, в стекловидном теле. В этом случае параллельные лучи сходятся в одну точку не на сетчатке, а где-то ближе ее. На сетчатке вместо точки возникает круг светорассеяния. Чтобы ясно видеть вдаль, близорукий человек должен поместить перед глазами вогнутые стекла, которые уменьшают преломляющую силу хрусталика и тем самым отодвигают изображение на сетчатку.
Сетчатка и ее строение. Сетчатка имеет сложную многослойную структуру. Здесь расположены два вида различных по своему функциональному значению фоторецепторов - палочки и колбочки - и несколько видов нервных клеток с их многочисленными отростками.
Каждая палочка или колбочка состоит из наружного членика, чувствительного к действию света, содержащего зрительный пигмент, и внутреннего сегмента, в котором находятся ядро и митохондрии, обеспечивающие энергетические процессы в фоторецепторной клетке. В палочках содержится пигмент родопсин, или зрительный пурпур, в колбочках - пигмент йодопсин.
При действии света в палочках и колбочках осуществляются физические и химические процессы. В частности, под влиянием света родопсин разрушается. При затемнении глаз происходит восстановление зрительного пурпура. Для этого необходим витамин А. Если же в организме витамин А отсутствует, то образование родопсина резко нарушается, что приводит к развитию так называемой куриной слепоты, т. е. неспособности видеть при слабом свете или в темноте. Йодопсин также подвергается разрушению под влиянием света и образуется в темноте. Установлено, что распад йодопсина в отличие от зрительного пурпура совершается в 4 раза медленнее. У человека в сетчатке имеется около 6-7 млн. колбочек и 110-125 млн. палочек. Палочки и колбочки распределены в сетчатке неравномерно. Центральная ямка сетчатки содержит только колбочки (до 140 тыс. на 1 мм2). По направлению к периферии сетчатки количество колбочек уменьшается, а палочек соответственно возрастает. Периферическая часть сетчатки содержит исключительно палочки. Участок сетчатки глаза, где сосредоточены только колбочки, получил название желтого пятна. Место выхода зрительного нерва из глазного яблока - сосок (диск) зрительного нерва - совсем не содержит фоторецепторов и нечувствителен к свету. Это так называемое слепое пятно. В существовании слепого пятна можно убедиться с помощью опыта Мариотта. Если, закрыв левый глаз, правым фиксировать крестик, изображенный на рис. 93, то при определенном расстоянии рисунка от глаза (приблизительно около 25 см) круг исчезает, так как его изображение падает на слепое пятно.
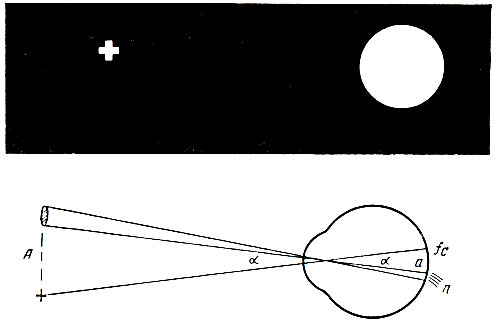
Рис. 93. Рисунок для проведения опыта Мариотта и схема хода лучей в этом опыте. Овал на рисунке представляет собой светлый кружок. Если правый глаз фиксирован на крестике, то изображение кружка падает на слепое пятно, лежащее у места вхождения зрительного нерва (n), а изображение крестика - на центральную ямку (fc)
Колбочки осуществляют дневное зрение и воспринимают цвета. Палочки обеспечивают сумеречное, ночное зрение.
Свет, попадая на сетчатку глаза, вызывает изменение зрительного пигмента в палочках и колбочках. Один из образовавшихся промежуточных продуктов превращения родопсина приводит фоторецепторы сетчатки глаза в возбуждение. Возникшие нервные импульсы передаются на нервные клетки сетчатки глаза, в которых осуществляется их сложная обработка. Переработанные нервные импульсы по волокнам зрительного нерва поступают в затылочную область - мозговой конец анализатора. Полагают, что по волокнам зрительного нерва передаются сигналы не о состоянии каждого отдельного рецептора, а об определенных параметрах того или иного изображения, об элементах зрительных образов.
Мозговой конец зрительного анализатора представлен 17, 18 и 19-м полями Бродмана. В нейронах этих участков коры головного мозга осуществляется переработка поступившей информации, в результате возникают зрительные ощущения - формы и размера предмета, его цвета, расположения в пространстве и т. д.
Адаптация. От освещенности предмета зависит чувствительность глаза к восприятию света. Так если человек перейдет из темного помещения в светлое, то в первое время наступает ослепление. Постепенно глаз адаптируется к свету за счет понижения чувствительности фоторецепторов сетчатки глаза. 15то явление получило название световой адаптации. При переходе из светлого помещения в темное человек сначала ничего не видит. Через некоторое время чувствительность фоторецепторов сетчатки повышается, появляются контуры предметов, затем начинают различаться их детали, т. е. проявляется темновая адаптация.
Бинокулярное зрение. Рассматривание предметов обоими глазами называют бинокулярным зрением. Когда человек смотрит на какой-либо предмет обоими глазами, то у него не получается восприятия двух одинаковых предметов. Это связано с тем, что изображения от всех предметов при бинокулярном зрении падают на соответственные, или идентичные, участки сетчатки, в результате чего в представлении человека эти два изображения сливаются в одно. Если слегка надавить сбоку на один глаз, то у человека начинает "двоиться" в глазах, так как в этом случае изображения от предмета падают на неидентичные участки сетчатки.
Бинокулярное зрение имеет большое значение в определении расстояния до предмета, его формы. Оценка величины предмета связана с размером его изображения на сетчатке и расстоянием предмета от глаза.
Цветовое зрение. Человек обладает способностью различать большое количество цветов. Наиболее признанной теорией, объясняющей механизм цветового зрения, является трехкомпонентная теория Ломоносова - Гельмгольца. В свое время М. В. Ломоносов (1756) высказал предположение о наличии в сетчатке трех типов элементов, воспринимающих при основных цвета. Эти положения М. В. Ломоносова были в дальнейшем разработаны Юнгом, а затем Гельмгольцем. Согласно трехкомпонентной теории, в сетчатке имеются три типа фоточувствительных колбочек, воспринимающих красный, зеленый и сине-фиолетовый цвета. Разложение светочувствительных веществ, находящихся в колбочках, вызывают раздражение нервных окончаний. Возбуждение, дошедшее до коры головного мозга, суммируемся, и возникает ощущение одного однородного цвета. Одинаковое и одновременное раздражение трех типов цветовоспринимающих элементов сетчатки даст ощущение белого цвета.
Возникновение ощущения того или иного цвета связано с раздражением глаза лучами видимой части спектра различной длины. Раздражение глаза электромагнитными волнами длиной 620-760 нм (6200-7600 А) вызывает ощущение красного цвета, 510-550 нм (5100-5500 А) - зеленого, 390-450 нм (3900-4500 А) - фиолетового цвета.
В настоящее время, используя микроэлектродную технику и регистрируя биотоки от отдельных нервных волокон, отходящих от колбочек, шведские физиологи обнаружили в сетчатке четыре типа колбочек, реагирующих на электромагнитные волны красной, зеленой и синей частей спектра. Четвертый тип колбочек возбуждается при изменении яркости света.
Орган слуха
Значение органа слуха заключается в восприятии звуковых колебаний. Звук представляет собой колебания частиц воздуха различной частоты, периодичности и амплитуды. Орган слуха включает наружное, среднее и внутреннее ухо.
Строение наружного уха. В состав наружного уха входят ушная раковина, наружный слуховой проход, барабанная перепонка. За счет ушной раковины улавливаются звуковые колебания. Наружный слуховой проход служит для проведения звуковых колебаний к барабанной перепонке. Колебания, возникающие в источнике звука, поступают к ближайшему уху на несколько долей миллисекунды раньше, чем к другому. Указанное обстоятельство определяет способность человека или животного улавливать направление звука.
Наружное ухо от среднего отделяется барабанной перепонкой. Она имеет форму вдавленной внутрь среднего уха воронки. С внутренней стороны барабанная перепонка соединена с рукояткой молоточка. Колебания барабанной перепонки происходят тогда, когда на нее падают звуковые колебания, улавливаемые наружным ухом. Натяжение барабанной перепонки в различных частях неодинаковое. Это приводит к тому, что она не имеет своего собственного периода колебаний и колеблется при всяком звуке соответственно длине его волны.
Строение среднего уха. В состав среднего уха входит система слуховых косточек - молоточек, наковальня, стремечко (рис. 94). Одна из этих косточек - молоточек - вплетена своей рукояткой в барабанную перепонку, другая сторона молоточка сочленена с наковальней. Наковальня соединена со стремечком, которое прилегает к мембране овального окна внутренней стенки среднего уха.
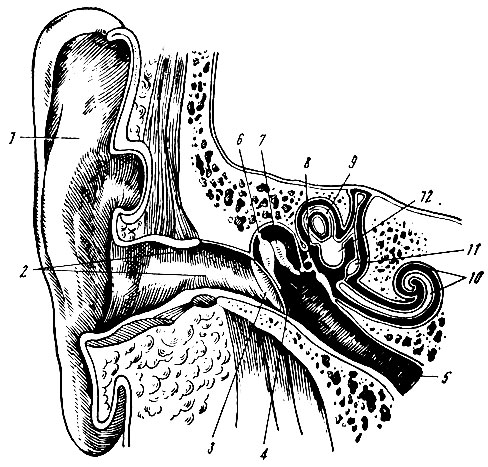
Рис. 94. Схема строения уха. 1 - ушная раковина; 2 - наружный слуховой проход; 3 - барабанная перепонка; 4 - полость среднего уха; 5 - слуховая труба; 6 - молоточек; 7 - наковальня; 8 - стремячко; 9 - полукружный канал; 10 - улитка; 11 - мешочек; 12 - маточка
Значение слуховых косточек состоит в том, что они участвуют в передаче колебаний барабанной перепонки, вызванных звуковыми волнами, к овальному окну, а затем на эндолимфу улитки внутреннего уха.
Овальное окно расположено на стенке, отделяющей среднее ухо от внутреннего. Там же имеется круглое окно. Колебания эндолимфы улитки, начавшиеся у овального окна, распространяются по ходам улитки, не затухая, до круглого окна.
При помощи особого канала - евстахиевой (слуховой) трубы, полость среднего уха соединяется с носоглоткой. Это обеспечивает поддержание в среднем ухе давления, равного атмосферному. Если давление в полости среднего уха отличается от атмосферного, то это приводит к понижению остроты слуха, так как нарушаются нормальные колебания барабанной перепонки.
Строение внутреннего уха. В состав внутреннего уха (лабиринта) входят преддверие, полукружные каналы и улитка, в которой расположены особые рецепторы, реагирующие на звуковые волны. Преддверие и полукружные каналы к органу слуха не относятся. Они представляют собой вестибулярный аппарат, который участвует в регуляции положения тела в пространстве и сохранении равновесия.
Улитка - это костный постепенно расширяющийся спиральный канал, образующий два с половиной витка. Костный канал на всем своем протяжении разделен двумя перепонками: более тонкой, называемой вестибулярной мембраной, или мембраной Рейснера, и более плотной и упругой, получившей название основной мембраны (рис. 95). Эти мембраны на вершине улитки соединяются. В этом месте имеется отверстие. Костный канал улитки за счет вестибулярной и основной мембран разделяется на три узких хода: верхний, средний и нижний (см. рис. 96).

Рис. 95. Поперечный разрез завитка улитки (по Расмуссену). 1 - лестница преддверия; 2 - барабанная лестница; 3 - вестибулярная мембрана; 4 - основная мембрана; 5 - перепоночный канал улитки; 6 - покровная мембрана; 7 - кортиев орган; 8 - секреторный эпителий; 9 - спиральная связка; 10 - спиральный ганглий; 11 - наружные волосковые клетки; 12 - внутренние волосковые клетки; 13 - нервные волокна, подходящие к волосковым клеткам
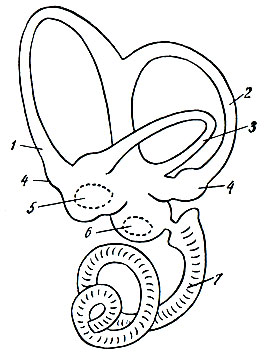
Рис. 96. Схема левого лабиринта млекопитающего. 1, 2, 3 - полукружные каналы; 4 - ампула полукружного канала; 5, 6 - преддверие; 5 - утрикулюс; 6 - саккулюс (пунктирной линией обозначены слуховые пятна); 7 - улитка
От овального отверстия начинается верхний ход, он продолжается до вершины улитки. Нижний ход улитки берет начало в области круглого окна и заканчивается на вершине улитки. Здесь через имеющееся отверстие происходит сообщение обоих ходов друг с другом. В результате верхний и нижний ходы являются как бы единым каналом, идущим от овального до круглого окна. В верхнем и нижнем ходах улитки имеются перилимфа, состав, которой сходен с составом спинномозговой жидкости. Мембраны овального и круглого окон отделяют перилимфу ходов от воздушной полости среднего уха.
Средний ход находится между верхним и нижним. Он образован вестибулярной и основной мембранами. Его полость заполнена эндолимфой и не сообщается с другими ходами улитки.
На основной мембране среднего хода улитки имеется звуковоспринимающий аппарат - кортиев орган. В его состав входят рецепторные волосковые клетки, которые превращают звуковые колебания в нервные импульсы, распространяющиеся по волокнам слухового нерва. Одна часть клетки располагается на основной мембране, вторая - в полости среднего хода улитки. На этом конце рецепторной клетки находятся волоски, которые омываются эндолимфой и располагаются в непосредственной близости от покровной мембраны. При проведении звуков через перилимфу и эндолимфу происходят колебания основной мембраны вместе с рецепторными клетками. При этом волоски рецепторных клеток контактируют с покровной мембраной и деформируются. Это приводит к возникновению возбуждения в рецепторных клетках.
Передача звуковых колебаний. Существует два вида передачи звуковых колебаний - воздушная и костная проводимость звука. При воздушной проводимости звука звуковые колебания усиливаются ушной раковиной и передаются по наружному слуховому проходу на барабанную перепонку. Она начинает колебаться с частотой, соответствующей частоте звука. Колебания барабанной перепонки передаются системе слуховых косточек: молотку, наковальне и стремечку. Звуковые колебания переключаются стремечком на мембрану овального окна и вызывают колебания перилимфы в верхнем и нижнем ходах улитки. В дальнейшем они доходят до круглого окна и приводят к смещению мембраны круглого окна наружу по направлению к полости среднего уха. Колебания перелимфы верхнего канала через вестибулярную мембрану передаются на эндолимфу среднего хода. Звуковые колебания, распространяющиеся по перилимфе и эндолимфе верхнего и среднего ходов, приводят в движение основную мембрану. Вместе с основной мембраной начинают колебаться волосковые клетки. Во время контакта этих клеток с покровной мембраной они возбуждаются, возникшие нервные импульсы по слуховому нерву и проводящим слуховым путям поступают в височную долю коры головного мозга. Нейроны височной доли коры головного мозга приходят в состояние возбуждения, и возникает ощущение звука.
При воздушной проводимости звука человек способен воспринимать звуки в очень широком диапазоне - от 16 до 20000 колебаний в 1 с.
Костная проводимость звука осуществляется через кости черепа. Если поставить ножку звучащего камертона на темя или сосцевидный отросток, то звук, будет слышен даже при закрытом слуховом проходе.
Это связано с тем, что звуковые колебания хорошо проводятся костями черепа, передаются сразу на перилимфу. Верхнего и нижнего ходов улитки внутреннего уха, а. затем - эндолимфу среднего хода. Происходит колебание основной мембраны с волосковыми клетками, в результате чего они возбуждаются и возникшие нервные импульсы в дальнейшем передаются к нейронам мозга.
Воздушная проводимость звука выражена лучше, чем костная. Если ножку звучащего камертона поставить на сосцевидный отросток и держать его до прекращения ощущения звука, а затем поднести этот же камертон к открытому слуховому проходу, то снова услышим звук.
Вестибулярный аппарат
Вестибулярный аппарату участвует регуляции положения и движения тела в поддержании равновесия. Вестибулярный аппарат состоит из преддверия и трех полукружных каналов внутреннего уха. Полукружные каналы - это узкие ходы правильной формы, которые располагаются в трех взаимно перпендикулярных плоскостях. Верхний, или передний, канал лежит во фронтальной, задний - в сагиттальной, а наружный - в горизонтальной плоскости. Один конец каждого канала колбообразно расширен и называется ампулой (рис. 96).
Преддверие вестибулярного аппарата состоит из двух частей - мешочка (саккулюс) и маточки (утрикулюс). Саккулюс располагается ближе к улитке внутреннего уха, утрикулюс - к полукружным каналам.
В обоих мешочках преддверия имеются возвышения, называемые пятнами. В них сосредоточены рецепторные клетки. На поверхности рецепторных клеток находится большое количество микроскопических кристаллических образование карбоната кальция. Эти образования называют отолитами. Они участвуют в возбуждении рецепторных клеток. При изменении положения головы, например при ее наклоне, изменяется давление отолитов на рецепторные клетки. что вызывает их возбуждение. В ампулах перепончатых полукружных каналов также имеются рецепторные клетки. Они сосредоточены в определенных местах, которые называют ампулярными кристами. Возбуждение рецепторных клеток происходит за счет перемещения эндолимфы каналов.
Повышение активности вестибулярного аппарата возникает только при ускоренном или замедленном вращательном либо прямолинейном движении, т. е. лишь под влиянием изменения скорости движения тела. Равномерное движение без ускорения или замедления не вызывает раздражения рецепторов вестибулярного аппарата. Раздражение отолитовых рецепторов или же рецепторов полукружных каналов происходит в зависимости от характера движения. Так, отолитовый аппарат возбуждается при ускоряющихся или замедляющихся прямолинейных движениях, тряске, качке, наклоне тела или головы в сторону, при которых наблюдается изменение давление отолитов на рецепторные клетки. Рецепторы полукружных каналов раздражаются в момент ускоренного или замедленного вращательного движения в какой-либо плоскости за счет движения эндолимфы.
При возбуждении вестибулярного аппарата возникают многочисленные рефлекторные реакции двигательного характера (вестибуломоторные), изменяющие работу внутренних органов (вестибуловегетативные), а также разнообразные ощущения (вестибулосенсорные реакции). Примером вестибуломоторных рефлексов является появление особого движения глаз (нистагма) после проведения вращательной пробы: глаза совершают ритмическое движение - сначала медленно движутся в сторону, противоположную вращению, а затем очень быстро - в сторону, совпадающую с направлением вращения. Вестибуловегетативные рефлексы проявляются в изменениях сердечного ритма, в сужении или расширении сосудов, падении артериального давления, усилении движений кишечника и желудка, рвоте и других сдвигах, возникающих во время или после раздражения вестибулярного аппарата. Вестибулосенсорные реакции проявляются в ощущении головокружения, нарушении ориентировки в окружающей среде, возникновении чувства тошноты.
Вестибулярный аппарат участвует в регуляции и перераспределении мышечного тонуса. Это особенно четко проявляется на животных с двусторонним или односторонним разрушением вестибулярного аппарата. Так, при двусторонней экстирпации этого аппарата у животных отмечается нарушение равновесия, невозможность сохранять вертикальное положение тела, при односторонней - отклонение головы в сторону операции (вследствие возбуждения рецепторов лабиринта на противоположной стороне), перераспределение мышечного тонуса - увеличение тонуса разгибателей и уменьшение тонуса сгибателей конечности на стороне операции.
Мышечно-суставная рецепция
Поступление в центральную нервную систему информации о положении тела в пространстве и степени сокращения каждой мышцы происходит при возбуждении проприорецепторов, находящихся в мышцах, сухожилиях, околосуставных сумках, надкостнице.
Проприорецепторами являются: мышечные веретена, находящиеся среди мышечных волокон; тельца Гольджи, расположенные в сухожилиях; пачиниевы тельца, обнаруженные в фасциях, покрывающих мышцы сухожилиях, связках и надкостнице. Показано, что изменение активности различных проприорецепторов происходит в момент сокращения или расслабления мышц. Так, возбуждение мышечных веретен отмечается при удлинении (в момент расслабления или растяжения) мышечных волокон. Тельца же Гольджи возбуждаются при сокращении мышечных волокон. При повышении активности мышечных веретен импульсы, поступающие от них в центральную нервную систему, облегчают сокращение данной мышцы и тормозят сокращение мышцы-антагониста. Импульсы, приходящие в центральную нервную систему от телец Гольджи, вызывают противоположную реакцию, т. е. тормозят сокращение данной мышцы и способствуют сокращению мышцы-антагониста.
Мышечные веретена всегда находятся в состоянии некоторого возбуждения, поэтому от мышечных веретен постоянно поступают нервные импульсы в центральную нервную систему, в спинной мозг. Это приводит к тому, что двигательные нервные клетки - мотонейроны - спинного мозга находятся в состоянии тонуса и непрерывно посылают редкие нервные импульсы по эфферентным путям к поперечнополосатым мышечным волокнам, обеспечивая их умеренное сокращение - тонус.
Орган вкуса и обоняния
Возникновение ощущения вкуса тесно связано с возбуждением вкусовых рецепторов, от которых нервные импульсы поступают в центральную нервную систему.
Рецепторы вкуса иначе называют вкусовыми почками. Они сосредоточены в основном на сосочках языка. Больше всего вкусовых рецепторов на кончике, краях и задней части языка. Вкусовые рецепторы не обнаружены на середине языка и его нижней поверхности. Рецепторы вкуса располагаются также на задней стенке глотки, мягком небе, миндалинах, надгортаннике.
По виду вкусовые почки напоминают луковицу. Каждая луковица состоит из собственной рецепторных (от 2 до 6) и опорных клеток. Рецепторные клетки являются специализированными эпителиальными клетками.
Вкусовые почки не достигают поверхности слизистой оболочки и отделены от нее небольшими углублениями - вкусовыми порами. Каждая вкусовая рецепторная клетка имеет микроворсинки. Они направлены к поверхности вкусовой поры. Считают, что различные химические вещества, входящие в состав пищи, раздражают микроворсинки вкусовых рецепторов, в результате чего они приходят в состояние возбуждения. Вкусовые рецепторы посылают нервные импульсы в центральную нервную систему по волокнам языкоглоточного и блуждающего нервов.
Ощущение вкуса может быть горьким, сладким, кислым и соленым. Имеется большая специализация вкусовых сосочков к восприятию химических раздражителей, обусловливающих неодинаковые вкусовые ощущения. Так, при локальном нанесении различных веществ на отдельные сосочки языка показано, что раздражение одних сосочков вызывает ощущение только сладкого вкуса, других - только горького и т. д. Вместе с тем имеются сосочки, возбуждение которых сопровождается двумя или тремя вкусовыми ощущениями.
Вкусовые сосочки сосредоточены преимущественно в определенных участках верхней поверхности языка. Так, ощущение сладкого воспринимается главным образом при раздражении передней части языка, горького - его основания.
Чтобы возникли вкусовые ощущения, раздражающее вещество должно находиться в растворенном состоянии. Об этом свидетельствует следующий опыт. Если вытереть насухо марлевой салфеткой язык и поместить на него порошкообразное вещество (соль или сахар), то вкусовое ощущение (соленого или сладкого) возникает не сразу, а через несколько минут, после того как вещество растворится в выделяющейся слюне.
Для возникновения ощущения вкуса имеют значение раздражения не только вкусовых, но и обонятельных рецепторов, а также тактильных, болевых и температурных рецепторов полости рта. За счет этого возникает ощущение "едкого", "вяжущего", "терпкого" вкуса.
Рецепторы обоняния сосредоточены в области верхних носовых ходов. Они находятся между цилиндрическими опорными клетками. На поверхности обонятельных клеток имеются реснички, которые постоянно находятся в движении. Это увеличивает возможность их контакта с молекулами пахучих веществ. Обонятельные рецепторы являются специализированным аппаратом, который возбуждается за счет свойств молекула пахучих веществ. Рецепторы обоняния очень чувствительны. Так, для получения ощущения запаха достаточно, чтобы было возбуждено 40 рецепторных клеток, причем на каждую из них должна действовать всего одна молекула пахучего вещества.
Ощущение запаха зависит от химической структуры и концентрации пахучего вещества в воздухе. Кроме того, на интенсивность ощущения влияет скорость тока воздуха через нос. Чем больше скорость поступления в нос воздуха с пахучим веществом, тем сильнее ощущение запаху
У обонятельных рецепторов наиболее выражена способность к адаптации, за счет которой снижается их чувствительность к действию пахучих веществ. Ощущение запаха при одной и той же концентрации пахучего вещества в воздухе возникает лишь в первый момент его действия на обонятельные клетки. В дальнейшем ощущение запаха ослабевает. Вместе с тем адаптация проявляется только по отношению к конкретному запаху и может не распространяться на другие пахучие вещества.
Количество слизи в полости носа также влияет на возбудимость обонятельных рецепторов. При повышенном выделении слизи, например во время насморка, происходит снижение чувствительности рецепторов обоняния к пахучим веществам.
Тактильная, температурная и болевая чувствительность
Тактильные рецепторы находятся на поверхности кожи и слизистых оболочках полости рта и носа. Они возбуждаются при прикосновении к ним или давлении на них.
К тактильным рецепторам относят мейснеровы тельца, расположенные в сосудах кожи, и меркелевы диски, имеющиеся в большом количестве на кончиках пальцев и губах. К рецепторам давления относят тельца Пачини, которые сосредоточены в глубоких слоях кожи, в сухожилиях, связках, брюшине, брыжейке кишечника.
Нервные ИМПУЛЬСЫ, возникшие в тактильных рецепторах, по чувствительным волокнам поступают в заднюю центральную извилину коры головного мозга.
В различных местах кожи тактильная чувствительность проявляется в неодинаковой степени. Так, она наиболее высока на поверхности губ, носа, языка, а на спине, подошве стоп, животе выражена в меньшей степени. Показано, что одновременное прикосновение к двум точкам кожи не всегда сопровождается возникновением ощущения двух воздействий. Если указанные точки лежат очень близко друг к другу, то возникает ощущение одного прикосновения. Наименьшее расстояние между точками кожи, при раздражении которых возникает ощущение двух прикосновении, называют порогом пространства. Пороги пространства неодинаковы в различных местах кожи: они минимальны на кончиках пальцев, губах и языке и максимальны на бедре, плече, спине.
Температура окружающей среды возбуждает терморецепторы, сосредоточенные в коже, на роговой оболочке глаза, в слизистых оболочках. Изменение температуры внутренней среды организма приводит к возбуждению температурных рецепторов, расположенных в гипоталамусе.
Различают терморецепторы, воспринимающие холод и тепло. Тепловые рецепторы представлены тельцами Руффини, холодовые - колбочками Краузе. Голые окончания афферентных нервных волокон также могут выполнять функции Холодовых и тепловых рецепторов.
Терморецепторы в коже располагаются на разной глубине: более поверхностно находятся холодовые, глубже - тепловые рецепторы. Вследствии этого время реакции на холодовые раздражения меньше, чем на тепловые. Терморецепторы сгруппированы в определенных точках поверхности тела человека, при этом холодовых точек значительно больше, чем тепловых. Выраженность ощущения тепла и холода зависит от места наносимого раздражения, величины раздражаемой поверхности и окружающей температуры.
Болевые ощущения возникают при действии любых раздражителе чрезмерной силы. Однако в настоящее время окончательно не установлено, какие рецепторы воспринимают боль. Одни исследователи считают, что особых рецепторов, воспринимающих боль, не существует, другие полагают, что возникновение боли связано с раздражением окончаний особых нервных волокон. Получены данные, свидетельствующие о том, что в формировании боли имеет значение образование в нервных окончаниях гистамина. Так, при подкожном введении гистамина в очень малой концентрации появляется ощущение боли. Возникновение боли связывают также с другими веществами, образующимися в тканях в месте повреждения. Такими веществами, в частности, являются брадикинин, XII фактор свертывания крови (фактор Хагемана).
Висцеральная чувствительность. Интерорецепторы, находящиеся во внутренних органах, получили название висцерорецепторов.
Висцерорецепторы имеют низкий порог раздражения. Они обладают большой специфичностью по отношению к действующим на них раздражителям. Во внутренних органах имеются рецепторы, реагирующие на механические раздражения (механорецепторы), на действие химических веществ (хеморецепторы), на сдвиги температуры внутренней среды организма (терморецепторы) и на изменение осмотического давления (осморецепторы).
Висцерорецепторы участвуют в регуляции работы внутренних органов, осуществляют рефлекторные взаимодействия между ними. Раздражение рецепторов внутренних органов в условиях нормы не сопровождается возникновением осознаваемых ощущений. Однако при возбуждении некоторых висцерорецепторов, например рецепторов мочевого пузыря и прямой кишки в случае растяжения их стенок, возникают ощущения позыва на мочеиспускание и дефекацию.
При использовании материалов сайта активная ссылка обязательна:
http://anfiz.ru/ 'Анатомия и физиология человека'