Понятие о двигательном аппарате
Приспособление организма человека и животных к постоянно меняющимся условиям внешней среды осуществляется за счет сложнейших рефлекторных реакций, важное место среди которых занимают двигательные акты. Перемещение в пространстве, нахождение пищи, трудовая деятельность, защита от врагов, продолжение рода невозможны без мышечной работы. В осуществлении ряда вегетативных функций, таких, как пищеварение, дыхание, кровообращение и т. д., всегда принимают участие мышцы.
Роль двигательной активности очень хорошо описал И. М. Сеченов в своей работе "Рефлексы головного мозга": "Все бесконечное разнообразие внешних проявлений мозговой деятельности сводится окончательно к одному лишь явлению - мышечному движению..."*.
* (Сеченов И. М. Рефлексы головного мозга. - М.: Изд-во АН СССР, 1961, с. 5.)
Двигательная деятельность выполняется специальной системой тканей и органов, получившей название двигательного аппарата.
Двигательный аппарат позвоночных животных состоит из сочлененных костей скелета и сложной системы поперечнополосатых мышц, приводящих в движение костные рычаги. С функциональной точки зрения, к двигательному аппарату должны быть отнесены мотонейроны и их аксоны, проводящие нервные импульсы к мышечным волокнам. Частью двигательного аппарата являются и мионевральные синапсы.
Нервно-мышечная часть двигательного аппарата получила название нервно-мышечной системы организма.
Нейромоторная единица. Анатомической и функциональной единицей скелетных мышц является нейромоторная единица. Установлено, что аксон (длинный отросток мотонейрона спинного мозга), войдя в мышцу, ветвится на множество веточек, каждая из которых иннервирует определенное количество мышечных волокон. Таким образом, под нейромоторной единицей следует понимать двигательный нейрон и иннервируемую им группу мышечных волокон (рис. 64).
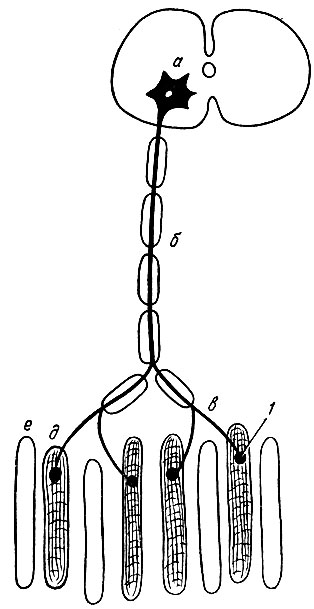
Рис. 64. Схема строения нейромоторной единицы. а - тело двигательной нервной клетки; б - двигательное нервное волокно; в - его разветвления: 1 - нервно-мышечное окончание; д - мышечные волокна, иннервируемые данной нервной клеткой; е - мышечные волокна, иннервируемые другими нервными клетками
В состав нейромоторной единицы может входить различное количество мышечных волокон, что зависит от специализации мышцы. Чем тоньше работа, тем меньшее количество мышечных волокон включено в нейромоторную единицу. Так, в составе нейромоторных единиц мышц глазного яблока обнаруживают три-четыре волокна, в мышцах же спины - несколько тысяч мышечных волокон. Двигательная единица работает как единое целое, импульсы, посылаемые мотонейроном, приводят в действие все образующие ее мышечные волокна.
Виды мышц. У позвоночных животных и человека существует три вида мышц: поперечнополосатые мышцы скелета, поперечнополосатая мышца сердца и гладкие мышцы внутренних органов, сосудов, кожи.
Основная функция скелетных мышц - сократительная, лежит в основе бесконечно разнообразной двигательной деятельности человека. За счет сократительной функции осуществляется перемещение тела в пространстве и поддержание определенной его позы. Скелетные мышцы выполняют рецепторную функцию, являясь своеобразным органом чувств. В толще мышц и сухожилий расположены многочисленные рецепторы. Наиболее изученными являются проприорецепторы (собственно рецепторы мышц), которые реагируют на растяжение и сокращение мышц. Кроме того, в мышцах обнаружены хемо- и терморецепторы. За счет сокращения мимических мышц проявляются те или иные эмоциональные состояния человека. Мышцы являются депо воды и солей и, следовательно, участвуют в регуляции водно-солевого обмена в организме. При сокращении мышцы часть энергии АТФ переходит в тепловую энергию, тем самым мышцы участвуют в терморегуляции. Наряду с клетками печени мышцы являются депо гликогена. В мышечной ткани осуществляются процессы синтеза и ресинтеза гликогена, АТФ, креатинфосфата. Наконец, мышцы являются депо кислорода за счет миогемоглобина, который находится в мышечных клетках. Кислород миоглобина используется в случае интенсивной физической нагрузки.
Скелетные мышцы состоят из большого количества мышечных волокон (мышечных клеток). Волокно поперечнополосатой мышцы имеет вытянутую форму, диаметр его от 10 до 100 мкм, длина волокна от нескольких сантиметров до 10·10-2-12·10-2 м (10-12 см). Мышечная клетка окружена тонкой электрогенной мембраной - сарколеммой, содержит саркоплазму (протоплазму) и многочисленные ядра. От электрогенной мембраны отходят элементы проводящей системы мышечного волокна. Сократительной частью мышечного волокна являются длинные мышечные нити - миофибриллы, проходящие внутри волокна от одного конца до другого и имеющие поперечную исчерченность. Миофибриллы очень тонки - диаметр их около 1-1,7 мкм, они отделены друг от друга тонким слоем протоплазмы.
Под световым микроскопом миофибриллы представляют собой образования, состоящие из правильно чередующихся между собой дисков А и I. Диски А - анизотропные, обладают двойным лучепреломлением, темные диски состоят из нитей сократительных белков - актина и миозина. Диски I - изотропные, не обладают двойным лучепреломлением, в светлых дисках обнаруживаются только нити актина. В средней части саркомера расположены диски А, по периферии-диски I (рис. 65).
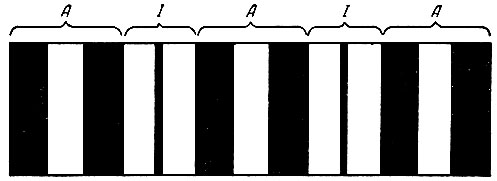
Рис. 65. Схема строения миофибриллы. Объяснение в тексте
Деятельность поперечнополосатой мускулатуры регулируется центральной нервной системой и ее ведущим отделом - корой головного мозга. Мышца снабжена чувствительными, двигательными и симпатическими нервными волокнами. Через двигательные окончания мышцам передаются импульсы от мотонейронов спинного мозга или клеток коры головного мозга. Эти импульсы приводят мышечные волокна в действие. Окончания чувствительных волокон связаны с проприорецепторами, расположенными в мышцах, сухожилиях, суставах, связках. Через симпатические нервные волокна центральная нервная система регулирует процессы обмена веществ в мышце и влияет таким образом на ее функциональное состояние.
В период относительного покоя скелетные мышцы полностью не расслабляются и сохраняют умеренную степень напряжения. Умеренную степень напряжения мышцы в состоянии покоя называют мышечным тонусом. Причиной мышечного тонуса являются поступающие к мышце редкие нервные импульсы от мотонейронов передних рогов спинного мозга, которые попеременно возбуждают различные нейромоторные единицы. Ритмическая активность мотонейронов поддерживается за счет расположенных выше нервных центров, а также нервных импульсов, поступающих от проприорецепторов мышц.
Основные физиологические свойства скелетных мышц. Мышечная ткань, как и все возбудимые ткани, обладает следующими физиологическими свойствами: возбудимостью, проводимостью, рефрактерностью, лабильностью. Специфическим свойством мышечной ткани является сократимость.
Возбудимость мышечной ткани ниже, чем нервной. Возбуждение, возникшее в каком-либо участке мышечного волокна, распространяется вдоль него. Скорость распространения возбуждения в мышечной ткани намного ниже, чем в нервной. Так, скорость распространения потенциала действия в поперечнополосатых мышцах теплокровных составляет около 5 м/с, а в двигательных нервных волокнах - 80-120 м/с.
Рефрактерный период мышечной ткани более продолжителен, чем нервный. Длительность рефрактерного периода нерва 14 м/с, а поперечнополосатой мышцы около 35 м/с.
Лабильность мышечной ткани значительно ниже, чем нервной. Действительно, икроножная мышца лягушки может воспроизводить 200-250 волн возбуждения в 1 с, а седалищный нерв - 500-1000.
Под сократимостью следует понимать способность мышечного волокна изменять свою длину и степень напряжения в ответ на раздражение пороговой силы. Скелетные мышцы могут работать в режиме изотонического и изометрического сокращения.
При изотоническом сокращении в основном изменяется длина мышечного волокна. Степень же напряжения его не претерпевает существенных изменений.
При изометрическом сокращении значительно возрастает напряжение мышечного волокна, длина же его практически не изменяется.
В целостном организме сокращение мышц никогда не бывает чисто изотоническим или чисто изометрическим, так как мышцы, поднимая груз (например, сгибая конечность в суставе), укорачиваются и вместе с тем меняют степень своего напряжения.
Одиночное мышечное сокращение, его фазы. Характер сокращения скелетной мышцы зависит от частоты раздражения (или от частоты поступления нервных импульсов). Различают одиночное и тетаническое сокращение мышц.
Раздражение мышцы или иннервирующего ее двигательного нерва одиночным стимулом вызывает одиночное мышечное сокращение (рис. 66). Запись кривой одиночного мышечного сокращения производят при помощи быстровращающегося кимографа, кривая имеет вид, представленный на рис. 66. Сокращение начинается не тотчас же после нанесения раздражения, а через определенный промежуток времени, который называют латентным, или скрытым, периодом возбуждения. Следовательно, латентный период - это время, прошедшее от нанесения раздражения до момента начала механической реакции мышцы. Латентный период одиночного сокращения икроножной мышцы лягушки составляет 0,01 с. Фаза сокращения длится 0,04 с, период расслабления более продолжителен, чем сокращение, и составляет 0,05 с. Продолжительность всего одиночного мышечного сокращения икроножной мышцы лягушки 0,10-0,12 с.
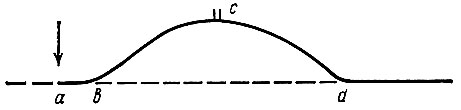
Рис. 66. Кривая одиночного мышечного сокращения икроножной мышцы лягушки. а - момент нанесения раздражения; b - начало сокращения; аb - латентный период; bc - фаза сокращения; cd - фаза расслабления
Суммация мышечных сокращений, зубчатый и гладкий тетанус. В естественных условиях к мышечным волокнам поступают не одиночные нервные импульсы, а их ряды. Когда мышца получает ряд часто следующих друг за другом нервных импульсов, она отвечает длительным сокращением. Длительное, слитное сокращение мышцы получило название тетанического сокращения, или тетануса. К тетаническому сокращению способны только скелетные мышцы. Гладкие мышцы и поперечнополосатая мышца сердца неспособны к тетаническому сокращению вследствие наличия продолжительного рефрактерного периода.
Тетанус возникает вследствие суммации одиночных мышечных сокращений. Для того чтобы возник тетанус, необходимо действие повторных раздражений (или нервных импульсов) на мышцу еще до того, как закончится ее одиночное сокращение.
Если раздражающие импульсы сближены и каждый из них приходится на тот момент, когда мышца только начала расслабляться, но не успела еще полностью расслабиться, то возникает зубчатый тип сокращения, который получил название неполного, несовершенного тетануса (клонус) (рис. 67).
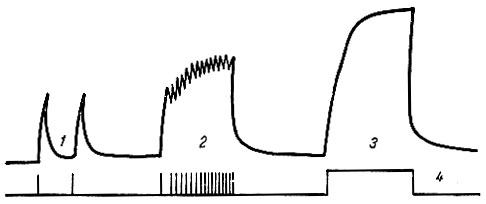
Рис. 67. Сокращения икроножной мышцы лягушки при разной частоте раздражений. 1 - одиночные сокращения; 2 - зубчатый тетанус; 3 - гладкий тетанус; 4 - отметки нанесения раздражений
Если раздражающие импульсы сближены настолько, что каждый последующий приходится на время, когда мышца еще не успела перейти к расслаблению от предыдущего раздражения, т. е. происходит на высоте ее сокращения, то возникает длительное непрерывное сокращение, получившее название гладкого, совершенного тетануса (см. рис. 67).
При анализе рис. 67 обращает на себя внимание следующая особенность: амплитуда зубчатого и гладкого тетануса выше, чем амплитуда одиночного мышечного сокращения. Н. Е. Введенский показал, что увеличение амплитуды тетануса связано с тем, что каждый последующий нервный импульс попадает в стадию экзальтации, которая возникает после предыдущей волны возбуждения.
Совершенный тетанус - нормальное рабочее состояние скелетных мышц, обусловливается посылкой из центральной нервной системы нервных импульсов с частотой 40-50 в 1 с. Зубчатый тетанус возникает при частоте нервных импульсов до 30 в 1 с. Если мышца получает 10-20 нервных импульсов в 1 с, то она находится в состоянии мышечного тонуса, т. е. умеренной степени напряжения.
Тетаническое сокращение скелетных мышц имеет преимущества перед одиночным сокращением: оно сильнее и продолжительнее, что дает возможность сохранить определенное положение тела, держать груз и т. д.
Кроме тетанического сокращения, встречается еще одна разновидность длительного сокращения мышц, которая получила название контрактуры. Контрактура продолжается и при снятии раздражителя. Контрактура мышцы наступает при нарушении обмена веществ или изменении свойств сократительных белков мышечной ткани.
Значение тетанического сокращения мышц в трудовой деятельности человека чрезвычайно велико, о чем свидетельствует образное выражение физиолога А. Ф. Самойлова: "...все, что составляет наше богатство в индустрии и в искусстве, ...все, что сделано "руками" человека, сделано тетаническим сокращением мышцы... только тетанусом мышца и служит нам: тетанус и есть миссия мышцы"*.
* (Самойлов А. Ф. Избранные статьи и речи. - М.-Л., 1940, с 307.)
Химические превращения в мышцах при сокращении. В основе мышечного сокращения лежит превращение химической энергии в механическую. Химические процессы, совершающиеся в мышце, могут происходить без кислорода и с его участием.
Бескислородная (анаэробная) фаза характеризуется процессом распада аденозинтрифосфорной кислоты (АТФ) и креатинфосфата и их восстановлением (ресинтезом).
АТФ распадается на аденозиндифосфорную (АДФ) и фосфорную кислоты. Выделяющаяся при этом энергия расходуется на мышечное сокращение, а фосфорная кислота используется на фосфорилирование глюкозы с образованием гексозофосфатов. Креатинфосфат распадается на креатин и фосфорную кислоту. Энергия, освобождающаяся при этом, и фосфорная кислота используются на восстановление АДФ и АТФ.
На последнем этапе гексозофосфаты расщепляются на молочную и фосфорную кислоты. Энергия этого распада и освобождающаяся фосфорная кислота необходимы для ресинтеза креатинфосфата.
Таким образом, анаэробные процессы при мышечном сокращении представляют собой цепь последовательного распада и ресинтеза макроэргических фосфорных соединений. При этом энергия для мышечного сокращения черпается при распаде АТФ. Все последующие реакции обеспечивают восстановление макроэргических фосфорных соединений.
Кислородная (аэробная) фаза химических превращений связана с процессами окисления молочной кислоты до углекислого газа и воды. При этом распадается только 1/5 часть молочной кислоты. Выделяющаяся энергия используется для превращения оставшейся части мелочной кислоты в глюкозу и далее в гликоген.
Следовательно, несмотря на цепь сложнейших химических превращений, затраты организма при мышечном сокращении сводятся в конечном итоге к потере части гликогена.
Образование тепла в мышцах при сокращении. В процессе сокращения не вся химическая энергия переходит в механическую, 40% ее превращается в тепловую.
Процессы теплообразования в мышцах в покое и при сокращении были подробно изучены английским физиологом Хиллом с помощью разработанного им высокочувствительного термоэлектрического метода. В процессе сокращения теплообразование протекает в две фазы. Первая получила название фазы начального теплообразования, вторая - запаздывающего, или восстановительного, те преобразования. Начальное тепло связано с процессами укорочения и расслабления мышц. Источником начального теплообразования являются химические процессы анаэробного распада углеводов. Восстановительное теплообразование происходит в течение нескольких минут после расслабления и обусловлено в основном окислительными процессами. Тепло, образующееся в мышцах, обеспечивает оптимальные условия для течения ферментных процессов.
Физиологические особенности гладких мышц. Гладкие мышцы образуют стенки (мышечный слой) внутренних органов и кровеносных сосудов.
Микроскопическое строение гладких мышц отличается от строения поперечнополосатых мышц тем, что в миофибриллах гладких мышц нет поперечной исчерченности. Это обусловлено хаотичным расположением сократительных белков в волокнах гладких мышц. Волокна гладких мышц относительно коротки - от 50 до 200 мкм.
Физиологические свойства гладких мышц в связи с особенностями их строения и уровня обменных процессов значительно отличаются от физиологических свойств поперечнополосатых мышц.
Гладкие мышцы менее возбудимы, чем поперечнополосатые. Возбуждение по ним распространяется с небольшой скоростью - от 2·10-2 до 15·10-2 м/с (2-15 см/с). Возбуждение в гладких мышцах может передаваться с одного волокна на другое в отличие от нервных волокон и волокон поперечнополосатых мышц.
Сокращение гладкой мускулатуры происходит более медленно и длительно. Так, сокращение гладкой мускулатуры кишечника кролика может продолжаться до 5 с, еще более медленно протекает расслабление. Вследствие большой продолжительности сократительного акта гладкая мышца даже под влиянием редких раздражителей может переходить в состояние длительного сокращения, которое напоминает тетанус скелетных мышц. Характерными для гладких мышц являются также длительные тонические сокращения.
Рефрактерный период в гладких мышцах более продолжителен, чем в скелетных (до нескольких секунд).
Важным свойством гладкой мышцы является ее большая пластичность, т. е. способность сохранять приданную растяжением длину без изменения напряжения. Данное свойство имеет существенное значение, так как некоторые органы брюшной полости (матка, мочевой пузырь, желчный пузырь) иногда значительно растягиваются.
Характерной особенностью гладких мышц является их способность к автоматической деятельности, которая обеспечивается нервными элементами, заложенными в стенках гладкомышечных органов.
Адекватным раздражителем для гладких мышц является их быстрое и сильное растяжение, что имеет большое значение для функционирования многих гладкомышечных органов (кишечник, мочеточники и другие полые органы).
Особенностью гладких мышц является также их высокая чувствительность к некоторым биологически активным веществам (ацетилхолин, адреналин, норадреналин, серотонин и др.).
Гладкие мышцы иннервируются симпатическими и парасимпатическими вегетативными нервами, которые, как правило, оказывают противоположное влияние на их функциональное состояние.
При использовании материалов сайта активная ссылка обязательна:
http://anfiz.ru/ 'Анатомия и физиология человека'