Глава X. Физиология нервно-мышечной системы
Понятие о возбудимых тканях. Возбуждение, возбудимость
Все живые ткани и клетки под влиянием раздражителей переходят из состояния физиологического покоя в состояние активности. Степень активного состояния живой ткани может быть различной.
Наиболее яркая ответная реакция на действие раздражителей наблюдается со стороны нервной и мышечной ткани, менее выражена она в железистой и соединительной ткани.
Основными физиологическими свойствами нервной и мышечной ткани являются: возбудимость, проводимость, рефрактерность, лабильность. Специфическим свойством мышечной ткани является сократимость.
Возбудимость - способность живой ткани отвечать на действие раздражителя изменением физиологических свойств и возникновением процесса возбуждения.
Возбуждение - это активный физиологический процесс, который возникает в ткани под действием раздражителей и характеризуется рядом общих и специфических признаков.
К общим признакам возбуждения, которые присущи всем возбудимым тканям, относятся: изменение уровня обменных процессов в тканях, выделение различных видов энергии - тепловой, электрической, а по некоторым данным, и лучистой.
Специфические признаки возбуждения проявляются в жизнедеятельности живых структур. Так, например, специфическим признаком возбуждения мышечной ткани является сокращение, железистой - выделение секрета, нервной - генерация нервного импульса.
Проводимостью называют способность живой ткани проводить волны возбуждения, точнее, электрические токи, которые получили название биопотенциалов.
Рефрактерность - это временное снижение возбудимости ткани, возникающее в результате возбуждения.
Все живые ткани в зависимости от особенностей обменных процессов могут возбуждаться в единицу времени определенное количество раз. Указанную способность тканей Н. Е. Введенский назвал лабильностью или функциональной подвижностью.
Таким образом, все живые ткани обладают рядом общих физиологических свойств. Универсальным свойством всего живого следует считать возбудимость. Различают две формы возбуждения: местное нераспространяющееся и импульсное, волнообразно распространяющееся.
При местном возбуждении активируются только отдельные структуры возбудимого образования. Местное возбуждение - это пусковой механизм для возникновения распространяющегося возбуждения. В этом его биологическое значение.
В процессе эволюции возбуждение стало распространяющимся, импульсным. Биологическое значение распространяющегося возбуждения заключается в том, что оно обеспечивает функциональное отправление раздражаемого образования, активируя его в целом.
Раздражители под пороговые, пороговые, надпороговые. Раздражитель - причина, способная вызвать ответную реакцию со стороны возбудимых тканей. Раздражители делят на электрические, химические, механические, температурные.
По биологическому признаку раздражители могут быть адекватными и неадекватными. Адекватные раздражители воздействуют на возбудимые системы в естественных условиях существования организма. Так, адекватным раздражителем для фоторецепторов сетчатки глаза являются световые лучи (кванты света). Неадекватные раздражители в естественных условиях существования организма не воздействуют на возбудимые структуры. Однако при достаточной силе и продолжительности могут вызвать ответную реакцию со стороны возбудимых тканей.
В условиях физиологического эксперимента в качестве раздражителя чаще всего используют электрический ток. Электрический ток легко дозировать, и он является адекватным раздражителем для возбудимых тканей, так как функциональная их активность всегда сопровождается электрическими явлениями.
По своей силе раздражители могут быть подпороговыми, пороговыми, надпороговыми.
Подпороговый раздражитель - это раздражитель такой силы, который не вызывает видимых изменений, но обусловливает возникновение физико-химических сдвигов в возбудимых тканях. Однако степень этих сдвигов недостаточна для возникновения распространяющегося возбуждения.
Пороговый раздражитель - это раздражитель минимальной силы, который впервые вызывает видимую ответную реакцию со стороны возбудимой ткани. Пороговую силу раздражителя называют порогом раздражения или возбуждения. Порог раздражения и является мерой возбудимости ткани. Между порогом раздражения и возбудимостью существует обратная зависимость: чем выше порог раздражения, тем ниже возбудимость, чем ниже порог раздражения, тем возбудимость выше.
Надпороговый раздражитель - это раздражитель, сила которого выше, чем сила порогового раздражителя.
Следует отметить, что порог раздражения - показатель достаточно изменчивый и значительно зависит от исходного функционального состояния возбудимой ткани.
Законы раздражения. Для возникновения возбуждения решающее значение имеет сила раздражителя. Чем больше сила раздражителя, тем выше, до определенного предела, ответная реакция со стороны возбудимой ткани - закон силы раздражения. Однако не меньшее значение имеет и длительность действия раздражителя. Французский ученый Лапик создал учение о хронаксии как пороговом времени, пороговой длительности раздражения, необходимой для возникновения возбуждения в живой ткани.
Зависимость между силой раздражителя и продолжительностью его воздействия, необходимого для возникновения минимальной ответной реакции живой структуры, очень хорошо можно проследить на так называемой кривой силы - времени (кривая Гоорвега-Вейса-Лапика) (рис. 51).
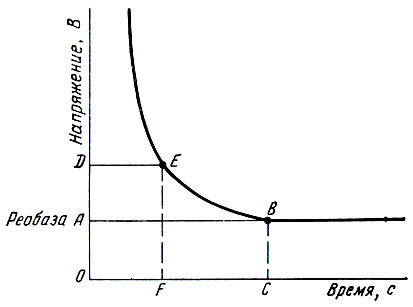
Рис. 51. Кривая сила - время. Объяснение в тексте
Из анализа кривой следует, что, как бы ни велика была сила раздражителя, при недостаточной длительности его воздействия ответной реакции не будет (точки слева от восходящей ветви гиперболы). Аналогичное явление наблюдается при продолжительном действии подпороговых раздражителей. Минимальная сила тока (или напряжения), способная вызвать возбуждение, названа Лапиком реобазой (отрезок ординаты ОА). Наименьший промежуток времени, в течение которого ток, равный по силе удвоенной реобазе, вызывает в ткани возбуждение, называют хронаксией (отрезок абсциссы OF), которая представляет собой показатель пороговой длительности раздражения. Хронаксия измеряется в δ (тысячные доли секунды). По величине хронаксии можно судить о скорости возникновения возбуждения в ткани: чем меньше хронаксия, тем быстрее возникает возбуждение. Хронаксия нервных и мышечных волокон человека равна тысячным и десятитысячным долям секунды, а хронаксия так называемых медленных тканей, например мышечных волокон желудка лягушки, - сотым долям секунды.
Определение хронаксии возбудимых тканей получило широкое распространение не только в эксперименте, но и в физиологии спорта, в клинике. В частности, путем измерения хронаксии мышцы невропатолог может установить наличие повреждения двигательного нерва. Необходимо отметить, что раздражитель может быть достаточно сильным, иметь пороговую длительность, но низкую скорость нарастания во времени до пороговой величины, возбуждение в этом случае не возникает. Приспособление возбудимой ткани к медленно нарастающему раздражителю получило название аккомодации. Аккомодация обусловлена тем, что за время нарастания силы раздражителя в ткани успевают развиться активные изменения, повышающие порог раздражения и препятствующие развитию возбуждения. Таким образом, скорость нарастания раздражения во времени, или градиент раздражения, имеет существенное значение для возникновения возбуждения.
Закон градиента раздражения. Реакция живого образования на раздражитель зависит от градиента раздражения, т. е. от срочности или крутизны нарастания раздражителя во времени: чем выше градиент раздражения, тем сильнее (до определенных пределов) ответная реакция возбудимого образования.
Следовательно законы раздражения отражают сложные взаимоотношения между раздражителем и возбудимой структурой при их взаимодействии. Для возникновения возбуждения раздражитель должен иметь пороговую силу, обладать пороговой длительностью и иметь определенную скорость нарастания во времени.
Биоэлектрические явления в живых тканях
Потенциал покоя. Между наружной поверхностью клетки и ее внутренним содержимым (протоплазмой) можно обнаружить разность потенциалов около 60-90 мВ. При этом поверхность клетки заряжена электроположительно по отношению к содержимому (протоплазме). Эту разность потенциалов называют потенциалом покоя, или мембранным потенциалом. Зарегистрировать мембранный потенциал можно с помощью микроэлектродов, предназначенных для внутриклеточного отведения биопотенциалов (рис. 52).
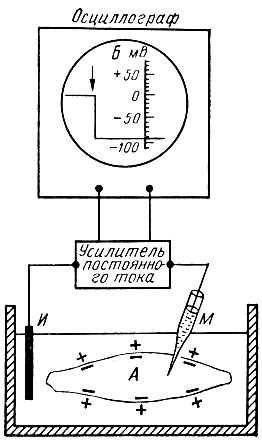
Рис. 52. Измерение мембранного потенциала мышечного волокна (А) с помощью внутриклеточного микроэлектрода (схема). М - микроэлектрод; И - индифферентный электрод. Луч на экране осциллографа (Б) показывает, что до прокола микроэлектродом мембраны разность потенциалов М и И была равна 0. В момент прокола (показан стрелкой) обнаружена разность потенциалов, свидетельствующая о том, что внутренняя сторона мембраны заряжена электроотрицательно по отношению к ее наружной поверхности
Ток покоя можно обнаружить, используя более простой методический прием. Один из электродов помещают на неповрежденную поверхность нервного или мышечного волокна, а другой - на поврежденный участок. Электроды соединяют с электроизмерительным прибором, который зарегистрирует разность потенциалов или ток покоя между поврежденным, заряженным электроотрицательно, и неповрежденным, электроположительным участком ткани.
Потенциал действия. При нанесении на участок нервного или мышечного волокна раздражителя достаточной силы и длительности возникает возбуждение, наиболее важным проявлением которого является быстрое колебание мембранного потенциала. При этом возбужденный участок заряжается электроотрицательно по отношению к невозбужденному.
Потенциал действия можно зарегистрировать двумя способами: с помощью электродов, приложенных к внешней поверхности волокна (внеклеточное отведение), и с помощью микроэлектрода, введенного внутрь протоплазмы (внутриклеточное отведение).
При внеклеточном способе регистрации ток действия имеет двухфазную структуру. Если приложить электроды осциллографа к неповрежденной поверхности нервного или мышечного волокна, разность потенциалов не обнаружится, так как неповрежденная поверхность волокна заряжена электроположительно. При нанесении раздражения к области С возникает потенциал действия. Волна электроотрицательности в первую очередь достигнет области электрода А, что сопровождается перемещением луча на экране осциллографа. В следующий момент волна электроотрицательности покидает область электрода А и распространяется в межэлектродной области. Разность потенциалов между электродами А и В не регистрируется и луч на экране осциллографа возвращается в исходное положение. При достижении волной электроотрицательности области электрода В между электродами вновь возникнет разность потенциалов, но противоположного знака. В результате появления разности потенциалов обратного знака луч на экране осциллографа отклонится вниз от нулевого уровня. Однако возбуждение продолжает распространяться и волна электроотрицательности покидает область электрода В. Разность потенциалов между электродами А и В исчезает, луч на экране осциллографа вновь возвращается в исходное положение (рис. 53).
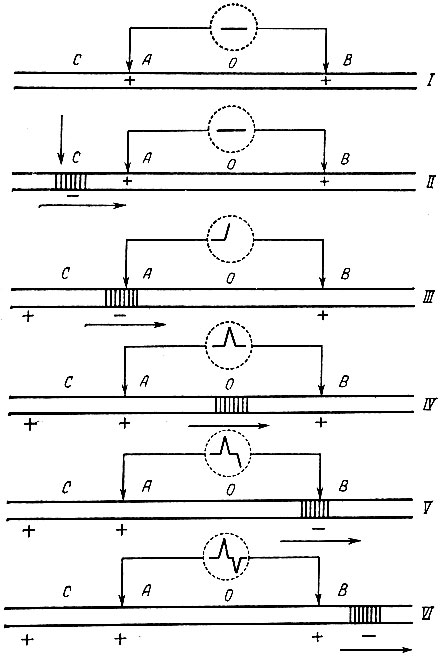
Рис. 53. Схема, объясняющая возникновение двухфазного тока действия. А и В - электроды осциллографа; О - осциллограф. Стрелкой показано место и момент нанесения раздражения; + и - характеризуют электрический заряд нерва в данный момент. Изменение заряда приводит к изменению положения луча на экране осциллографа
Регистрация потенциала действия с помощью электронных усилителей и микроэлектродов позволила выяснить достаточно сложную его структуру (рис. 54).
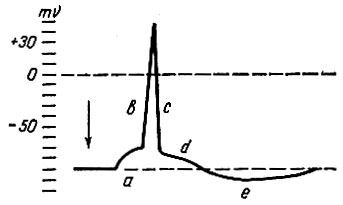
Рис. 54. Схема потенциала действия, зарегистрированного с помощью внутриклеточного микроэлектрода. а - местные колебания мембранного потенциала; b - восходящая часть пика потенциала действия (фаза деполяризации); с - нисходящая часть пика потенциала действия (фаза реполяризации); d - отрицательный следовой потенциал; е - положительный следовой потенциал. Стрелкой показан момент нанесения раздражения
В потенциале действия различают местные колебания мембранного потенциала, пик потенциала действия и следовые потенциалы - отрицательный и положительный. Пик потенциала действия представлен восходящей (В) и нисходящей (С) ветвями. Во время восходящей части пика потенциала действия заряд мембраны быстро снижается до нуля (деполяризация) и возникает разность потенциалов противоположного знака. В связи с этим восходящая часть пика получила название фазы деполяризации. В момент нисходящей части пика происходит восстановление исходного уровня поляризации (электрического состояния) мембраны. Вследствие этого эту часть потенциала действия называют фазой реполяризации. Следовые потенциалы связаны с восстановительными процессами, медленно развивающимися в нервных и мышечных волокнах по окончании процесса возбуждения. Продолжительность потенциала действия в нервных и мышечных волокнах составляет 0,1-5 мс, при этом фаза реполяризации всегда продолжительнее фазы деполяризации. Следовые потенциалы имеют низкую амплитуду - несколько милливольт и более продолжительны, чем потенциал действия. Длительность следовых потенциалов может колебаться в весьма широких пределах - от нескольких миллисекунд до нескольких десятков и даже сотен миллисекунд.
Происхождение потенциала покоя и потенциала действия. Предложено много теорий, объясняющих происхождение биопотенциалов. Наиболее полно экспериментально обоснована мембранная теория, предложенная немецким исследователем Бернштейном (1902, 1912). В современный период эта теория модифицирована и экспериментально разработана Ходжкиным, Хаксли, Катцем (1949-1952).
Установлено, что в основе биоэлектрических явлений лежит неравномерное распределение (асимметрия) ионов в цитоплазме клетки и окружающей ее среде. Так, протоплазма нервных и мышечных клеток содержит в 30-50 раз больше ионов калия, в 8-10 раз меньше ионов натрия и в 50 раз меньше ионов хлора, чем внеклеточная жидкость. Кроме того, в состав цитоплазмы клетки входят органические анионы (крупномолекулярные соединения, несущие отрицательный заряд), которые отсутствуют во внеклеточной среде.
Сторонники мембранной теории основной причиной ионной асимметрии считают наличие клеточной мембраны со специфическими свойствами.
Мембрана клетки - это уплотненный слой цитоплазмы, толщина которого около 10 нм (100 А). Использование электронно-микроскопических методов исследования позволило определить тонкую структуру мембраны (рис. 55). Клеточная мембрана состоит из двойного слоя молекул фосфолипидов, который покрыт изнутри слоем белковых молекул, а снаружи - слоем молекул сложных углеводов - мукополисахаридов. В мембране имеются специальные каналы - "поры", через которые вода и ионы проникают внутрь клетки. Предполагают, что для каждого иона имеются специальные каналы. В связи с этим проницаемость мембраны для тех или иных ионов будет зависеть от размеров пор и диаметров самих ионов.
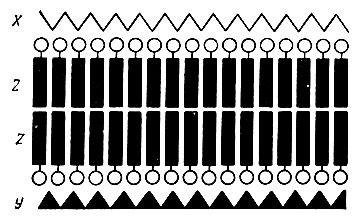
Рис. 55. Схема молекулярной структуры клеточной мембраны. Показан бимолекулярный липидный слой - Z (кружками обозначены полярные группы фосфолипидов) и два нелипидных монослоя: наружный - мукополисахаридный, Х; внутренний - белковый, Y
В состоянии относительного физиологического покоя мембрана обладает повышенной проницаемостью для ионов калия, проницаемость же ее для ионов натрия резко снижена.
Таким образом, особенности проницаемости клеточной мембраны, а также размеры самих ионов являются одной из причин, обеспечивающих асимметрию распределения ионов по обе стороны клеточной мембраны. Ионная же асимметрия - одна из основных причин возникновения потенциала покоя, при этом ведущая роль принадлежит неравномерному распределению ионов калия.
Ходжкиным выполнены классические опыты на гигантском нервном волокне кальмара. Выравнивали концентрацию ионов калия внутри волокна и в окружающей жидкости - потенциал покоя исчезал. Если же волокно заполнялось искусственным солевым раствором, близким по составу к внутриклеточной жидкости, между внутренней и наружной сторонами мембраны устанавливалась разность потенциалов, примерно равная потенциалу покоя нормального волокна (50-80 мВ).
Механизм возникновения потенциала действия значительно сложнее. Основная роль в возникновении токов действия принадлежит ионам натрия. При действии раздражителя пороговой силы проницаемость мембраны клетки для ионов натрия возрастает в 500 раз и превышает проницаемость для ионов калия в 10-20 раз. В связи с этим натрий лавинообразно устремляется в клетку, что приводит к перезарядке клеточной мембраны. Наружная поверхность заряжается отрицательно по отношению к внутренней. Происходит деполяризация клеточной мембраны, сопровождающаяся реверсией мембранного потенциала. Под реверсией мембранного потенциала понимают то количество милливольт (мВ), на которое потенциал действия превышает потенциал покоя. Восстановление исходного уровня мембранного потенциала (реполяризация) осуществляется за счет резкого снижения натриевой проницаемости (инактивация) и активного переноса ионов натрия из цитоплазмы клетки в окружающую среду.
Доказательства натриевой гипотезы потенциала действия также были получены Ходжкиным. Действительно, если потенциал действия имеет натриевую природу, то, варьируя концентрацию ионов натрия, можно изменить величину потенциала действия. Оказалось, что при замене 2/3 морской воды, которая является нормальной окружающей средой для гигантского аксона кальмара, на изотонический раствор декстрозы, т. е. при изменении концентрации натрия в окружающей среде на 2/3, потенциал действия уменьшается наполовину.
Таким образом, возникновение биопотенциалов является функцией биологической мембраны, обладающей избирательной проницаемостью. Величина потенциала покоя и потенциала действия обусловливается ионной асимметрией в системе клетка - среда.
Изменения возбудимости в разные фазы потенциала действия. Развитие в нервном или мышечном волокне потенциала действия сопровождается выраженными изменениями возбудимости, которые происходят по определенным фазам. Для их изучения нерв или мышцу подвергают действию двух коротких, но сильных электрических раздражителей, следующих друг за другом через определенные интервалы времени. Различают следующие фазы изменения возбудимости: 1 - начальное повышение возбудимости; 2 - рефрактерный период, который состоит из фаз абсолютной и относительной рефрактерности; 3 - супернормальный период, или фаза экзальтации и 4 - субнормальный период возбудимости (рис. 56).
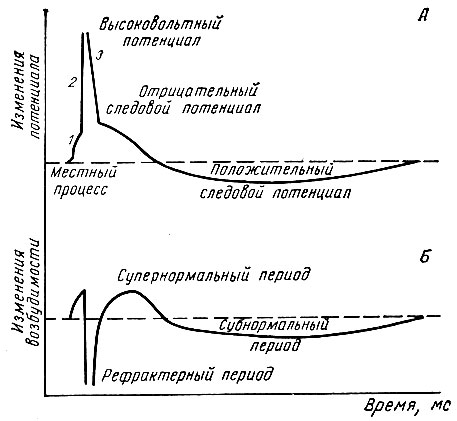
Рис. 56. Изменения возбудимости нервного волокна в различные фазы потенциала действия и следовых изменений мембранного потенциала (по Моргану). Для наглядности длительность первых двух фаз на каждой кривой несколько увеличена. Пунктирной линией на рисунке А обозначен потенциал покоя, на рисунке Б - исходный уровень возбудимости
Начальное повышение возбудимости совпадает по времени с местными колебаниями мембранного потенциала и связано с повышением проницаемости мембраны для всех ионов, но особенно для ионов натрия. Возникновение пика потенциала действия сопровождается резким снижением возбудимости ткани - состоянием рефрактерности. Стадия абсолютной рефрактерности совпадает по времени с восходящей частью спайка. В этот момент ткань не приходит в состояние возбуждения даже при действии надпороговых раздражителей. Возникновение абсолютной невозбудимости ткани связано с тем, что дальнейшее повышение натриевой проницаемости в этот период времени невозможно. Продолжительность абсолютной рефрактерности в нервном волокне составляет 0,002 с, в скелетной мышце теплокровных - 0,005 с. Относительный рефрактерный период совпадает с нисходящим коленом пика потенциала действия. Возбудимость в эту стадию постепенно восстанавливается и в ответ на надпороговые раздражители может вновь возникать потенциал действия. Указанное явление связано с восстановительными процессами на мембране и как следствие этого возможностью повышения натриевой проницаемости на надпороговый раздражитель. Длительность относительного рефрактерного периода в нервном волокне 0,012 с, в мышце 0,03 с. Относительная рефрактерность сменяется фазой экзальтации (Н. Е. Введенский), или супернормальной возбудимости. В этот период ткань отвечает возникновением возбуждения на раздражитель подпороговой (для исходного состояния) силы. Продолжительность фазы экзальтации в нервном волокне 0,02 с, в мышце 0,05 с. Период супернормальной возбудимости соответствует отрицательному следовому потенциалу.
Вслед за фазой экзальтации возникает стадия субнормальной возбудимости, совпадающая с положительным следовым потенциалом. Возбудимость в эту фазу незначительно снижена по сравнению с исходным ее уровнем.
Учение Н. Е. Введенского о функциональной подвижности (лабильности)
Лабильность, или функциональная подвижность, - одно из физиологических свойств живых тканей. Это свойство описано в 1892 г. Н. Е. Введенским, который установил, что скорость протекания процесса возбуждения в тканях различна. Каждая возбудимая ткань способна на раздражение отвечать только определенным количеством волн возбуждения. Так, нервное волокно способно воспроизводить до 1000 имп/с, поперечнополосатая мышца - 200-250 имп/с, мионевральный синапс - 100-125 имп/с (данные Н. Е. Введенского для нервно-мышечного препарата лягушки).
Мерой лабильности, по Н. Е. Введенскому, является то наибольшее количество волн возбуждения, которое возбудимая ткань может воспроизводить в 1 с в точном соответствии с ритмом наносимых раздражений без явлений трансформации (переделки) ритма.
Лабильность - величина весьма подвижная и может изменяться в достаточно широких пределах. В частности, лабильность широко варьирует в процессе ритмического раздражения. В одних случаях вследствие взаимодействия волн возбуждения лабильность может повыситься, в других - понизиться. Повышение лабильности может привести к тому, что недоступные ранее ритмы деятельности станут доступными - произойдет "усвоение ритма" (А. А. Ухтомский). Явление усвоения ритма играет важную роль в процессах врабатывания и тренировки. Снижение лабильности, происходящее в процессе деятельности, приводит к иному результату, способность ткани к ритмической работе уменьшается.
Лабильность может быть измерена косвенным путем по величине хронаксии возбудимых тканей. Чем короче хронаксия, тем выше лабильность. Определение лабильности весьма важно в физиологии труда и спорта.
При использовании материалов сайта активная ссылка обязательна:
http://anfiz.ru/ 'Анатомия и физиология человека'