Глава IX. Физиология эндокринной системы
Общая характеристика желез внутренней секреции
Все железы организма принято делить на две группы. К первой группе относят железы, имеющие выводные протоки и выполняющие внешнесекреторную функцию, - зкзокринные, ко второй группе - железы, не имеющие выводных протоков и выделяющие свой секрет непосредственно в межклеточные щели. Из межклеточных щелей секрет попадает в кровь, лимфу или цереброспинальную жидкость. Такие железы получили название эндокринных, или желез внутренней секреции.
Эндокринные железы расположены в разных частях организма и имеют разнообразную морфологическую структуру. Они развиваются из эпителиальной ткани, интерстициальных клеток, нейроглии и нервной ткани. Продукты деятельности желез внутренней секреции в отличие от секретов называют инкретами, или гормонами.
Термин "гормон" (от греч. hormao - двигаю, возбуждаю, побуждаю) предложен английскими физиологами Бейлисом и Старлингом (1905), которые выделили из слизистой оболочки двенадцатиперстной кишки специальное вещество - секретин, способствующее образованию поджелудочного сока.
Гормоны вырабатываются в эндокринных железах двух типов: 1) железах со смешанной функцией, осуществляющих наряду с внутренней и внешнюю секрецию; 2) железах, выполняющих только функцию органов внутренней секреции. К первой группе относят половые железы - гонады - и поджелудочную железу, ко второй - гипофиз, эпифиз, щитовидную, околощитовидную, вилочковую железы и надпочечники.
Гормоны - это химические соединения, обладающие высокой биологической активностью и в малых количествах дающие значительный физиологический эффект.
Железы внутренней секреции обильно снабжены рецепторами и иннервируются вегетативной нервной системой. По химической природе гормоны делят на три группы: 1) полипептиды и белки; 2) аминокислоты и их производные; 3) стероиды.
Гормоны циркулируют в крови в свободном состоянии и в виде соединений с белками. В связи с белками гормоны, как правило, переходят в неактивную форму.
Свойства гормонов. 1) Дистантный характер действия. Органы и системы, на которые действуют гормоны, обычно расположены далеко от места их образования в эндокринных железах. Так, в гипофизе, расположенном у основания мозга, продуцируются тропные гормоны, действие которых реализуется в щитовидной и половых железах, а также в надпочечниках. Женские половые гормоны образуются в яичнике, но их действие осуществляется в молочной железе, матке, влагалище.
2) Строгая специфичность действия. Реакции органов и тканей на гормоны строго специфичны и не могут быть вызваны иными биологически активными веществами. Например, удаление гипофиза у молодого растущего организма приводит к остановке роста, что связано с выпадением действия гормона роста. Одновременно происходит атрофия щитовидной железы, гонад и надпочечников. Предотвратить задержку роста и атрофию названных желез после гипофизэктомии можно только пересадкой (трансплантацией) гипофиза, инъекциями суспензии гипофиза или очищенных тропных гормонов.
3) Высокая биологическая активность. Гормоны образуются эндокринными железами в малых количествах. При введении извне они эффективны также в очень небольших концентрациях. Ежедневная доза гормона надпочечников преднизолона, поддерживающая жизнь человека, у которого удалены оба надпочечника, составляет всего 10 мг.
Ежедневная потребность в гормонах. Ежедневная минимальная потребность в гормонах для взрослого здорового человека представлена в табл. 13.
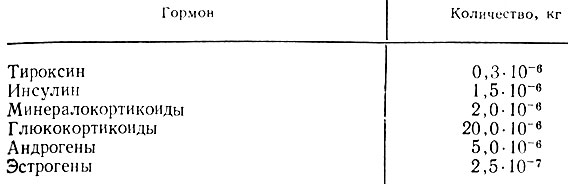
Таблица 13. Суточная потребность здоровых людей в гормонах
Действие гормонов на функции органов и систем организма опосредуется двумя основными механизмами. Гормоны могут оказывать свое влияние через нервную систему, а также гуморально, непосредственно воздействуя на активность органов, тканей и клеток.
Типы воздействия гормонов на организм. Физиологическое действие гормонов весьма разнообразно. Они оказывают выраженное влияние на обмен веществ, дифференциацию тканей и органов, рост и метаморфоз. Гормоны обладают способностью изменять интенсивность функций органов и организма в целом.
Механизм действия гормонов очень сложен. Основную свою функцию - влияние на обменные процессы, рост и половое созревание - они осуществляют в тесной связи с центральной нервной системой и воздействуя на ферментные системы организма.
Гормоны могут изменять интенсивность синтеза ферментов, активировать одни ферментативные системы и блокировать другие. Например, один из гормонов островков Лангерганса поджелудочной железы - глюкагон - активирует фермент печени фосфорилазу и усиливает тем самым переход гликогена в глюкозу. Одновременно он повышает активность содержащегося в печени фермента инсулиназы, которая разрушает избыток продуцируемого бета-клетками островков Лангерганса инсулина. В результате действия этих гормонов осуществляется регуляция углеводного обмена.
Наряду с непосредственным влиянием на ферментные системы тканей действие гормонов на строение и функции организма может осуществляться более сложными путями при участии нервной системы. Так, гормоны могут воздействовать на интерорецепторы, обладающие специфической чувствительностью к ним. Такие хеморецепторы расположены в стенках различных кровеносных сосудов. Вероятно, они имеются и в тканях.
Таким образом, гормоны, транспортируемые кровью по всему организму, могут действовать на эффекторные органы двумя путями: непосредственно, без участия нервного механизма, и через нервную систему. В последнем случае раздражение хеморецепторов служит началом рефлекторной реакции, которая изменяет функциональное состояние нервных центров.
Физиологическая роль желез внутренней секреции. 1) Гормоны участвуют в регуляции и интеграции функций организма. В сложно устроенных животных организмах имеются два механизма регуляции - нервный и эндокринный. Оба механизма тесно связаны между собой и осуществляют единую нейроэндокринную регуляцию. Вместе с тем нейроны различных уровней центральной нервной системы, включая ее высший отдел - кору головного мозга, участвуют в регуляции функций эндокринных желез. Железы внутренней секреции под влиянием нервных импульсов выделяют в кровь гормоны, в особенности в периоды, когда организм подвергается каким-либо неблагоприятным воздействиям или испытывает потребность в большем по сравнению с исходным количестве гормона.
Гормоны в отличие от нервных влияний реализуют действие медленно, поэтому биологические процессы, обусловленные ими, протекают также медленно. Эта особенность гормонов обеспечивает им существенную роль в регуляции формообразовательных явлений, развивающихся в широком временном интервале.
2) Гормоны адаптируют организм к изменяющимся условиям внутренней и внешней среды организма. Например, гипергликемия стимулирует секрецию инсулина поджелудочной железой, что приводит к восстановлению уровня глюкозы в крови.
3) Гормоны восстанавливают измененное равновесие внутренней среды организма. Например, при понижении уровня глюкозы в крови из мозгового слоя надпочечников выбрасывается большое количество адреналина, который усиливает гликогенолиз в печени, в результате чего нормализуется уровень глюкозы в крови.
Таким образом, основная роль гормонов в организме связана с их влиянием на морфогенез, обменные процессы и гомеостаз, т. е. с сохранением постоянства состава и свойств внутренней среды организма.
Регуляция образования гормонов. Продукция гормонов в железах внутренней секреции регулируется вегетативной нервной системой, промежуточным мозгом (гипоталамус) и корой головного мозга. Гормоны желез внутренней секреции в свою очередь оказывают сильное влияние на функции центральной нервной системы, особенно на состояние нейронов коры головного мозга. Следовательно, связь между эндокринными железами и центральной нервной системы двусторонняя.
В гормональной регуляции эндокринной деятельности большое значение имеет принцип авторегуляции. Например, тропные гормоны передней дозы гипофиза регулируют функции периферических эндокринных желез. При повышении же в крови уровня гормонов этих желез тормозится гормонообразовательная функция передней доли гипофиза. Принцип авторегуляции осуществляется и на основе сдвигов в химическом составе крови. Так, инсулин снижает содержание глюкозы в крови, что приводит к усиленному поступлению в сосудистое русло гормона-антагониста - адреналина, который путем мобилизации гликогена печени восстанавливает состав универсальной внутренней среды организма.
Судьба гормонов. Гормоны в процессе обмена изменяются функционально и структурно. Кроме того, часть гормонов утилизируется клетками организма, другая выводится с мочой. Гормоны подвергаются инактивации за счет соединения с белками, образования соединений с глюкуроновой кислотой, активности ферментов печени, процессов окисления.
Методы изучения функций желез внутренней секреции. Существуют клинические, анатомо-гистологические и экспериментальные методы исследования активности эндокринных желез.
Экспериментальные методы включают: экстирпацию (удаление), трансплантацию (пересадку) желез, экстирпацию с последующей трансплантацией удаленной железы, нагрузку организма животных гормонами, раздражение нервов или денервацию железы, метод условных рефлексов.
Во всех случаях ведут наблюдение за поведением животных, устанавливают и изучают измененные функции и обмен веществ в организме.
К современным методам исследования функций желез внутренней секреции относят следующие: 1) используют химические вещества (аллоксан) для повреждения бета-клеток островков Лангерганса и блокады ферментов (метилтиоурацил) щитовидной железы, участвующих в образовании гормонов; 2) применяют метод радиоактивных изотопов, например 131I, для изучения гормонообразовательной функции щитовидной железы; 3) широко используют биохимические методы определения содержания гормонов в крови, цереброспинальной жидкости, моче.
Функции желез внутренней секреции могут быть снижены (гипофункция) или повышены (гиперфункция).
Роль желез внутренней секреции в жизнепроявлениях организма животных и человека рассматривается в следующих разделах главы.
Гипофиз
В системе эндокринных желез гипофиз занимает особое положение. О гипофизе говорят как о центральной железе внутренней секреции. Это связано с тем, что гипофиз за счет своих специальных тропных гормонов регулирует деятельность других, так называемых периферических желез.
Гипофиз расположен в гипофизарной ямке турецкого седла клиновидной кости черепа. При помощи ножки он связан с основанием мозга.
Строение гипофиза. По своему строению гипофиз является сложным органом. Он состоит из аденогипофиза, который включает переднюю и среднюю доли, и нейрогипофиза, состоящего из задней доли. Аденогипофиз имеет эпителиальное происхождение, нейрогипофиз и его ножка - нейрогенное.
Гипофиз хорошо снабжается кровью. Особенностью кровообращения передней доли гипофиза является наличие портальной (воротной) системы сосудов, которые связывают ее с гипоталамусом. Установлено, что ток крови в воротной системе направляется от гипоталамуса к гипофизу (рис. 43).
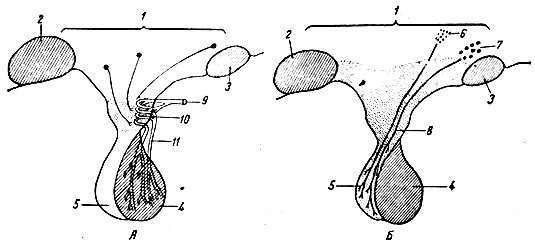
Рис. 43. Схема сосудистой связи гипоталамуса и передней доли гипофиза (А), а также нервной связи гипоталамуса и задней доли гипофиза (Б). 1 - промежуточный мозг; 2 - мамиллярные тела; 3 - перекрест зрительного нерва; 4 - передняя доля гипофиза; 5 - задняя доля гипофиза; 6 - паравентрикулярное ядро гипоталамуса; 7 - супраоптическое ядро; 8 - гипоталамо-гипофизарный тракт; 9 - артерия; 10 - первичная капиллярная сеть; 11 - гипоталамо-гипофизарная воротная сеть
Иннервация передней доли гипофиза представлена симпатическими и парасимпатическими нервными волокнами. Задняя доля гипофиза иннервируется нервными волокнами, берущими начало от нервных клеток супраоптического и паравентрикулярного ядер гипоталамуса.
Гормоны передней доли гипофиза. Гормоны, образующиеся в передней доле гипофиза, принято делить на две группы. К первой группе относятся гормон роста (соматотропин) и пролактин. Вторая группа включает тропные (кринотропные) гормоны: тиреотропный гормон (тиреотропин), адренокортикотропный гормон (кортикотропин) и гонадотропные гормоны (гонадотропины)*.
* (В скобках указаны названия гормонов, рекомендованные комиссией по биохимической номенклатуре Международного общества по чистой и прикладной химии и Международного биохимического общества.)
Гормон роста (соматотропин) участвует в регуляции роста, что обусловлено его способностью усиливать образование белка в организме. Наиболее выражено влияние гормона на костную и хрящевую ткань. Под влиянием соматотропина происходит усиленный рост эпифизарных хрящей в длинных костях верхних и нижних конечностей, что обусловливает увеличение их длины.
В зависимости от того, в какой период жизни отмечается нарушение соматотропной функции гипофиза, обнаруживают различные изменения роста и развития организма человека. Если активность передней доли гипофиза (гиперфункция) возникает в детском организме, то это приводит к усиленному росту тела в длину - гигантизм (рис. 44). При снижении функции передней доли гипофиза (гипофункция) в растущем организме происходит резкая задержка роста - карликовость (рис. 45). Избыточное образование гормона у взрослого человека не отражается на росте тела в целом, так как он уже завершен. Отмечается увеличение размеров тех частей тела, которые еще сохраняют способность к росту (пальцы рук и ног, кисти и стопы, нос и нижняя челюсть, язык, органы грудной и брюшной полости). Это заболевание получило название акромегалия (от греч. akros - конечность, megas - большой).
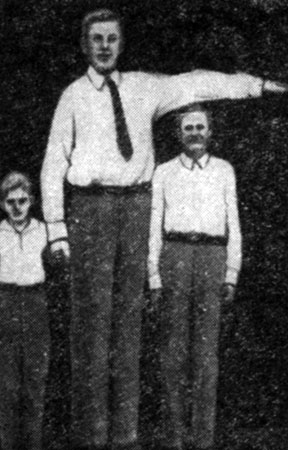
Рис. 44. Гигантизм. Справа - человек нормального роста, слева - подросток
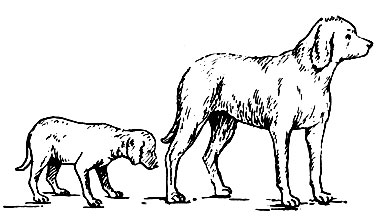
Рис. 45. Собаки одного помета. Возраст 12 мес. Слева - собака, которой в возрасте 21/2 мес удален гипофиз, справа - нормальная собака
Пролактин способствует образованию молока в альвеолах молочной железы. Свое действие на молочную железу пролактин оказывает после предварительного влияния на нее женских половых гормонов - эстрогенов и прогестерона. Эстрогены вызывают рост протоков молочной железы, прогестерон - развитие ее альвеол. После родов усиливается секреция гипофизом пролактина и наступает лактация. Важным фактором, способствующим секреции пролактина, является акт сосания, который через нервно-рефлекторный механизм стимулирует образование и выделение пролактина передней долей гипофиза.
Тиреотропный гормон (тиреотропин) избирательно действует на щитовидную железу, стимулируя ее функцию. Если удалить или разрушить гипофиз у животных, то наступает атрофия щитовидной железы. Введение тиреотропина, наоборот, вызывает разрастание ткани щитовидной железы, и происходит ее гипертрофия.
Под воздействием гормона наступают также и гистологические изменения в щитовидной железе, свидетельствующие о повышении ее активности: уменьшается количество коллоида в полостях фолликулов, происходит его вакуолизация, а затем и разжижение. Клетки фолликулов приобретают цилиндрическую форму. Тиреотропин активизирует протеолитические ферменты, под влиянием которых происходит расщепление тиреоглобулина и освобождение из него гормонов тироксина и трийодгиронина. Тиреотропин облагает также способностью стимулировать образование белка тиреоглобулина в клетках фолликулов щитовидной железы и поступление его в полость фолликула.
Адренокортикотропный гормон (кортикотропин) является физиологическим стимулятором пучковой и сетчатой зон коры надпочечников, которые образуют гормоны глюкокортикоиды.
Удаление гипофиза у животных приводит к атрофии коркового слоя надпочечников. Атрофические процессы захватывают все зоны коры, но наиболее глубокие изменения происходят в клетках сетчатой и пучковой зон.
Кортикотропин вызывает распад и тормозит синтез белка в организме. В этом отношении гормон является антагонистом соматотропина, который усиливает синтез белка. Кортикотропин, как и глюкокортикоиды, тормозит развитие основного вещества соединительной ткани, снижает проницаемость капилляров. Эти эффекты лежат в основе противовоспалительного действия гормона. Под влиянием адренокортикотропного гормона происходит уменьшение размера и массы лимфатических узлов, селезенки и особенно вилочковой железы, уменьшается количество лимфоцитов в периферической крови, возникает эозинопения.
К гонадотропинам относят три гормона: фолликулостимулирующий (фоллитропин), лютеинизирующий (лютропин) и лютеотропный гормон.
Фолликулостимулирующий гормон стимулирует рост везикулярного фолликула в яичнике, секрецию фолликулярной жидкости, формирование оболочек, окружающих фолликул. Влияние фоллитропина на образование женских половых гормонов - эстрогенов - небольшое. Этот гормон имеется как у женщин, так и у мужчин. У мужчин под влиянием фоллитропина происходит образование половых клеток - сперматозоидов.
Лютеинизирующий гормон необходим для роста везикулярного фолликула яичника на стадиях, предшествующих овуляции, и для самой овуляции. Без этого гормона не происходит овуляции и образования желтого тела на месте лопнувшего фолликула. Лютропин стимулирует образование эстрогенов. Однако, для того чтобы этот гормон осуществил свое действие на яичник (рост фолликулов, овуляция, секреция эстрогенов), необходимо длительное воздействие лютропина на везикулярные фолликулы.
Под воздействием лютеинизирующего гормона происходит также образование желтого тела из лопнувшего фолликула. Лютропин имеется как у женщин, так и у мужчин. У мужчин этот гормон способствует образованию мужских половых гормонов - андрогенов.
Лютеотропный гормон способствует функционированию желтого тела и образованию им гормона прогестерона.
Гормон средней доли гипофиза. В средней доле гипофиза образуется гормон меланотропин, или интермедии, который оказывает влияние на пигментный обмен. Если у лягушки разрушить гипофиз, то спустя некоторое время после этого изменяется цвет кожи лягушки - становится более светлым.
Гормоны задней доли гипофиза. Задняя доля гипофиза тесно связана с супраоптическим и паравентрикулярным ядрами гипоталамической области. Клетки этих ядер способны к нейросекреции. Образовавшийся нейросекрет транспортируется по аксонам нейронов этих ядер (по так называемому гипоталамо-гипофизарному тракту) в заднюю долю гипофиза. Установлено, что в нервных клетках паравентрикулярного ядра образуется гормон окситоцин, а в нейронах супраоптического ядра - вазопрессин. Накапливаются гормоны в клетках задней доли гипофиза - питуицитах. Однако питуициты нейрогипофиза не пассивные депо гормонов: в этих клетках гормоны превращаются в активную форму.
Вазопрессин выполняет в организме две функции. Первая связана с влиянием гормона на гладкую мускулатуру артериол, тонус которых он увеличивает, что приводит к повышению величины артериального давления. Вторая и основная функция связана с антидиуретическим действием вазопрессина. Антидиуретический эффект вазопрессина выражается в его способности усиливать обратное всасывание воды из канальцев почек в кровь. По мнению советского физиолога А. Г. Генецинского, это связано с тем, что вазопрессин повышает активность фермента гиалуронидазы, которая усиливает распад уплотняющего вещества в канальцах почек - гиалуроновой кислоты. В результате этого канальцы почек теряют водонепроницаемость и вода всасывается в кровь.
Уменьшение образования вазопрессина является причиной несахарного диабета (несахарное мочеизнурение). При этом заболевании выделяется большое количество мочи (иногда десятки литров в сутки), в которой не содержится сахара (в отличие от сахарного диабета). Одновременно у таких больных возникает сильная жажда.
Окситоцин избирательно действует на гладкую мускулатуру матки, усиливая ее сокращение. Сокращение матки резко усиливается, если она предварительно находилась под воздействием эстрогенов. Во время беременности окситоцин не влияет на матку, так как под действием гормона желтого тела прогестерона она становится нечувствительной ко всем раздражениям.
Окситоцин стимулирует также выделение молока. Под влиянием окситоцина усиливается именно выделение молока, а не его секреция, которая находится под контролем гормона передней доли гипофиза пролактина. Акт сосания рефлекторно стимулирует выделение окситоцина из нейрогипофиза.
Регуляция образования гормонов гипофиза. Регуляция образования гормонов гипофиза достаточно сложна и осуществляется несколькими механизмами.
Гипоталамическая регуляция. Доказано, что нейроны гипоталамуса обладают способностью вырабатывать нейросекрет, который содержит в своем составе соединения белковой природы. Эти вещества по сосудам, соединяющим гипоталамус и аденогипофиз, поступают в аденогипофиз, где оказывают свое специфическое действие, стимулируя или угнетая образование гормонов передней и средней долей гипофиза.
Регуляция образования гормонов в передней доле гипофиза осуществляется по принципу обратной связи. Между передней долей гипофиза и периферическими железами внутренней секреции существуют двусторонние отношения: кринотропные гормоны передней доли гипофиза активируют деятельность периферических эндокринных желез, которые в зависимости от их функционального состояния влияют на продукцию тропных гормонов передней доли гипофиза. Так, если в крови снижается уровень тироксина, то при этом происходит усиленное образование в передней доле гипофиза тиреотропного гормона. Наоборот, при избыточной концентрации тироксина в крови он тормозит образование в гипофизе тиреотропного гормона. Двусторонние взаимоотношения имеются между гипофизом и гонадами, гипофизом и щитовидной железой, гипофизом и корой надпочечников. Такое взаимоотношение получило название плюс-минус-взаимодействие. Тропные гормоны передней доли гипофиза стимулируют (плюс) функцию периферических желез, а гормоны периферических желез подавляют (минус) продукцию и выделение гормонов передней доли гипофиза.
В последнее время установлено, что существует обратная связь между гипоталамусом и тропными гормонами передней доли гипофиза. Например, гипоталамус стимулирует секрецию в передней доле гипофиза тиреотропина. Повышение концентрации в крови этого гормона приводит к торможению секреторной активности нейронов гипоталамуса, принимающих участие в освобождении тиреотропина в гипофизе.
На образование гормонов в передней доле гипофиза выраженное влияние оказывает вегетативная нервная система: симпатический ее отдел усиливает выработку кринотропных гормонов, парасимпатический угнетает.
Эпифиз (шишковидная железа)
Эпифиз - это образование конусовидной формы, которое нависает над верхними буграми четверохолмия. По внешнему виду железа напоминает еловую шишку, что и дало повод к ее названию.
Шишковидная железа состоит из паренхимы и соединительнотканной стромы. В состав паренхимы входят крупные светлые клетки, которые называют пинеальными.
Кровоснабжение эпифиза осуществляется кровеносными сосудами мягкой мозговой оболочки. Иннервация железы изучена недостаточно, однако известно, что этот орган получает нервные волокна непосредственно из центральной нервной системы и симпатического отдела вегетативной нервной системы.
Физиологическая роль шишковидной железы. Из ткани эпифиза выделены два соединения - мелатонин и гломерулотропин. Мелатонин участвует в регуляции пигментного обмена - он обесцвечивает меланофоры, т. е. оказывает действие, противоположное действию гормона средней доли гипофиза интермедина. Гломерулотропин участвует в стимуляции секреции гормона альдостерона корковым слоем надпочечников. Однако этот эффект гломерулотропина признают не все.
Щитовидная железа
Щитовидная железа состоит из двух долей, расположенных на шее по обеим сторонам трахеи ниже щитовидного хряща (рис. 46).
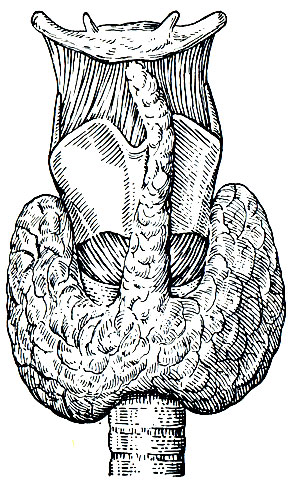
Рис. 46. Щитовидная железа человека
Щитовидная железа хорошо снабжается кровью и по кровоснабжению занимает одно из первых мест в организме. Иннервируется железа сетью нервных волокон, идущих к ней из нескольких источников: от среднего шейного симпатического узла, блуждающих, языкоглоточного и подъязычного нервов.
Щитовидная железа имеет дольчатое строение. Ткань каждой доли железы состоит из множества замкнутых железистых пузырьков, называемых фолликулами. Стенка каждого фолликула образована одним слоем эпителиальных клеток, форма которых в зависимости от функционального состояния щитовидной железы изменяется от кубической до призматической. Полость фолликула заполнена однородной вязкой массой желтоватого цвета, называемой коллоидом. Количество коллоида и его консистенция зависят от фазы секреторной деятельности и могут отличаться в разных фолликулах одной железы. В коллоиде щитовидной железы находится йодсодержащий белок тиреоглобулин.
Гормоны щитовидной железы. В щитовидной железе вырабатываются йодированные гормоны - тироксин (тетрайодтиронин) и трийодтиронин. Содержание тироксина в крови выше, чем трийодтиронина. Однако активность трийодтиронина в 4-10 раз выше, чем тироксина. В настоящее время известно, что в организме человека и животных имеется специальный гормон - тирокальцитонин, который участвует в регуляции кальциевого обмена. Основным источником этого гормона в организме млекопитающих служит щитовидная железа. Тирокальцитонин образуется парафолликулярными клетками щитовидной железы, которые расположены вне железистых ее фолликулов. Под влиянием тирокальцитонина снижается уровень кальция в крови. Гормон тормозит выведение кальция из костной ткани и увеличивает его отложение в ней. Тирокальцитонин угнетает функцию остеокластов, разрушающих костную ткань, и активирует функцию остеобластов, участвующих в образовании новой костной ткани.
Транспорт гормонов щитовидной железы. Основным гормоном щитовидной железы, циркулирующим в крови, является тироксин. Кроме тироксина, в крови присутствуют незначительные количества трийодтиронина. Оба гормона находятся в крови не в свободном виде, а в соединении с белками глобулиновой фракции.
При поступлении тироксина в кровоток он захватывается, в частности, клетками печени, где образует парные соединения с глюкуроновой кислотой, которые не обладают гормональной активностью и выводятся желчью в желудочно-кишечный тракт. Образование парных соединений тироксина с глюкуроновой кислотой рассматривают как путь инактивации гормона, благодаря которому предотвращается чрезмерное насыщение им крови.
Опыты с радиоактивным 131I показали, что в организме взрослого человека ежесуточно полностью разрушается в среднем около 300 мкг тироксина и трийодтиронина.
Регуляция образования гормонов щитовидной железы. Гормон передней доли гипофиза тиреотропин влияет на все стадии образования йодированных гормонов в щитовидной железе. При удалении у животных гипофиза интенсивность образования гормонов в щитовидной железе резко уменьшается.
Между тиреотропным гормоном гипофиза и гормонами щитовидной железы существуют взаимоотношения по типу прямых и обратных связей: тиреотропин стимулирует образование гормонов в щитовидной железе, а избыток тиреоидных гормонов в крови тормозит продукцию тиреотропного гормона в передней доле гипофиза.
Установлена зависимость между содержанием йода и гормонообразовательной активностью щитовидной железы. Малые дозы йода стимулируют, а большие тормозят процессы гормонопоэза.
Важную роль в регуляции образования гормонов в щитовидной железе выполняет вегетативная нервная система. Возбуждение ее симпатического отдела приводит к повышению, а преобладание парасимпатического тонуса обусловливает снижение гормонообразовательной функции этой железы.
Гипоталамическая область также оказывает выраженное влияние на образование гормонов в щитовидной железе. В нейронах гипоталамуса образуются вещества, которые, поступая в переднюю долю гипофиза, стимулируют синтез тиреотропина. При недостатке в крови гормонов щитовидной железы происходит усиленное образование этих веществ в гипоталамусе, а при избыточном содержании - торможение их синтеза, что в свою очередь уменьшает продукцию тиреотропина в передней доле гипофиза.
На функцию щитовидной железы оказывает влияние и ретикулярная формация ствола мозга. Показано, что при возбуждении нейронов ретикулярной формации происходит повышение функциональной активности щитовидной железы.
Кора головного мозга также участвует в регуляции активности щитовидной железы. Так, установлено, что в первый период после удаления коры головного мозга у животных отмечается повышение активности щитовидной железы, но в дальнейшем функция железы значительно снижается.
Физиологическая роль гормонов щитовидной железы. Йодсодержащие гормоны оказывают выраженное влияние на функции центральной нервной системы, высшую нервную деятельность, на рост и развитие организма, на все виды обмена веществ.
1) Влияние на функции центральной нервной системы. Длительное введение собакам больших доз тироксина приведет к повышению возбудимости, усилению сухожильных рефлексов, дрожанию конечностей. Удаление щитовидной железы у животных резко снижает их двигательную активность, ослабляет оборонительные реакции. Введение тироксина повышает двигательную активность собак и восстанавливает безусловные рефлексы, ослабленные или исчезнувшие после тиреоидэктомии.
2) Влияние на высшую нервную деятельность. У собак после удаления щитовидной железы условные рефлексы и дифференцировочное торможение вырабатываются с большим трудом. Сформированный условный рефлекс оказывается на следующий день утраченным, и его приходится снова вырабатывать. Введение тироксина усиливает процесс возбуждения в коре больших полушарий, что приводит в нормализации условнорефлекторной активности животных.
3) Влияние на процессы роста и развития. У амфибий тироксин стимулирует метаморфоз. Если у головастиков удалить зачаток щитовидной железы, то они утрачивают способность превращаться в лягушек.
Удаление щитовидной железы в молодом возрасте вызывает задержку роста тела млекопитающих (рис. 47). Нарушается развитие скелета. Центры окостенения появляются поздно. Животные становятся карликами. Замедляется развитие почти всех органов, половых желез.
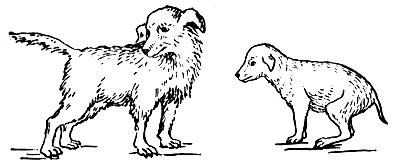
Рис. 47. Собаки одного помета. Справа собака, которой вскоре после рождения удалена щитовидная железа; слева - нормальная собака
4) Влияние на обмен веществ. Тироксин воздействует на обмен белков, жиров, углеводов и минеральный обмен. Гормон усиливает расходование всех видов питательных веществ, повышает потребление тканями глюкозы. Под влиянием тироксина в организме заметно уменьшается запас жира в депо и гликогена в печени.
Многообразное действие йодированных гормонов на обмен веществ связано с их влиянием на внутриклеточные процессы окисления и синтеза белка. Усиление энергетических и окислительных процессов под влиянием тиреоидных гормонов является причиной исхудания, обычно возникающего при гипертиреозе.
При введении животным гормонов щитовидной железы происходит значительное усиление основного обмена веществ. Так, если ввести собаке 1 мг тироксина, то суточный расход энергии увеличивается примерно на 1000 ккал.
5) Влияние на вегетативные функции организма. Тироксин увеличивает частоту сердечных сокращений, дыхательных движений, повышает потоотделение. Гормон снижает способность крови к свертыванию и повышает ее фибринолитическую способность. Это связано с тем, что гормон уменьшает образование в печени, почках, легких и сердце факторов, участвующих в процессе свертывания крови, и увеличивает синтез антикоагулянтов, а также веществ, стимулирующих фибринолитические свойства крови.
Нарушение функции щитовидной железы может сопровождаться или повышением, или снижением ее гормонообразовательной активности.
Если недостаточность функции щитовидной железы (гипотиреоз) проявляется у человека в детском возрасте, то возникает кретинизм (рис. 48). При этом заболевании наблюдаются нарушение пропорций тела, задержка роста, психического и полового развития. Для внешнего облика кретина характерны постоянно открытый рот и высунутый язык.

Рис. 48. Кретинизм
При недостаточной функциональной активности щитовидной железы может возникнуть и другое патологическое состояние, которое получило название микседемы (слизистый отек). Заболевание встречается преимущественно в детском и старческом возрасте, а также у женщин в климактерическом периоде.
У больных микседемой отмечаются психическая заторможенность, вялость, сонливость, снижение интеллекта и возбудимости симпатического отдела вегетативной нервной системы, нарушение половых функций. Наблюдается угнетение интенсивности всех видов обмена веществ. Основной обмен веществ снижен на 30-40%. Масса тела увеличена за счет повышения количества тканевой жидкости. У больных отмечается одутловатость лица.
При повышении функциональной активности щитовидной железы (гипертиреоз) возникает заболевание - тиреотоксикоз (базедова болезнь) (рис. 49). Характерными признаками этого заболевания являются увеличение щитовидной железы, пучеглазие, учащение сердечных сокращений, повышение обмена веществ, особенно основного, и температуры тела, увеличенное потребление пищи и вместе с тем исхудание. Значительные сдвиги отмечаются в активности нервной и мышечной систем. Наблюдается повышенная возбудимость и раздражительность, изменяются соотношения тонуса отделов вегетативной нервной системы, преобладает возбуждение симпатической нервной системы. Сухожильные рефлексы усилены, иногда отмечается мышечное дрожание. У больных обнаруживается мышечная слабость и быстрая утомленность.

Рис. 49. Тиреотоксикоз
Паращитовидные железы
Паращитовидные железы - парный орган. У человека имеются две пары околощитовидных желез, расположенных на поверхности или погруженных внутрь щитовидной железы.
Паращитовидные железы хорошо снабжаются кровью. Они имеют как симпатическую (от шейных ганглиев), так и парасимпатическую (блуждающий нерв) иннервацию.
Гормон паращитовидных желез. Паращитовидные железы вырабатывают паратгормон, образование которого происходит в главных и оксифильных клетках этих желез. Из паращитовидных желез гормон поступает непосредственно в кровь.
Паратгормон регулирует обмен кальция в организме и поддерживает постоянство его уровня в крови. В норме содержание кальция в крови у человека составляет 2,25-2,75 ммоль/л (9-11 мг%). При недостаточности паращитовидных желез (гипопаратиреоз) происходит значительное снижение уровня кальция в крови. Наоборот, при усилении деятельности паращитовидных желез (гиперпаратиреоз) наблюдается повышение концентрации кальция в крови.
Известно, что костная ткань скелета является главным депо кальция в организме, поэтому имеется определенная зависимость между уровнем кальция в крови и содержанием его в костной ткани. Паратгормон регулирует процессы обызвествления и декальцификации в костях. Влияя на обмен кальция, гормон одновременно воздействует на обмен фосфора в организме.
Считают, что паратгормон ослабляет обратное всасывание и усиливает выведение фосфатов с мочой. При усиленном образовании гормона наблюдается потеря фосфатов за счет их мобилизации из костной ткани. Освободившийся из соединений кальций начинает накапливаться в крови в увеличенных количествах. Таким образом, гиперкальциемия является одним из показателей повышенной функции паращитовидных желез.
После удаления паращитовидных желез в крови снижается уровень кальция и возрастает содержание фосфатов. Следовательно, между концентрацией кальция и фосфатов в крови существуют обратные соотношения.
Удаление паращитовидных желез у животных или их недостаточная функция у человека приводит к развитию вялости, потере аппетита, рвоте, фибриллярным подергиваниям мышц, спастическим конвульсиям, переходящим в тетанию. Фибриллярные подергивания одиночных мышц переходят в интенсивные спастические сокращения групп мышц, преимущественно конечностей, лица и затылка. Спазм гортани, паралич дыхательных мышц и остановка сердца приводят к смерти.
Регуляция деятельности паращитовидных желез. Активность этих желез определяется уровнем кальция в крови. Между гормонообразовательной функцией паращитовидных желез и уровнем кальция существует обратная зависимость. Если в крови концентрация кальция нарастает, то это приводит к снижению функциональной активности паращитовидных желез. При уменьшении же уровня кальция в крови происходит повышение гормонообразовательной функции паращитовидных желез.
Вилочковая железа (тимус)
Вилочковая железа - парный дольчатый орган, расположенный в верхнем отделе переднего средостения. Она состоит из двух долей неодинаковой величины, соединенных между собой прослойкой соединительной ткани. Каждая доля вилочковой железы включает в себя небольшие дольки, в которых различают корковый и мозговой слой. Корковое вещество представлено паренхимой, в которой имеется большое количество лимфоцитов. В мозговом веществе содержатся эпителиальные и липоидные клетки.
Вилочковая железа хорошо снабжается кровью. Иннервация железы осуществляется парасимпатическими (блуждающими) и симпатическими нервами, берущими начало от нижнего шейного и верхнего грудного симпатических ганглиев.
Физиологическая роль вилочковой железы. Эндокринная функция вилочковой железы до сих пор еще окончательно не выяснена. Попытки получить гормон этой железы пока не увенчались успехом.
Считают, что вилочковая железа играет большую роль в регуляции иммунных процессов организма, стимулируя образование антител, которые обеспечивают реакцию на чужеродный белок. Вилочковая железа контролирует развитие и распределение лимфоцитов, участвующих в иммунных реакциях.
Показано, что недифференцированные стволовые клетки, которые образуются в костном мозге, выходят в кровоток и поступают в вилочковую железу. В ней они размножаются и дифференцируются в лимфоциты тимусного происхождения (Т-лимфоциты). Полагают, что именно эти лимфоциты ответственны за развитие клеточного иммунитета. Т-лимфоциты составляют большую часть циркулирующих в крови лимфоцитов.
Вилочковая железа достигает максимального развития в детском возрасте. После наступления полового созревания развитие ее прекращается и железа начинает атрофироваться. В связи с этим полагают, что она стимулирует рост организма и тормозит развитие половой системы. Высказываются предположения, что вилочковая железа влияет на обмен кальция и нуклеиновый обмен.
Физиологическое значение вилочковой железы состоит также в том, что она содержит большое количество витамина С, уступая в этом отношении только надпочечникам.
При увеличении вилочковой железы у детей возникает тимико-лимфатический статус. Считают, что указанное состояние является врожденной конституциональной особенностью организма. При этом статусе, кроме увеличения вилочковой железы, происходит разрастание лимфатической ткани. Характерен внешний вид больного: пастозное одутловатое лицо, рыхлость подкожной клетчатки, тучность, тонкая кожа, мягкие волосы.
Поджелудочная железа
Поджелудочная железа относится к железам со смешанной функцией. Ацинозная ткань этой железы вырабатывает поджелудочный сок, который через выводной проток выделяется в полость двенадцатиперстной кишки. Внутрисекреторная деятельность поджелудочной железы проявляется в ее способности образовывать гормоны, которые поступают из железы непосредственно в кровь.
Морфологическим субстратом эндокринной функции поджелудочной железы служат островки Лангерганса, разбросанные среди ее железистой ткани. Островки неравномерно расположены по всей железе: преимущественно в ее хвостовой части и только небольшое количество в головном отделе железы.
Островки Лангерганса состоят из клеток трех типов: альфа-, бета- и гамма-клеток. Основную массу островков Лангерганса составляют бета-клетки. Около Vs общего количества клеток приходится на долю альфа-клеток, которые по своим размерам крупнее бета-клеток и расположены преимущественно по периферии железы. Показано, что у человека на 1 г железы приходится от 2700 до 25250 островков Лангерганса.
Поджелудочная железа иннервируется симпатическими нервами, идущими из солнечного сплетения, и веточками блуждающего, нерва. Однако иннервация ацинозной ткани и клеток островков Лангерганса осуществляется совершенно обособленно. Нервные волокна, иннервирующие островки Лангерганса, не соединяются с нервами экзокринного железистого аппарата поджелудочной железы. Каждый островок содержит значительное количество ганглиозных клеток, принадлежащих к вегетативной нервной системе.
Гистохимически установлено, что в островковой ткани железы содержится большое количество цинка. Цинк является также составной частью инсулина. Железа имеет обильное кровоснабжение.
Гормоны поджелудочной железы. Показано, что бета-клетки островков Лангерганса образуют гормон инсулин, альфа-клетки синтезируют глюкагон. В эпителии мелких выводных протоков происходит образование липокаической субстанции, которую одни исследователи относят к панкреатическим гормонам, другие рассматривают как вещество энзиматической природы.
Физиологическое значение инсулина. Инсулин участвует в регуляции углеводного обмена. Под действием гормона происходит уменьшение концентрации сахара в крови - возникает гипогликемия. Если уровень сахара в крови в норме составляет 4,45-6,65 ммоль/л (80-120 мг%), то под влиянием инсулина в зависимости от вводимой дозы он становится ниже 4,45 ммоль/л (80 мг%). Понижение уровня глюкозы в крови под влиянием инсулина связано с тем, что гормон способствует превращению глюкозы в гликоген в печени и мышцах. Кроме того, инсулин повышает проницаемость клеточных мембран для глюкозы. В связи с этим происходит усиленное проникновение глюкозы внутрь клетки, где осуществляется ее утилизация. Значение инсулина в регуляции углеводного обмена заключается также в том, что он предупреждает распад белков и превращает их в глюкозу. Инсулин также стимулирует синтез белка из аминокислот и их активный транспорт в клетки. Инсулин регулирует жировой обмен, способствуя образованию высших жирных кислот из продуктов углеводного обмена. Гормон тормозит мобилизацию жира из жировой ткани.
Активность инсулина выражают в лабораторных и клинических единицах. Лабораторная, или кроличья, единица - это количество гормона, которое у здорового кролика массой в 2 кг уменьшает содержание сахара в крови до 2,22 ммоль/л (40 мг%). За одну единицу действия (ЕД), или интернациональную единицу (ИЕ), принимают активность 0,04082 мг кристаллического инсулина. Клиническая единица составляет 1/3 лабораторной.
Регуляция секреции инсулина. В основе регуляции секреции инсулина лежит нормальное содержание глюкозы в крови. Гипергликемия приводит к увеличению поступления инсулина в кровь. Гипогликемия уменьшает образование и поступление гормона в сосудистое русло. Установлено, что паравентрикулярные ядра (высшие вегетативные центры парасимпатической нервной системы) гипоталамической области принимают непосредственное участие в регуляции образования и секреции инсулина поджелудочной железой. При увеличении концентрации сахара в крови происходит повышение активности нервных клеток паравентрикулярного ядра. Возникшие в нейронах нервные импульсы передаются к дорсальным ядрам блуждающего нерва, расположенным в продолговатом мозге. От нервных клеток этих ядер возбуждение по волокнам блуждающего нерва распространяется до ганглиев, расположенных непосредственно в ткани поджелудочной железы. В дальнейшем по аксонам нервных клеток указанных ганглиев импульсы поступают к бета-клеткам островков Лангерганса, что приводит к усилению образования и секреции инсулина. Инсулин превращает глюкозу в гликоген, и уровень сахара в крови восстанавливается до нормальных величин. Если количество глюкозы станет ниже нормы и возникнет гипогликемия, то происходит торможение активности паравентрикулярных ядер гипоталамуса и как следствие этого возбуждает не только нейроны паравентрикулярных ядер, но и непосредственно рецепторный аппарат островков Лангерганса, что также вызывает увеличение секреции инсулина.
Подтверждением положения о том, что образование инсулина регулируется уровнем глюкозы в крови, являются опыты с пересадкой собакам нескольких поджелудочных желез. У собаки с четырьмя поджелудочными железами не уменьшилось содержание глюкозы в крови. Следовательно, четыре поджелудочные железы в организме собаки приспосабливали свою гормонообразовательную функцию к уровню глюкозы в крови и не вызывали гипогликемическое состояние.
Обнаружено, что функция островков Лангерганса зависит также от функциональных взаимосвязей между гипофизом и паравентрикулярными ядрами гипоталамуса. Гипофиз тормозит активность нейронов паравентрикулярных ядер, что приводит к уменьшению образования инсулина бета-клетками островков Лангерганса поджелудочной железы. Ослабление влияния гипофиза на паравентрикулярные ядра сопровождается стимуляцией секреции инсулина.
Секреция инсулина регулируется вегетативной нервной системой: возбуждение блуждающих нервов стимулирует образование и выделение гормона, а симпатические нервы тормозят эти процессы.
Секреция инсулина происходит и рефлекторно при раздражении рецепторов ряда рефлексогенных зон, Так, при гипергликемическом состоянии возбуждаются хеморецепторы каротидных синусов, в результате чего осуществляется рефлекторный выброс инсулина в кровоток и уровень сахара в крови нормализуется.
Количество инсулина в крови зависит от активности фермента инсулиназы, который разрушает гормон. Наибольшее количество фермента содержится в печени и скелетных мышцах. При однократном протекании крови через печень инсулиназой разрушается до 50% инсулина.
Недостаточность внутрисекреторной функции поджелудочной железы, сопровождающаяся уменьшением секреции инсулина, приводит к заболеванию, которое получило название сахарного диабета, или сахарного мочеизнурения. Основными проявлениями этого заболевания являются гипергликемия, глюкозурия (появление сахара в моче), полиурия (увеличенное до 10 л/сут, выделение мочи), полифагия (повышенный аппетит), полидипсия (повышенная жажда), возникающая вследствие потери воды и солей.
Нарастание сахара в крови у больных диабетом, количество которого может составлять 16,65-44,00 ммоль/л (300-800 мг%), является результатом ослабления гликогенеза в печени и мышцах, а также нарушения утилизации глюкозы клетками организма. У больных диабетом нарушается не только углеводный обмен, но и обмен белков и жиров.
Физиологическое значение глюкагона. Глюкагон участвует в регуляции углеводного обмена. По характеру своего действия на обмен углеводов он является антагонистом инсулина. Под влиянием глюкагона происходит расщепление гликогена в печени до глюкозы. В результате этого концентрация глюкозы в крови повышается. Кроме того, глюкагон стимулирует расщепление жира в жировой ткани.
Регуляция секреции глюкагона. На образование глюкагона в альфа-клетках островков Лангерганса оказывает влияние количество глюкозы в крови При повышении содержания глюкозы в крови происходит торможение секреции глюкагона, при понижении - увеличение уровня гормона. Значение концентрации глюкозы в крови в образовании глюкагона было показано в опытах с перфузией изолированной поджелудочной железы: если в перфузируемой жидкости количество глюкозы было повышено, то наблюдалось снижение выброса глюкагона из железы в оттекающую жидкость. На образование глюкагона в альфа-клетках оказывает влияние и передняя доля гипофиза. Установлено, что гормон роста - соматотропин - повышает активность альфа-клеток и они усиленно продуцируют глюкагон.
Физиологическое значение липокаина. Гормон способствует утилизации жиров за счет стимуляции образования липидов и окисления жирных кислот в печени. Липокаин предотвращает жировое перерождение печени у животных после удаления поджелудочной железы.
Надпочечники
Надпочечники являются парными железами. Они располагаются непосредственно над верхними полюсами почек. Железы окружены плотной соединительнотканной капсулой и погружены в жировую ткань. Пучки соединительнотканной капсулы проникают внутрь железы, переходят в перегородки, которые делят надпочечники на два слоя - корковый и мозговой. Корковый слой имеет мезодермальное происхождение, мозговой развивается из зачатка симпатического ганглия.
Корковый слой надпочечников состоит из трех зон - клубочковой, пучковой и сетчатой.
Клетки клубочковой зоны лежат непосредственно под капсулой, собраны в клубочки. В пучковой зоне клетки расположены в виде продольных столбиков или пучков. Свое название сетчатая зона получила вследствие сетчатого характера расположения ее клеток. Все три зоны коркового слоя надпочечников не только представляют, собой морфологически обособленные структурные образования, но и выполняют разные физиологические функции.
Мозговой слой надпочечников состоит из хромаффинной ткани, в которой имеется два вида хромаффинных клеток - образующих адреналин и норадреналин. В настоящее время считают, что мозговой слой надпочечников представляет собой видоизмененный симпатический ганглий.
Надпочечники обильно снабжаются кровью и иннервируются симпатическими и парасимпатическими нервами. Симпатическая иннервация осуществляется чревными нервами, а также нервными волокнами, идущими из солнечного сплетения. Парасимпатическая иннервация надпочечников представлена ветвями блуждающего нерва. Имеются данные о том, что в иннервации надпочечников принимают участие диафрагмальные нервы.
Надпочечники представляют собой эндокринный орган, который имеет жизненно важное значение. Удаление надпочечников приводит к смерти. Показано, что жизненно необходимым является корковый слой надпочечников.
Гормоны коркового слоя надпочечников делят на три группы: 1) глюкокортикоиды - гидрокортизон, кортизон и кортикостерон, 2) минералокортикоиды - альдостерон, дезоксикортикостерон; 3) половые гормоны - андрогены, эстрогены, прогестерон.
Образование гормонов происходит преимущественно в какой-то одной зоне коры надпочечников. Так, минералокортикоиды образуются в клетках клубочковой зоны, глюкокортикоиды - пучковой, половые гормоны - сетчатой.
По химическому строению гормоны коры надпочечников являются стероидами. Образование их происходит из холестерина. Для синтеза гормонов коры надпочечников необходима также аскорбиновая кислота.
Физиологическое значение глюкокортикоидов. Эти гормоны оказывают влияние на обмен углеводов, белков и жиров. Они усиливают процесс образования глюкозы из белков, повышают отложение гликогена в печени. Глюкокортикоиды являются антагонистами инсулина в регуляции углеводного обмена: задерживают утилизацию глюкозы в тканях, а при передозировке могут привести к повышению концентрации сахара в крови и появлению его в моче.
Глюкокортикоиды оказывают катаболическое влияние на белковый обмен, вызывая распад тканевого белка и задерживая включение аминокислот в белки. Поскольку размножение и рост клеток организма не может происходить без синтеза белка, глюкокортикоиды задерживают формирование грануляций и последующее образование рубца, что отрицательно сказывается на заживлении ран.
Глюкокортикоиды являются противовоспалительными гормонами, так как обладают способностью угнетать развитие воспалительных процессов, в частности за счет понижения проницаемости мембран сосудов и уменьшения активности фермента гиалуронидазы.
Глюкокортикоиды подавляют синтез антител и тормозят реакцию взаимодействия чужеродного белка (антигена) с антителом.
Глюкокортикоиды оказывают выраженное влияние на кроветворные органы. Введение глюкокортикоидов в организм приводит к обратному развитию вилочковой железы и лимфоидной ткани, что сопровождается понижением количества лимфоцитов в периферической крови, а также уменьшением содержания эозинофилов.
Выведение глюкокортикоидов из организма осуществляется двумя путями: 75-90% поступивших в кровь гормонов удаляется с мочой, 10-25% - с калом и желчью.
Физиологическое значение минералокортикоидов. Эти гормоны участвуют в регуляции минерального обмена. В частности, альдостерон усиливает обратное всасывание ионов натрия в почечных канальцах и уменьшает обратное всасывание ионов калия. В результате этого понижается выделение натрия с мочой и увеличивается выделение калия, что приводит к повышению концентрации ионов натрия в крови и тканевой жидкости и увеличению в них осмотического давления. Возрастание осмотического давления во внутренней среде организма сопровождается задержкой воды и способствует повышению артериального давления.
Минералокортикоиды способствуют развитию воспалительных реакций. Провоспалительное действие этих гормонов связано с их способностью повышать проницаемость капилляров и серозных оболочек.
Минералокортикоиды участвуют в регуляции тонуса кровеносных сосудов. Альдостерон обладает способностью увеличивать тонус гладких мышц сосудистой стенки, повышая тем самым артериальное давление. При недостатке минералокортикоидов, обусловленном снижением функции коры надпочечников, наблюдается гипотония.
Суточная секреция минералокортикоидов составляет примерно 0,14 мг. Выводятся гормоны из организма с мочой (ежедневно 12-14 мкг).
Физиологическое значение половых гормонов коры надпочечников. Эти гормоны имеют большое значение в развитии половых органов в детском возрасте, т. е. тогда, когда внутрисекреторная функция половых желез еще слабо развита. Половые гормоны коры надпочечников обусловливают развитие вторичных половых признаков. Они оказывают также анаболическое действие на белковый обмен: синтез белка в организме усиливается за счет увеличенного включения в его молекулу аминокислот.
При недостаточной функции коры надпочечников развивается заболевание, получившее название "бронзовая болезнь", или болезнь Аддисона. Ранними признаками заболевания являются бронзовая окраска кожи, особенно на руках, шее, лице, повышенная утомляемость при физической и умственной работе, потеря аппетита, тошнота, рвота. Больной становится очень чувствительным к холоду и болевым раздражениям, более восприимчивым к инфекциям.
При повышенной функции коры надпочечников, что чаще всего связано с наличием в ней опухоли, не только увеличивается образование гормонов, но и отмечается преобладание синтеза половых гормонов над выработкой глюкокортикоидов и минералокортикоидов. В результате этого у таких больных начинают резко изменяться вторичные половые признаки. Например, у женщин могут появляться вторичные половые признаки мужчин: борода, грубый мужской голос, прекращение менструаций.
Регуляция образования глюкокортикоидов. Важную роль в регуляции образования глюкокортикоидов в коре надпочечников выполняет адренокортикотропный гормон (АКТГ) передней доли гипофиза. Влияние АКТГ на образование глюкокортикоидов в коре надпочечников осуществляется по принципу прямых и обратных связей: кортикотропин стимулирует продукция глюкокортикоидов, а избыточное содержание этих гормонов в крови приводит к торможению синтеза АКТГ в передней доле гипофиза.
Помимо гипофиза, в регуляции образования глюкокортикоидов участвует гипоталамус. Показано, что в ядрах переднего отдела гипоталамуса вырабатывается нейросекрет, который содержит фактор белковой природы, стимулирующий образование и освобождение кортикотропина. Этот фактор через общую систему кровообращения гипоталамуса и гипофиза попадает в переднюю его долю и способствует образованию АКТГ. Таким образом, в функциональном отношении гипоталамус, передняя доля гипофиза и кора надпочечников находятся в тесной связи, поэтому говорят о единой гипоталамо-гипофизарно-надпочечниковой системе.
Установлено, что под влиянием адреналина - гормона мозгового вещества - происходит усиленное образование глюкокортикоидов в коре надпочечников.
Регуляция образования минералокортикоидов. На образование минералокортикоидов оказывает влияние концентрация ионов натрия и калия в организме. Повышенное количество ионов натрия в крови и тканевой жидкости приводит к торможению секреции альдостерона в коре надпочечников, что обусловливает усиленное выделение натрия с мочой. Блокада образования минералокортикоидов происходит также при недостаточном содержании ионов калия в крови. При недостатке во внутренней среде организма ионов натрия продукция альдостерона увеличивается и как следствие этого повышается обратное всасывание этих ионов в почечных канальцах. Избыточная концентрация ионов калия в крови также стимулирует образование в коре надпочечников альдостерона. Таким образом, ионы натрия и калия оказывают на минералокортикоидную функцию коры надпочечников противоположное действие.
На образование минералокортикоидов оказывает влияние также количество тканевой жидкости и плазмы крови. Увеличение их объема приводит к торможению секреции альдостерона, что сопровождается усиленным выделением ионов натрия и связанной с ним воды.
Гормоны мозгового слоя надпочечников. Мозговой слой надпочечников вырабатывает катехоламины. Основной гормон мозгового слоя - адреналин. Вторым гормоном является предшественник адреналина в процессе его биосинтеза - норадреналин. В венозной крови, оттекающей от надпочечника, адреналин составляет до 80-90% общего количества катехоламинов.
Образование адреналина и норадреналина осуществляется хромаффинными клетками. Хромаффинныё клетки содержатся не только в мозговом веществе надпочечников, но и в других органах: аорте, у места разделения сонных артерий, среди клеток симпатических ганглиев малого таза, а также в отдельных ганглиях симпатической цепочки. Все эти клетки образуют так называемую адреналовую систему, в которой вырабатывается адреналин и близкие к нему физиологически активные вещества.
Физиологическое значение адреналина и норадреналина. Адреналин выполняет функции гормона, он поступает из надпочечников в кровь постоянно. При некоторых чрезвычайных состояниях организма (острое понижение артериального давления, кровопотеря, охлаждение организма, гипогликемия, повышенная мышечная деятельность, эмоции - боль, страх, ярость) увеличивается образование и выделение гормона в сосудистое русло.
Возбуждение симпатической нервной системы сопровождается увеличенным поступлением в кровь адреналина и норадреналина. Эти катехоламины усиливают и удлиняют эффекты симпатической нервной системы. На функции органов и активность физиологических систем адреналин оказывает такое же действие, как и симпатическая нервная система. Адреналин выраженно влияет на углеводный обмен, усиливая гликогенолиз в печени и в мышцах, в результате чего повышается содержание глюкозы в крови. При введении адреналина и усилении его продукции возникает гипергликемия и глюкозурия. Адреналин расслабляет бронхиальные мышцы, расширяя тем самым просвет бронхов и бронхиол. Он повышает возбудимость и сократимость сердечной мышцы, а также увеличивает частоту сердечных сокращений. Гормон повышает тонус сосудов, в связи с чем увеличивается артериальное давление. Однако на коронарные сосуды сердца, легких, головного мозга и работающих мышц адреналин оказывает не прессорное, а сосудорасширяющее действие.
Адреналин повышает работоспособность скелетных мышц. В этом проявляется его адаптационно-трофическое влияние на функции организма. Адреналин угнетает моторную функцию желудочно-кишечного тракта и повышает тонус его сфинктеров.
Адреналин относят к так называемым гормонам короткого действия. Это связано с тем, что в крови и тканях гормон быстро разрушается под воздействием фермента моноаминоксидазы до продуктов, которые не обладают гормональной активностью.
Норадреналин в отличие от адреналина выполняет функцию медиатора - передатчика возбуждения с нервных окончаний на эффектор. Норадреналин участвует также в передаче возбуждения в нейронах центральной нервной системы.
Регуляция образования гормонов мозгового слоя. Образование гормонов в мозговом слое надпочечников хромаффинными клетками регулируется нервной системой. М. Н. Чебоксаров (1910) впервые показал, что при раздражении чревных нервов, которые по своей функции являются симпатическими, наступает усиление, а при перерезке их - уменьшение выхода адреналина из надпочечников. Одновременно при раздражении чревного нерва в кровь из надпочечников поступает норадреналин.
Секреторная функция мозгового слоя надпочечников контролируется гипоталамической областью головного мозга, так как в задней группе его ядер расположены высшие вегетативные центры симпатической нервной системы. При раздражении нейронов гипоталамуса происходит выброс адреналина из надпочечников и увеличение его содержания в крови.
Кора головного мозга влияет на поступление адреналина в сосудистое русло, что доказывается методом условных рефлексов.
Выделение адреналина из мозгового слоя надпочечников может происходить рефлекторно, например, при мышечной работе, эмоциональном возбуждении, охлаждении организма и при других воздействиях на организм. Выделение адреналина из надпочечников регулируется уровнем сахара в крови. При гипогликемическом состоянии организма происходит рефлекторное выделение адреналина из хромаффинных клеток адреналовой системы.
Участие надпочечников в общем адаптационном синдроме организма. Гормоны коры надпочечников повышают сопротивляемость организма к воздействиям различных факторов (охлаждение, голодание, травма, гипоксия, химическая или бактериальная интоксикация и т. д.). При этом наступают однотипные, неспецифические изменения в организме, проявляющиеся в первую очередь быстрым выделением кортикостероидов, особенно глюкокортикоидов, под влиянием кортикотропина.
Наступающие в организме изменения в ответ на действие чрезвычайных (стрессорных) раздражителей получили название общего адаптационного синдрома. Этот термин принадлежит канадскому патологу и эндокринологу Селье, который в течение многих лет изучал сущность общего адаптационного синдрома и обусловливающие его механизмы.
В дальнейшем было показано, что в развитии общего адаптационного синдрома участвует и мозговой слой надпочечников.
Установлено, что симпатико-адреналовая система начинает реакцию, которая развивается в организме в условиях чрезвычайного напряжения, гормоны коры надпочечников поддерживают и продолжают эту реакцию, в результате которой повышается уровень работоспособности эффекторных клеток.
Селье описаны фазы общего адаптационного синдрома, сущность и значение которых освещается при изучении патологической физиологии.
Половые железы
Половые железы - семенники у мужчин и яичники у женщин - относятся к железам со смешанной функцией. За счет внешнесекреторной функции этих желез образуются мужские и женские половые клетки - сперматозоиды и яйцеклетки. Внутрисекреторная функция проявляется в выработке мужских и женских половых гормонов, которые поступают в кровь.
Половые железы имеют хорошо выраженную сосудистую систему, за счет которой осуществляется их обильное кровоснабжение.
Иннервация половых желез обеспечивается постганглионарными симпатическими нервными волокнами, идущими из солнечного сплетения, и парасимпатическим тазовым нервом.
Развитие половых желез и поступление из них в кровь половых гормонов определяет половое развитие и созревание. Половая зрелость у человека наступает в возрасте 12-16 лет. Она характеризуется полным развитием первичных и появлением вторичных половых признаков.
К первичным половым признакам относятся половые железы (семенники, яичники) и половые органы (половой член, предстательная железа, влагалище, матка, яйцеводы). Они определяют возможность осуществления полового акта и деторождения.
Вторичные половые признаки - это те особенности половозрелого организма, по которым мужчина и женщина отличаются друг от друга. У мужчин вторичными половыми признаками являются растительность на лице, волосяной покров на теле, изменение голоса, формы тела, а также психики и поведения. У женщин к вторичным половым признакам относят особенности расположения волос на теле, изменение формы тела, развитие молочных желез.
Значение половых гормонов в развитии половых признаков отчетливо проявляется в опытах с удалением (кастрация) и пересадкой половых желез у петуха и курицы. Если удалить половые железы у этих птиц, то после кастрации они по виду начинают приближаться к среднему, бесполому типу (рис. 50). Пересадка им половых желез другого пола приводит к развитию внешних признаков и реакций, присущих противоположному полу: петух приобретает признаки и черты поведения, свойственные курице (феминизация), у курицы появляются качества, свойственные петуху (маскулинизация).

Рис. 50. Изменение пола. 1 - нормальный петух; 2 - нормальная курица; 3 - кастрированный петух; 4 - кастрированная курица; 5 - кастрированный петух, которому пересажены яичники курицы; 6 - кастрированная курица, которой пересажены семенники петуха
Мужские половые гормоны. Образование мужских половых гормонов происходит в особых клетках семенников - интерстициальных. Мужские половые гормоны называют андрогенами. В настоящее время установлено наличие в семенниках двух андрогенов - тестостерона и андростерона. Ежедневная потребность человека в андрогенах составляет около 5 мг. За сутки у мужчин выделяется с мочой 3-10 мкг андрогенов.
Гормоны стимулируют рост и развитие полового аппарата, мужских вторичных половых признаков и появление половых рефлексов. Если вводить андрогены неполовозрелым самцам, то у них преждевременно развиваются половые органы и вторичные половые признаки. Введение андрогенов кастратам-самцам приводит к устранению у них последствий кастрации.
Андрогены необходимы для нормального созревания мужских половых клеток - сперматозоидов. При отсутствии гормонов не образуются подвижные зрелые сперматозоиды. Кроме того, андрогены способствуют более длительному сохранению двигательной активности мужских половых клеток. Андрогены необходимы также для проявления полового инстинкта и осуществления связанных с ним поведенческих реакций.
Андрогены оказывают большое влияние на обмен веществ в организме. Они увеличивают образование белка в различных тканях, особенно в мышцах, уменьшают содержание жира в организме, повышают основной обмен веществ.
Андрогены оказывают влияние на функциональное состояние центральной нервной системы, на высшую нервную деятельность. После кастрации у самцов наступают резкие сдвиги в высшей нервной деятельности, нарушается процесс торможения в коре больших полушарий.
Женские половые гормоны. Образование женских половых гормонов - эстрогенов - происходит в фолликулах яичника. Фолликул представляет собой пузырек, стенка которого образована трехслойной оболочкой. Синтез эстрогенов осуществляется оболочкой фолликула. В желтом теле яичника, которое развивается на месте лопнувшего фолликула, вырабатывается гормон прогестерон. Ежедневная потребность организма женщины в эстрогенах составляет 0,25 мг. За сутки у женщины выделяется с мочой 16-36 мкг эстрогенов.
Эстрогены стимулируют рост яйцеводов, матки, влагалища, вызывают разрастание внутреннего слоя матки - эндометрия, способствуют развитию вторичных женских половых признаков и проявлению половых рефлексов. Кроме того, эстрогены вызывают усиление сокращений мышцы матки, повышают ее чувствительность к гормону задней доли гипофиза окситоцину. Они стимулируют также развитие и рост молочных желез. Прогестерон обеспечивает нормальное протекание беременности. Под его воздействием происходит разрастание слизистой оболочки эндометрия матки. Это создает благоприятные условия для имплантации оплодотворенной яйцеклетки в эндометрий матки. Прогестерон способствует также развитию вокруг имплантированной яйцеклетки так называемой децидуальной ткани. Прогестерон тормозит сокращение мускулатуры беременной матки и уменьшает ее чувствительность к окситоцину. Прогестерон задерживает созревание и овуляцию фолликулов за счет угнетения образования гормона передней доли гипофиза лютропина.
Регуляция образования гормонов половых желез. Образование половых гормонов в половых железах находится под контролем фолликулостимулирующего, лютеинизирующего и лютеотропного гормонов передней доли гипофиза.
У самок фолликулостимулирующий гормон способствует росту и развитию фолликулов, у самцов - созреванию половых клеток - сперматозоидов. Лютеинизирующий гормон обусловливает продукцию мужских и женских половых гормонов, а также овуляцию и образование на месте лопнувшего граафова пузырька желтого тела. Под влиянием лютеотропного гормона происходит синтез гормона желтого тела. Противоположное действие на функции половых желез оказывает гормон эпифиза мелатонин, который тормозит деятельность половых желез.
Функция половых желез регулируется нервной системой. Показано, что нервная система оказывает влияние на активность яичников и семенников рефлекторным путем за счет изменения образования в гипофизе гонадотропных гормонов.
Центральная нервная система участвует в регуляции нормального полового цикла. При изменении функционального состояния центральной нервной системы, например при сильных эмоциях (испуг, горе), может произойти нарушение полового цикла или даже его прекращение (эмоциональная аменорея).
Таким образом, регуляция гормонообразовательной функции половых желез осуществляется по общему принципу за счет нервных и гуморальных (гормональных) воздействий.
Понятие о тканевых гормонах. В настоящее время известно, что специализированные клетки различных органов и тканей вырабатывают биологически активные вещества. Эти вещества получили название тканевых гормонов. Тканевые гормоны оказывают многообразные влияния на регуляцию деятельности тех органов, где они образуются.
Большая группа тканевых гормонов синтезируется слизистой оболочкой желудочно-кишечного тракта. Эти гормоны влияют на образование и секрецию пищеварительных соков, а также моторную функцию желудочно-кишечного тракта.
В тканях образуются тканевые гормоны, которые участвуют в регуляции местного кровообращения (гистамин расширяет кровеносные сосуды, серотонин обладает прессорным эффектом).
К тканевым гормонам относят также компоненты кининовой системы организма - калликреин, под влиянием которого образуется сосудорасширяющий полипептид - брадикинин.
В последние годы значительную роль в местной регуляции физиологических функций отводят простагландинам - большой группе веществ, образующихся в микросомах всех тканей организма из ненасыщенных жирных кислот. Различные виды простагландинов участвуют в регуляции секреции пищеварительных соков, процесса агрегации тромбоцитов, изменении тонуса гладких мышц сосудов и бронхов.
К тканевым гормонам относят и медиаторы нервной системы - ацетилхолин и норадреналин.
При использовании материалов сайта активная ссылка обязательна:
http://anfiz.ru/ 'Анатомия и физиология человека'