Вегетативная часть нервной системы
(Данные по анатомии вегетативной нервной системы в настоящем издании дополнены исследованиями Д. М. Голуба, а также сведениями из монографии Митчелла (1953), Ф. А. Поемного и Е. П. Семеновой (1960). Более широко использованы данные В. П. Воробьева.)
Выше отмечалась коренная качественная разница в строении, развитии и функции гладкой и поперечнополосатой мускулатуры. Эти функциональные различия связаны с разницей в иннервации: скелетная мускулатура получает двигательные импульсы от анимальной, соматической части нервной системы, гладкая мускулатура - от вегетативной.
Вегетативная нервная система управляет деятельностью всех органов, участвующих в осуществлении растительных функций организма (питание, дыхание, выделение, размножение, циркуляция жидкостей), а также осуществляет трофическую иннервацию (И. П. Павлов).
Трофическая функция вегетативной нервной системы определяет питание тканей и органов применительно к выполняемой ими функции в тех или иных условиях внешней среды (адаптационно-трофическая функция).
Известно, что изменения в состоянии высшей нервной деятельности отражаются на функции внутренних органов и, наоборот, изменение внутренней среды организма оказывает влияние на функциональное состояние центральной нервной системы. Вегетативная нервная система усиливает или ослабляет функцию специфически работающих органов. Эта регуляция имеет тонический характер, поэтому вегетативная нервная система изменяет тонус органа. Так как одно и то же нервное волокно способно действовать лишь в одном направлении и не может одновременно повышать и понижать тонус, то сообразно с этим вегетативная нервная система распадается на два отдела или системы: симпатическую и парасимпатическую - pars sympathica et pars parasympathica.
Симпатический отдел по своим основным функциям является трофическим. Он осуществляет усиление окислительных процессов, потребление питательных веществ, усиление дыхания, учащение деятельности сердца, увеличение поступления кислорода к мышцам.
Роль парасимпатического отдела охраняющая: сужение зрачка при сильном свете, торможение сердечной деятельности, опорожнение полостных органов.
Сравнивая область распространения симпатической и парасимпатической иннервации, можно, во-первых, обнаружить преобладающее значение одного какого-либо вегетативного отдела. Мочевой пузырь, например, получает в основном парасимпатическую иннервацию, и перерезка симпатических нервов не изменяет существенно его функции; только симпатическую иннервацию получают потовые железы, волосковые мышцы кожи, селезенка, надпочечники. Во-вторых, в органах с двойной вегетативной иннервацией наблюдается взаимодействие симпатических и парасимпатических нервов в форме определенного антагонизма. Так, раздражение симпатических нервов вызывает расширение зрачка, сужение сосудов, ускорение сердечных сокращений, торможение перистальтики кишечника; раздражение парасимпатических нервов приводит к сужению зрачка, расширению сосудов, замедлению сердцебиения, усилению перистальтики.
Однако так называемый антагонизм симпатической и парасимпатической систем не следует понимать статически, как противопоставление функций этих двух отделов. Эти системы взаимодействующие, соотношение между ними динамически меняется на различных фазах функции того или иного органа; они могут действовать и антагонистически, и синергически.
Антагонизм и синергизм - две стороны единого процесса. Нормальные функции нашего организма обеспечиваются согласованным действием этих двух отделов вегетативной системы. Эта согласованность и регуляция функций осуществляется корой головного мозга.
Автономия деятельности вегетативной нервной системы не является абсолютной и проявляется лишь в местных реакциях коротких рефлекторных дуг. Поэтому предложенный PNA термин "автономная нервная система" не является точным, чем и объясняется сохранение старого, более правильного и логичного термина "вегетативная нервная система". Деление вегетативной нервной системы на симпатический и парасимпатический отделы проводится главным образом на основании физиологических и фармакологических данных, но имеются и морфологические отличия, обусловленные строением и развитием этих отделов нервной системы.
Поэтому охарактеризуем сначала морфологические особенности вегетативной нервной системы в сравнении с анимальной. Опишем прежде всего центры вегетативной нервной системы (рис. 339).
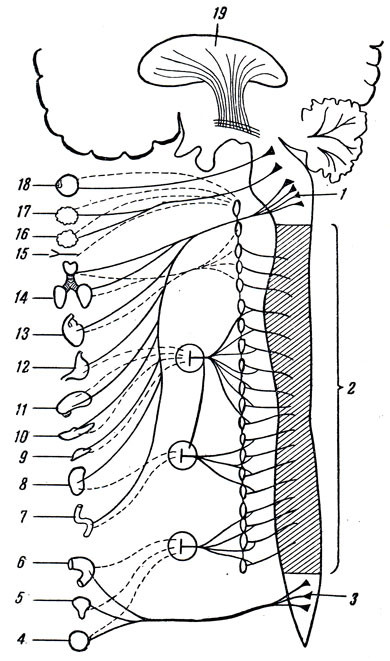
Рис. 339. Общая схема вегетативной нервной системы. Пунктиром обозначены постганглионарные волокна симпатической системы, идущие к органам; непрерывной линией - волокна парасимпатической и преганглионарные волокна симпатической системы. Область центров симпатической системы в спинном мозгу заштрихована. 1 - центры парасимпатической системы в головном мозгу (краниальный отдел); 2 - центры симпатической нервной системы; 3 - центры парасимпатической системы в нижнем конце спинного мозга (крестцовый отдел); 4 - половые органы; 5 - мочевой пузырь; 6 - толстая кишка; 7 - тонкая кишка; 8 - почка; 9 - надпочечник; 10 - поджелудочная железа; 11 - печень; 12 - желудок; 13 - сердце; 14 - легкие; 15 - сосуды головы; 16, 17 - слюнные железы; 18 - глаз; 19 - полосатое тело
Анимальные нервы выходят из мозгового ствола и спинного мозга на всем их протяжении сегментарно, причем эта сегментарность сохраняется частично и на периферии. Вегетативные нервы выходят только из нескольких отделов (очагов) центральной нервной системы. Имеются 4 таких очага, откуда выходят вегетативные нервы:
1. Мезенцефалический отдел - в среднем мозгу (nucl. accessorius или ядро Якубовича и непарное срединное ядро III пары головных нервов).
2. Бульварный - в продолговатом мозгу и мосте (ядра VII, IX и X пар головных нервов). Оба эти отдела объединяются под названием краниального.
3. Тораколюмбальный отдел - в боковых рогах спинного мозга на протяжении сегментов CVIII, ThI-LIII.
4. Сакральный отдел - в боковых рогах спинного мозга на протяжении сегментов SII-SIV.
Тораколюмбальный отдел относится к симпатической системе, а краниальный и сакральный - к парасимпатической (см. рис. 339, рис. 340).
Над этими очагами доминируют высшие вегетативные центры, которые не являются симпатическими или парасимпатическими, а объединяют в себе регуляцию обоих отделов вегетативной части нервной системы. Они являются надсегментарными и расположены в стволе и плаще мозга, а именно:
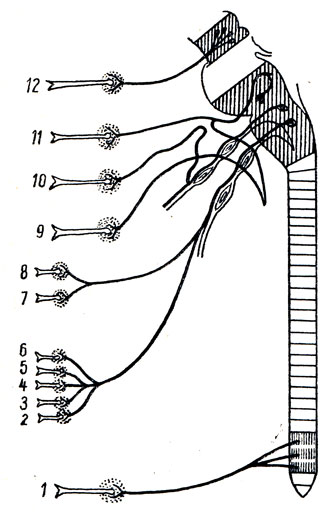
Рис. 340. Схема парасимпатической нервной системы. Преганглионарные волокна изображены сплошными линиями, постганглионарные - указателями. 1 - подчревное сплетение: парасимпатические преганглионарные волокна подходят в составе тазовых нервов, постганглионарные направляются к прямой кишке, мочевому пузырю, половым органам. Постганглионарные волокна: 2 - к почкам; 3 - к поджелудочной железе; 4 - к печени; 5 - к кишечнику; 6 - к желудку; 7 - к бронхам; 8 - к сердцу (2 - 8 - преганглионарные волокна подходят в составе блуждающего нерва); 9 - подчелюстной узел: преганглионарные волокна подходят по нерву, носящему название барабанной струны (ветвь лицевого нерва); постганглионарные направляются к подчелюстной и подъязычной слюнным железам; 10 - ушной узел: преганглионарные волокна подходят по ветви языкоглоточного нерва; постганглионарные направляются к околоушной железе; 11 - крылонебный узел: преганглионарные волокна подходят по большому каменистому нерву (ветвь лицевого нерва); постганглионарные направляются к слезной железе и железам полости рта и носа; 12 - ресничный узел: преганглионарные волокна подходят по глазодвигательному нерву, постганглионарные направляются к мышце, суживающей зрачок, и ресничной мышце глаза
1. Задний мозг: сосудодвигательный центр на дне IV желудочка (Ф. О. Овсянников); мозжечок, которому приписывают регуляцию ряда вегетативных функций (сосудодвигательные рефлексы, трофика кожи, скорость заживления ран и др.)
2. Средний мозг: серое вещество сильвиева водопровода.
3. Промежуточный мозг: hypothalamus (tuber cinereum).
4. Концевой мозг: полосатое тело.
Наибольшее значение для вегетативной регуляции имеет гипоталамическая область, которая является одним из самых древних отделов головного мозга, хотя и в ней различают более старые образования и филогенетически более молодые.
Ядра гипоталамической области связаны посредством гипоталамо-гипофизарного пучка с гипофизом, с которым образуют гипоталамо-гипофизарную систему. Эта система, действуя с помощью инкретов гипофиза, является регулятором всех эндокринных желез.
Гипоталамическая область регулирует деятельность всех органов растительной жизни, объединяя и координируя их функции.
Объединение вегетативных и анимальных функций всего организма осуществляется в коре большого мозга (В. Я. Данилевский, В. М. Бехтерев), особенно в премоторной зоне.
Кора, будучи, по И. П. Павлову, комплексом корковых концов анализаторов, получает раздражения от всех органов, в том числе и от органов растительной жизни, и через посредство своих эфферентных систем, в том числе и вегетативной нервной системы, оказывает влияние на эти органы. Следовательно, существует двусторонняя связь коры и внутренностей - кортико-висцеральная связь (К. М. Быков, В. Н. Черниговский и др.). Благодаря этому все вегетативные функции подчиняются коре головного мозга, которая ведает всеми процессами организма.
Таким образом, вегетативная нервная система есть не автономная система, как это считали до И. П. Павлова, а специализированная часть единой нервной системы, подчиненная высшим отделам ее, включая и кору большого мозга. Поэтому, как и в анимальной части нервной системы, в вегетативной части можно различать центральный и периферический ее отделы. К центральному отделу относятся описанные выше очаги и центры в спинном и головном мозгу, а к периферическому - нервные узлы, нервы, сплетения и периферические нервные окончания.
В последнее время появились сообщения (Н. Г. Колосов, А. А. Милохин) о том, что вегетативные узлы имеют свою афферентную иннервацию, благодаря которой они находятся под контролем центральной нервной системы.
Значительные отличия имеет рефлекторная дуга (рис. 341).
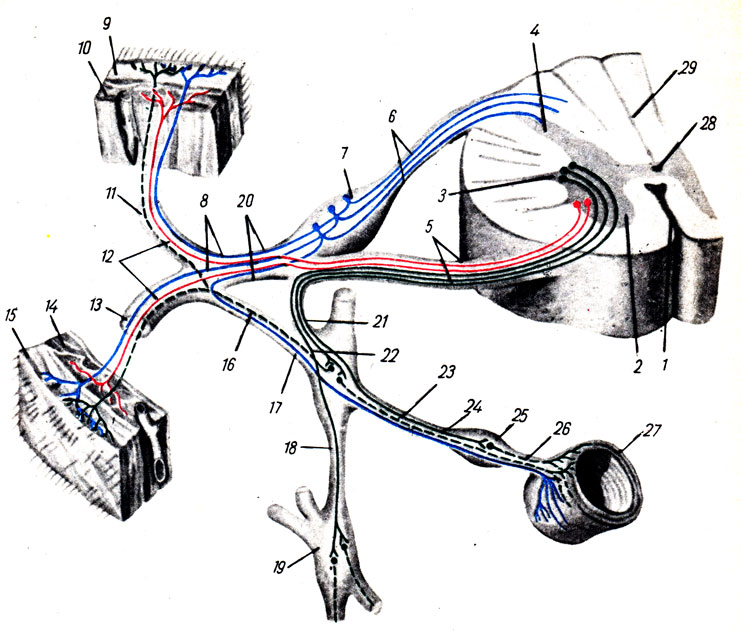
Рис. 341. Схема рефлекторной дуги симпатической нервной системы и анимальной (по Р. Д. Синельникову). 1 - fissura mediana ant.; 2 - cornu anterius; 3 - cornu laterale; 4 - cornu posterius; 5 - radix ventralis; 6 - radix dorsalis; 7 - ganglion spinale; 8 - афферентные волокна (соматосенсорные); 9, 15 - кожа; 10, 14 - мышца; 11 - r. dorsalis n. spinalis; 12 - двигательные волокна клеток переднего рога спинного мозга; 13 - r. ventralis n. spinalis; 16 - r. communicans griseus (librae postganglionares n. spinalis.); 17 - афферентные волокна (висцеросенсорные); 18 - r. interganglionaris tr. symp.; 19 - ganglion trunci symp; 20 - nervus spinalis; 21 - r. communicans albus (fibrae preganglionares к ganglion prevertebrale); 22 - r. communicans albus (fibrae preganglionares к ganglion trunci sympathici); 23 - fibrae postganglionares от ganglion trunci sympathici; 24 - fibrae preganglionares к ganglion prevertebrale; 25 - ganglion prevertebrale; 26 - fibrae postganglionares от ganglion prevertebrale; 27 - орган (кишка); 28 - canalis centralis; 29 - medulla spinalis
Клеточное тело воспринимающего нейрона как для анимальной, так и для вегетативной нервной системы помещается в спинальном узле, ganglion spinale, куда стекаются афферентные пути как от органов животной жизни, так и от органов растительной жизни и который, таким образом, является смешанным анимально-вегетативным узлом (А. Г. Кнорре, И. Д. Лев, 1963). Клеточное тело вставочного нейрона вегетативной части нервной системы в отличие от анимальной части нервной системы помещается в боковых рогах спинного мозга. При этом аксон вставочного анимального нейрона, исходящий из клеток заднего рога, заканчивается в пределах спинного мозга среди клеток его передних рогов. Что же касается вставочного нейрона вегетативной нервной системы, то он в спинном мозгу не заканчивается, а выходит за его пределы, к нервным узлам, расположенным на периферии. Выйдя из спинного мозга, аксон вставочного нейрона подходит или к узлам симпатического ствола, ganglia trunci sympathici, относящимся к симпатическому отделу вегетативной части нервной системы (это узлы как бы первого порядка; они образуют пограничный симпатический ствол), или волокна не заканчиваются в этих узлах, а направляются к промежуточным узлам, ganglia intermedia, расположенным более периферично, между симпатическим стволом и органом (ganglia sernilunaria, ganglia mesenterica). Это узлы второго порядка, они также относятся к симпатической системе. Наконец, волокна могут доходить, не прерываясь, до узлов, лежащих или около органа (околоорганные узлы, например ganglia ciliare, oticum и др.), или в толще органа (внутриорганные, интрамуральные узлы); и те и другие, являющиеся узлами третьего порядка, называют конечными узлами (ganglia teminalia). Они относятся к парасимпатическому отделу вегетативной части нервной системы*. Все волокна, идущие до узлов первого, второго или третьего порядка и являющиеся аксонами промежуточного нейрона, называются предузловыми волокнами, rami preganglionares. Они покрыты миелином.
* (Кроме макроскопически видимых обособленных узлов, по ходу вегетативных нервов встречаются мигрировавшие сюда в ходе эмбрионального развития небольшие группы эффекторных нейронов - микроганглии.)
Третий, эффекторный, нейрон анимальной рефлекторной дуги помещается в передних рогах спинного мозга, а эффекторный нейрон вегетативной рефлекторной дуги вынесен в процессе развития из центральной нервной системы в периферическую, ближе к рабочему органу, и располагается в вегетативных нервных узлах. Из такого расположения эффекторных нейронов на периферии вытекает главный признак вегетативной нервной системы - двухнейронность эфферентного периферического пути: первый нейрон - вставочный; тело его лежит в вегетативных ядрах головных нервов или боковых рогах спинного мозга, а нейрит идет к узлу; второй - эфферентный, тело которого лежит в узле, а нейрит достигает рабочего органа. Эффекторные нейроны симпатических нервов начинаются в ganglia trunci sympathici (узлах первого порядка) или ganglia intermedia (узлах второго порядка), а для парасимпатических нервов - в около- или внутриорганных узлах, ganglia terminalia (третьего порядка); так как в названных узлах происходит синапс вставочных и эфферентных нейронов, то отмеченная разница между симпатическим и парасимпатическим отделами вегетативной нервной системы связана именно с этими нейронами.
Аксоны эфферентных вегетативных нейронов почти лишены миелина - безмякотные (серые). Они составляют послеузловые волокна, rami postganglionares. Послеузловые волокна симпатической нервной системы, отходящие от узлов пограничного симпатического ствола, расходятся в двух направлениях. Одни волокна идут к внутренностям и составляют висцеральную часть симпатической системы. Другие волокна образуют rami communicantes grisei, соединяющие симпатический ствол с анимальными нервами. В составе последних волокна достигают соматических органов (аппарата движения и кожи), в которых иннервируют гладкую мускулатуру сосудов и кожи, а также железы.
Совокупность описанных эфферентных вегетативных волокон, идущих от узлов симпатического ствола до органов сомы, составляет соматическую часть симпатической системы. Такая структура обеспечивает функцию вегетативной нервной системы, которая регулирует обмен веществ всех частей организма применительно к непрерывно изменяющимся условиям среды и условиям функционирования (работы) тех или иных органов и тканей.
Соответственно этой наиболее универсальной своей функции, связанной не с какими-либо отдельными органами и системами, а со всеми частями, со всеми органами и тканями организма, вегетативная система и морфологически характеризуется универсальным, повсеместным распространением в организме, проникая во все его органы и ткани.
Следовательно, симпатическая нервная система иннервирует не только внутренности, но и сому, обеспечивая в ней обменные и трофические процессы.
В результате каждый орган, по И. П. Павлову, находится под тройным нервным контролем, в связи с чем он различает три вида нервов: 1) функциональные, осуществляющие функцию данного органа; 2) сосудодвигательные, обеспечивающие грубую доставку крови к органу, и 3) трофические, регулирующие усвоение из доставленной крови питательных веществ.
Висцеральная часть симпатической системы содержит все эти три вида нервов для внутренностей, а соматическая часть, симпатической системы - только сосудодвигательные и трофические. Что же касается функциональных нервов для органов сомы (скелетной мускулатуры и др.), то они идут в составе соматической, анимальной, нервной системы.
Таким образом, основное отличие эфферентной части вегетативной нервной системы от эфферентной части анимальной заключается в том, что анимальные, соматические нервные волокна, выйдя из центральной нервной системы, идут до рабочего органа нигде не прерываясь, тогда как вегетативные волокна на своем пути от мозга до рабочего органа прерываются в одном из узлов первого, второго или третьего порядка. Вследствие этого эфферентный путь вегетативной системы разбивается на две части, из которых он и состоит: предузловые миелиновые (мякотные) волокна, rami preganglionares, и послеузловые, лишенные миелина (безмякотные) волокна, rami postganglionares.
Наличие узлов в эфферентной части рефлекторной дуги составляет характерный признак вегетативной нервной системы, отличающий ее от анимальной (см. рис. 341).
Определенные отличия имеют и нервы. Афферентные пути вегетативной системы не имеют характера макроскопически видимых нервов, а их волокна идут в составе других нервов (nn. splanchnici major et minor, n. vagus, задние корешки и др.). При этом для симпатической системы характерно то, что связанная с ней чувствительная иннервация может распространяться на значительные расстояния и, следовательно, симпатическая система может рассматриваться как система окольной иннервации (Д. М. Голуб, 1963).
Так, например, афферентные спинальные нервные волокна, участвующие в формировании солнечного сплетения, иннервирующего органы брюшной полости, происходят из многочисленных спинномозговых узлов (С5-L3). Это обстоятельство определяет множественность и многосегментарность путей и источников афферентной иннервации органов брюшной полости. Этим же объясняется и то, что чувство боли от внутренностей может передаваться как по вегетативным, так и по анимальным нервам (Mitchell, 1953).
Наряду с этим в последнее время установлено (Д. М. Голуб, 1963), что имеется не только диффузность в отношении распределения чувствительных нейронов и путей хода волокон, но и преимущественное участие определенных спинномозговых узлов в иннервации внутренностей. Следовательно, среди источников и путей афферентной иннервации внутренностей можно выделить основные и дополнительные. Это деление тесно связано с представлением об окольных путях афферентной спинальной иннервации внутренностей. Окольные пути в патологических условиях (перерыв спинного мозга и др.) могут играть роль компенсаторных путей, возмещающих функцию нарушенных основных путей, компенсаторных приспособлений в виде "перекрытия" в афферентной иннервации органов.
Что же касается эфферентных путей вегетативной системы, то они образуют ясно выраженные нервы и узлы. Поэтому можно говорить о двух центробежных путях единой нервной системы: один путь - это анимальные, соматические, двигательные нервы, а другой - вегетативные. Вегетативные нервы образуют сплетения вокруг кровеносных сосудов, вместе с которыми они подходят и входят в органы. Наличие сплетений вокруг сосудов составляет характерный признак вегетативной нервной системы, отличающий ее от анимальной.
Как уже говорилось выше, вегетативная нервная система характеризуется универсальным, повсеместным распространением в организме. Она имеет широкую область эфферентной иннервации, охватывающую все органы и ткани тела, не исключая и скелетной мускулатуры (последнюю она тонизирует). В этом и состоит морфологическая особенность вегетативной нервной системы в противоположность анимальной, которая иннервирует центробежными волокнами только скелетные мышцы, т. е. имеет сравнительно ограниченную область эфферентной иннервации.
Для понимания строения ее необходимо учитывать развитие вегетативной нервной системы*.
* (Излагается частично по А. Г. Кнорре, И. Д. Лев и Е. К. Сеппу.)
Гладкая мускулатура беспозвоночных регулируется ганглиозно-сетевидной нервной системой, которая, кроме этой специальной функции, регулирует также и обмен веществ. Приспособление, прилаживание уровня обмена веществ к изменяющейся функции органов называется адаптацией (adaptare - прилаживать), а соответственная функция нервной системы - адаптационно-трофической (Л. А. Орбели). Адаптационно-трофическая функция есть наиболее общая и весьма древняя функция нервной системы, существовавшая у примитивных предков позвоночных. В дальнейшем ходе эволюции сильнее всего прогрессировали аппарат движения (развитие твердого скелета и поперечнополосатой мускулатуры) и органы чувств, т. е. органы животной жизни. Поэтому та часть нервной системы, которая была связана с ними, т. е. анимальная часть нервной системы, претерпела наиболее резкие изменения и приобрела новые признаки, в частности: изоляцию волокон при помощи миелиновых оболочек (мякотные волокна), большую скорость проведения возбуждения (12-100 м/с). Напротив, органы растительной жизни претерпели более медленную и менее прогрессивную эволюцию, поэтому связанная с ними часть нервной системы сохранила за собой наиболее общую функцию - адаптационно-трофическую. Эта часть нервной системы и есть вегетативная нервная система.
Наряду с некоторой специализацией она сохранила ряд древних примитивных черт: отсутствие у большинства нервных волокон миелиновых оболочек (безмякотные волокна), меньшую скорость проведения возбуждения (0,3-10 м/с), а также меньшую концентрацию и централизацию эффекторных нейронов, оставшихся разбросанными на периферии, в составе ганглиев, нервов и сплетений. При этом эффекторный нейрон оказался расположенным вблизи рабочего органа или даже в толще его.
Такое периферическое расположение эффекторного нейрона обусловило главную морфологическую особенность вегетативной нервной системы - двухнейронность эфферентного периферического пути, состоящего из вставочного и эффекторного нейронов.
С появлением туловищного мозга (у бесчерепных) возникающие в нем импульсы адаптации идут по вставочным нейронам, обладающим быстрыми темпами возбуждения; выполняется же адаптация гладкой мускулатурой и железами, к которым подходят эффекторные нейроны, отличающиеся медленной проводимостью (о чем говорилось выше). Это противоречие разрешается в процессе эволюции благодаря развитию специальных нервных узлов, в которых устанавливаются контакты вставочных нейронов с эффекторными, причем один вставочный нейрон вступает в связь со многими эффекторными (примерно 1:32). Этим достигается переключение мякотных волокон, обладающих большой скоростью проведения раздражений, на безмякотные, обладающие малой скоростью. В результате весь эфферентный периферический путь вегетативной нервной системы разбивается на две части - предузловую и послеузловую, а сами узлы становятся трансформаторами темпов возбуждения с быстрых на медленные.
У низших рыб, когда образуется головной мозг, в нем развиваются центры, объединяющие деятельность органов, вырабатывающих внутреннюю среду организма.
Так как в этой деятельности, кроме гладкой мускулатуры, принимает участие и скелетная поперечнополосатая, то возникает потребность в координации работы гладких и поперечнополосатых мышц. Например, жаберные крышки приводятся в движение скелетной мускулатурой; так же и у человека в акте дыхания участвует как гладкая мускулатура бронхов, так и поперечнополосатая грудной клетки. Такую координацию осуществляет развивающийся в заднем мозгу специальный рефлекторный аппарат в виде системы блуждающего нерва (бульбарный отдел парасимпатической нервной системы).
В центральной нервной системе возникают и другие образования, которые подобно блуждающему нерву выполняют функцию координации совместной деятельности скелетной мускулатуры, обладающей быстрыми темпами возбуждения, и гладкой мускулатуры и желез, обладающих медленными темпами. Сюда относится та часть глазодвигательного нерва, которая осуществляет при помощи гладких и поперечнополосатых мышц глаза стандартную установку ширины зрачка, аккомодации и конвергенции соответственно силе освещения и расстоянию до рассматриваемого объекта по тем же принципам, как это делает фотограф (мезенцефалический отдел парасимпатической нервной системы). Сюда относится и та часть крестцовых нервов (II-IV), которые осуществляют стандартную функцию тазовых органов (мочевого пузыря и прямой кишки) - опорожнение, в которой участвуют как гладкие мышцы названных органов, так и поперечнополосатые мышцы таза и брюшного пресса - сакральный отдел парасимпатической системы. В среднем и промежуточном мозгу развился центральный адаптационный аппарат в виде серого вещества вокруг сильвиева водопровода и серого бугра (hypothalamus).
Наконец, в коре мозга возникли центры, объединяющие высшие анимальные и вегетативные функции.
Развитие вегетативной нервной системы в онтогенезе (эмбриогенезе) идет иначе, чем в филогенезе.
Вегетативная часть нервной системы возникает из общего с анимальной частью источника - нейроэктодермы, чем доказывается единство всей нервной системы.
Из общего зачатка нервной системы выселяются симпатобласты, которые скапливаются в определенных местах, образуя сначала узлы симпатического ствола, а затем промежуточные узлы, а также нервные сплетения. Отростки клеток симпатического ствола, объединяясь в пучки, образуют rami communicantes grisei.
Сходным образом развивается и головная часть вегетативной нервной системы. Зачатки парасимпатических узлов выселяются из продолговатого мозга или ганглиозной пластинки и совершают дальнюю миграцию вдоль ветвей тройничного, блуждающего и других нервов, оседая по их ходу или образуя интрамуральные ганглии.
Образование новых нервных путей (реиннервация органов)
Экспериментальными исследованиями Д. М. Голуба и его сотрудников (I960, 1964) доказано при подшивании некоторых внутренних органов друг к другу (тонкой и толстой кишки к мочевому пузырю) возникновение новых афферентных путей, связывающих эти органы с центральной нервной системой (со спинным мозгом).
Возможность управления развитием нервной системы и образования новых афферентных путей (реиннервация) имеет практическое значение для хирургического восстановления органов и их нарушенной нервной системы.
Симпатическая нервная система
Исторически симпатическая система возникает как сегментарный отдел, поэтому и у человека она имеет сегментарный характер строения.
Центральный отдел симпатической системы
Центральный отдел симпатической системы располагается в боковых рогах спинного мозга на уровне CVIII, ThI-LIII, в nucleus intermediolateralis. От него отходят волокна, иннервирующие гладкую мускулатуру внутренностей, органов чувств (глаз), железы. Кроме того, здесь располагаются сосудодвигательные, пиломоторные и потоотделительные центры. Считают (и это подтверждается клиническим опытом), что различные отделы спинного мозга оказывают влияние на трофику, терморегуляцию и обмен веществ.
Периферический отдел симпатической системы
Периферический отдел симпатической системы образуется прежде всего двумя симметричными стволами, triincus sympathicus dexter et sinister, расположенными по бокам позвоночника на всем его протяжении от основания черепа до копчика, где оба ствола своими каудальными концами сходятся в одном общем узле. Каждый из этих двух симпатических стволов слагается из ряда нервных узлов первого порядка, соединяющихся между собой посредством продольных межузловых ветвей, rami interganglionares, состоящих из нервных волокон. Кроме узлов симпатических стволов (ganglia trunci sympdthici), в состав симпатической системы входят указанные выше ganglia intermedia.
По новейшим данным (В. С. Абашидзе, 1963), симпатический ствол, начиная с верхнего шейного узла, содержит элементы парасимпатической и даже анимальной нервной системы.
Отростки клеток, заложенных в боковых рогах тораколюмбального отдела спинного мозга, выходят из спинного мозга через передние корешки и, отделившись от них, идут в составе rami communicantes dlbi к симпатическому стволу (см. рис. 341). Здесь они или соединяются синапсом с клетками узлов симпатического ствола, или же, пройдя через его узлы без перерыва, достигают одного из промежуточных узлов. Это так называемый преганглионарный путь. От узлов симпатического ствола или (если там не было перерыва) от промежуточных узлов отходят безмякотные волокна постганглионарного пути, направляющиеся к кровеносным сосудам и внутренностям.
Поскольку симпатическая система имеет соматическую часть, она связана со спинномозговыми нервами, обеспечивающими иннервацию сомы. Эта связь осуществляется посредством серых соединительных ветвей, rami communicdntes grisei, которые представляют собой участок постганглионарных волокон на протяжении от узлов симпатического ствола до n. spinalis. В составе rami communicantes grisei и спинномозговых нервов постганглионарные волокна распространяются в сосудах, железах и гладких мышцах кожи туловища и конечностей, а также в поперечнополосатой мускулатуре, обеспечивая ее трофику и тонус.
Таким образом, симпатическая нервная система соединяется с анимальной посредством двоякого рода соединительных ветвей: белых и серых, rami communicantes albi et grisei. Белые соединительные ветви (мякотные) являются преганглионарными волокнами. Они идут от центров симпатической системы через передние корешки к узлам симпатического ствола. Поскольку центры лежат на уровне грудных и верхних поясничных сегментов, то и rami communicantes albi имеются лишь в пределах от I грудного до III поясничного спинномозгового нерва. Rami communicantes grisei, постганглионарные волокна, обеспечивают вазомоторные и трофические процессы сомы; они соединяют пограничный ствол со спинномозговыми нервами на всем его протяжении. Шейный отдел симпатического ствола имеет связь и с головными нервами. Следовательно, все сплетения анимальной нервной системы содержат в составе своих пучков и нервных стволов волокна симпатической системы, чем подчеркивается единство этих систем.
Симпатический ствол
Каждый из двух симпатических стволов подразделяют на четыре отдела: шейный, грудной, поясничный (или брюшной) и крестцовый (или тазовый).
Шейный отдел идет от основания черепа до шейки I ребра; симпатический ствол располагается позади сонных артерий на глубоких мышцах шеи. В его состав входят три шейных симпатических узла - верхний, средний и нижний.
Ganglion cervicale superius является самым крупным узлом симпатического ствола, имея в длину около 20 мм и ширину 4-6 мм. Лежит он на уровне II и части III шейных позвонков позади внутренней сонной артерии и медиально от n. vagus.
Ganglion cervicale medium, небольшой величины, располагается обыкновенно в месте перекреста a. thyreoidea inferior с сонной артерией, нередко отсутствует или может распадаться на два узелка.
Ganglion cervicale inferius, довольно значительной величины, расположен позади начальной части позвоночной артерии; нередко сливается с I, а иногда и II грудным узлом, образуя общий звездчатый узел, ganglion cervicothoracicum, s. ganglion stellatum. Некоторые авторы (Whrete, 1959) описывают 4 шейных узла симпатического ствола, которые связаны с развитием сегментарных артерий: верхний, средний, нижний и звездчатый.
От шейных узлов отходят нервы для головы, шеи и груди. Их можно разделить на восходящую группу, направляющуюся к голове, на нисходящую - опускающуюся к сердцу, и группу для органов шеи, направляющуюся к ним почти непосредственно от места отхода.
Нервы для головы отходят от верхнего и нижнего шейных узлов и делятся на группу, проникающую в полость черепа, и группу, подходящую к голове снаружи.
Первая группа представлена n. caroticus interims, отходящим от верхнего шейного узла, и n. vertebralis, отходящим от нижнего шейного узла. Оба нерва, сопровождая одноименные артерии, образуют вокруг них сплетения: plexus caroticus internus и plexus vertebralis; вместе с артериями они проникают в полость черепа, где анастомозируют между собой и дают ветви к сосудам мозга, оболочкам, гипофизу, стволам III, IV, V, VI пар головных нервов и барабанному нерву.
Plexus caroticus internus продолжается в plexus cavernosus, который окружает a. carotis interna на участке прохождения ее через sinus cavernosus.
Ветви сплетений распространяются, кроме самой внутренней сонной артерии, также по ее разветвлениям. Из ветвей plexus caroticus internus следует отметить n. petrosus profundus, который присоединяется к n. petrosus major и вместе с ним образует n. canalis pterygoidei, подходящий через одноименный канал к ganglion pterygopalatinum.
Вторая группа симпатических нервов головы, наружная, составляется двумя ветвями верхнего шейного узла, nn. carotid externi, которые, образовав сплетения вокруг наружной сонной артерии, сопровождают ее разветвления на голове. От сплетения отходит стволик к ушному узлу, g. oticum; от лицевого сплетения, plexus facialis, сопровождающего одноименную артерию, отходит ветвь к подчелюстному узлу.
Через посредство ветвей, входящих в сплетения вокруг сонной артерии и ее ветвей, верхний шейный узел дает волокна к сосудам (вазоконстрикторы) к железам головы: потовым, слезной, слизистым и слюнным, а также к гладким мышцам волос и к мышце, расширяющей зрачок (см. "Орган зрения"), m. dilatator pupillae. Центр расширения зрачка, centrum ciliospinale, находится в спинном мозгу на уровне от VIII шейного до II грудного сегмента.
Органы шеи получают нервы от всех трех шейных узлов; кроме того, часть нервов отходит от межузловых участков шейного отдела симпатического ствола, а часть - от сплетений сонных артерий.
Веточки от сплетений следуют по ходу ветвей наружной сонной артерии, носят одноименные названия и вместе с ними подходят к органам, в силу чего число отдельных симпатических сплетений равно числу артериальных ветвей. Из нервов, отходящих от шейной части пограничного ствола, отмечают гортаноглоточные ветви от верхнего шейного узла - rami laryngopharyngei, которые частью идут с n. laryngeus superior (ветвь n. vagi) к гортани, частью спускаются к боковой стенке глотки; здесь они вместе с ветвями языкоглоточного, блуждающего и верхнего гортанного нервов образуют глоточное сплетение - plexus pharyngeus.
Нисходящая группа ветвей шейной части симпатического ствола представлена nn. cardiaci cervicales superior, medius et inferior, отходящими от соответствующих шейных узлов. Шейные сердечные нервы спускаются в грудную полость, где вместе с симпатическими грудными сердечными нервами и ветвями блуждающего нерва участвуют в образовании сердечных сплетений (см. иннервацию сердца).
Грудной отдел симпатического ствола располагается впереди шеек ребер, прикрыт спереди плеврой. В его состав входят 10-12 узлов более или менее треугольной формы. Грудной отдел характеризуется присутствием белых соединительных ветвей, rami communicantes dlbi, соединяющих передние корешки спинномозговых нервов с узлами симпатического ствола. Ветви грудного отдела: 1) nn. cardiaci thoracici отходят от верхних грудных узлов и участвуют в образовании plexus cardiacus (подробное описание сердечных сплетений см. при описании сердца); 2) rami communicantes grisei, безмякотные - к межреберным нервам (соматическая часть симпатической системы); 3) rami pulmonales - к легким, образуют plexus pulmonalis;
4) rami aortici образуют сплетение на грудной аорте, plexus aorticus thoracicus, и частью на пищеводе, plexus esophageus, а также на грудном протоке (во всех указанных сплетениях принимает участие и n. vagus);
5) nn. splanchnici major et minor - большой и малый чревные нервы; n. splanchnicus major начинается несколькими корешками, отходящими от V-IX грудных узлов; корешки n. splanchnicus major идут в медиальном направлении и сливаются на уровне IX грудного позвонка в один общий ствол, проникающий через промежуток между мышечными пучками ножек диафрагмы в брюшную полость, где он входит в состав plexus celiacus; n. splanchnicus minor начинается от X-XI грудных узлов и также входит в plexus celiacus, проникая через диафрагму вместе с большим чревным нервом или отделенный от него несколькими мышечными пучками. В чревных нервах проходят сосудосуживающие волокна, как это видно из того обстоятельства, что при перерезке этих нервов сосуды кишечника сильно переполняются кровью; в nn. splanchnici содержатся, кроме того, волокна, тормозящие движение желудка и кишок, а также волокна, служащие проводниками ощущений от внутренностей (афферентные волокна симпатической системы).
Поясничный, или брюшной, отдел симпатического ствола состоит из четырех, иногда из трех узлов. Симпатические стволы в поясничном отделе расположены на более близком расстоянии один от другого, чем в грудной полости, так что узлы лежат на переднебоковой поверхности поясничных позвонков вдоль медиального края m. psoas major. Rami communicantes dlbi имеются только с двумя или тремя верхними поярничными нервами.
От брюшного отдела симпатического ствола на всем протяжении отходит большое количество ветвей, которые вместе с nn. splanchnici major et minor и брюшными отделами блуждающих нервов образуют самое большое непарное чревное, или солнечное, сплетение, plexus celiacus. В формировании солнечного сплетения участвуют также многочисленные спинномозговые узлы (С5-L3) (Д. М. Голуб, 1963). Оно лежит на передней полуокружности брюшной аорты, позади поджелудочной железы, и окружает начальные части чревного ствола (truncus celiacus) и верхней брыжеечной артерии. Сплетение занимает участок между почечными артериями, надпочечниками и аортальным отверстием диафрагмы и включает парный узел чревной артерии, ganglion celiacum, и иногда непарный узел верхней брыжеечной артерии, ganglion mesentericum superius, лежащий под корнем последней.
От чревного сплетения отходит ряд меньших парных сплетений к диафрагме, надпочечникам, почкам, а также plexus testiculdris (ovaricus), следующих по ходу одноименных артерий. Имеется также ряд непарных сплетений, к отдельным органам по стенкам артерий, название которых они носят. Из последних верхнее брыжеечное сплетение, pl. mesentericus superior, снабжает поджелудочную железу, тонкую и толстую кишку до половины протяжения поперечной ободочной, а также яичник.
Вторым главным источником иннервации органов полости живота является сплетение на аорте, plexus aorticus abdominalis, составленное из двух стволов, отходящих от чревного сплетения, и веточек от поясничных узлов симпатического ствола. От аортального сплетения отходит нижнее брыжеечное сплетение, plexus mesentericus inferior, для поперечной и нисходящей части ободочной кишки, сигмовидной и верхних отделов rectum (pl. rectales superiores). У места отхождения plexus mesentericus inf. располагается одноименный узел, g. mesentericum inferius. Его постганглионарные волокна идут в тазе в составе nn. hypogastrici.
Аортальное сплетение продолжается вначале в непарное верхнее подчревное сплетение, pl. hypogastricus superior, которое у мыса раздваивается и переходит в сплетение таза, или нижнее подчревное сплетение (pl. hypogastricus inferior s. pl. pelvinus). Волокна, происходящие из верхних поясничных сегментов, по своей функции являются сосудодвигательными (вазоконстрикторами) для полового члена, двигательными для матки и сфинктера мочевого пузыря.
Крестцовый, или тазовый, отдел имеет обычно четыре узла; располагаясь на передней поверхности крестца вдоль медиального края передних крестцовых отверстий, оба ствола книзу постепенно сближаются друг с другом и затем оканчиваются в одном общем непарном узле - ganglion impar, находящемся на передней поверхности копчика. Узлы тазового отдела, как и поясничного, связаны между собой не только продольными, но и поперечными стволиками.
От узлов крестцового отдела симпатического ствола отходит ряд ветвей, которые соединяются с ветвями, отделяющимися от нижнего брыжеечного сплетения, и образуют пластинку, протягивающуюся от крестца к мочевому пузырю; это так называемое нижнее подчревное или тазовое сплетение, pl. hypogastricus inferior s. pl. pelvinus. Сплетение имеет свои узелки - ganglia pelvina. В сплетении различают несколько отделов: 1)передненижний отдел, в котором выделяют верхнюю часть, иннервирующую мочевой пузырь, - plexus vesicdlis, и нижнюю, снабжающую у мужчин предстательную железу (pl. prostdtlcus), семенные пузырьки и семявыносящий проток (pl. deferent Idlis) и пещеристые тела (nn. cavernosi penis); 2) задний отдел сплетения снабжает прямую кишку (pl. rectdles medii et infer lores). У женщин выделяют еще 3) средний отдел, нижняя часть которого дает ветви к матке и влагалищу (pi. uterovagindlis), пещеристым телам клитора (nn. covernosl clltoridls), а верхняя - к матке и яичникам.
Парасимпатическая система
Парасимпатическая система исторически развивается как надсегментарный отдел, и поэтому центры ее располагаются не только в спинном мозгу, но и в головном.
Центры парасимпатической системы
Центральная часть парасимпатической системы состоит из головного, или краниального, отдела и спинномозгового, или сакрального, отдела. Некоторые авторы (Л. Я. Пинес, 1957) считают, что парасимпатические центры располагаются в спинном мозгу не только в области крестцовых сегментов, но и в других отделах его, в частности в пояснично-грудном отделе между передним и задним рогом, в так называемой интермедиарной зоне. Центры дают начало эфферентным волокнам задних корешков, вызывающих расширение сосудов, задержку потоотделения и торможение сокращения гладких мышц волос в области туловища и конечностей.
Краниальный отдел в свою очередь состоит из центров, заложенных в среднем мозгу (мезэнцефалическая часть) и в ромбовидном мозгу - в мосте и продолговатом мозгу (бульбарная часть).
1. Мезэнцефалическая часть представлена nucleus accessorius n. oculomotorii (ядро Якубовича) и срединным непарным ядром, за счет которых иннервируется гладкая мускулатура глаза - m. sphincter pupillae и m. ciliaris.
2. Бульварная часть представлена milceus salivatorius superior n. facialis (точнее, n. intermedins), nucleus salivatorius inferior n. glossopharyngei и nucleus dorsalis n. vagi (см. соответствующие нервы).
Сакральный отдел (рис. 342). Парасимпатические центры лежат в спинном мозгу, в nucleus intermediolateralis бокового рога на уровне II-IV крестцовых сегментов.
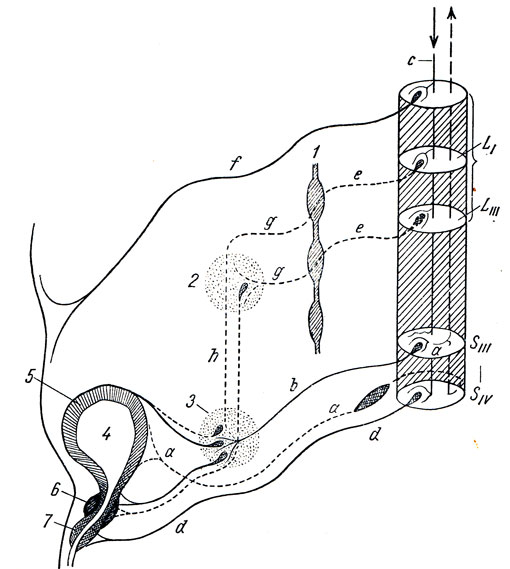
Рис. 342. Схема иннервации мочевого пузыря. а - чувствительный путь к спинному мозгу; b - n. splanchnicus pelvinus; с - кортикоспиналь-ный путь d - n. pudendus; е - rr. communicantes; f - n. iliohypogastricus; gg - nn. mesenterici; h - nn. hypogastrici; 1 - симпатическая цепочка; 2 - ganglion mesentericum inf.; 3 - pl. hypogastricus и расположенные поблизости терминальные узлы парасимпатической системы; 4 - vesica urinaria; 5 - m. detrussor; 6 - m. sphincter vesicae; 7 - m. sphincter urethrae
Периферический отдел парасимпатической системы
Периферическая часть краниального отдела парасимпатической системы представлена: 1) преганглионарными волокнами, идущими в составе III, VII, IX и X пар головных нервов (по Mitchell, 1957, и в составе I и XI); 2) терминальными узлами, расположенными вблизи органов, а именно: ganglia ciliare, pterygopalatinum, submandibular, oticum и 3) постганглионарными волокнами; постганглионарные волокна имеют или самостоятельный ход, как, например, nn. ciliares breves, отходящие от ganglion ciliare, или идут в составе каких-либо нервов, как, например, постганглионарные волокна, отходящие от ganglion oticum и идущие в составе n. auriculotemporalis. Некоторые авторы (Гассер, 1955; Гесс, 1956; Митчелл, 1957, и др.) указывают, что парасимпатические волокна выходят также и из различных сегментов спинного мозга через дорсальные корешки, направляясь к стенкам туловища и конечностей.
Периферическая часть сакрального отдела парасимпатической системы представлена волокнами, которые в составе передних корешков II-IV крестцовых нервов и далее в составе их передних ветвей, образующих plexus sacralis (анимальное сплетение), входят в малый таз. Здесь они отделяются от сплетения и в виде nn. splanchnici pelvini направляются к plexus hypogastricus inferior, иннервируя вместе с последним тазовые внутренности: прямую кишку с colon sigmoideum, мочевой пузырь, наружные и внутренние половые органы. Раздражение nn. splanchnici pelvini вызывает сокращение прямой кишки и мочевого пузыря (m. detrusor urinae) с ослаблением их сфинктеров. Волокна же симпатического подчревного сплетения задерживают опорожнение этих органов; они же возбуждают сокращение матки, тогда как nn. splanchnici pelvini его тормозят. Nn. splanchnici pelvini содержат в себе еще сосудорасширяющие волокна (nn. erigentes) для corpora cavernosa penis et clitoridis, обусловливающие эрекцию. Парасимпатические волокна, отходящие от сакрального отдела спинного мозга, идут в тазовые сплетения не только в составе nn. erigentes et nn.splanchnici pelvini, но и в составе nervus pudendus (преганглионарные волокна). Срамной нерв является сложным нервом, содержащим в своем составе, кроме анимальных волокон, также и вегетативные (симпатические и парасимпатические), участвующие в нижнем подчревном сплетении. Симпатические волокна, отходящие от узлов крестцового отдела симпатического ствола в качестве постганглионарных, присоединяются к срамному нерву в полости малого таза и проходят через нижнее подчревное сплетение к тазовым органам.
К парасимпатической нервной системе относится также так называемая интрамуральная нервная система.
В стенках ряда полостных органов находятся нервные сплетения, содержащие мелкие узлы (терминальные) с ганглиозными клетками и безмякотными волокнами - ганглиозно-сетевидная, или интрамуральная, система.
Интрамуральная система особенно выражена в пищеварительном тракте, где она представлена несколькими сплетениями.
1. Мышечное сплетение, plexus myentericus Auerbachi лежащее между продольной и кольцевой мускулатурой пищеварительной трубки.
2. Подслизистое сплетение, plexus submucosus Meissneri, находящееся в подслизистой ткани. Последнее переходит в сплетение желез и ворсинок.
К периферии от названных сплетений располагается диффузная нервная сеть, "сеть Леонтовича". К сплетениям подходят нервные волокна от симпатической и парасимпатической систем. В интрамуральных сплетениях происходит переключение предузловых волокон парасимпатической системы на послеузловые.
Краткий обзор вегетативной иннервации органов
(Афферентные пути вегетативной иннервации даны в основном по Mitchell (1957).)
Иннервация глаза. В ответ на определенные зрительные раздражения, идущие от сетчатки, осуществляется конвергенция и аккомодация зрительного аппарата.
Конвергенция глаз - сведение зрительных осей обоих глаз на рассматриваемом предмете - происходит рефлекторно, сочетанным сокращением поперечнополосатых мышц глазного яблока. Этот рефлекс, необходимый для бинокулярного зрения, связан с аккомодацией глаза. Аккомодация - способность глаза ясно видеть предметы, находящиеся от него на различных расстояниях, - зависит от сокращения гладкой мускулатуры - m. ciliaris и m. sphincter pupillae. Поскольку деятельность гладкой мускулатуры глаза осуществляется совместно с сокращением его поперечнополосатых мышц, вегетативная иннервация глаза будет рассмотрена вместе с анимальной иннервацией его двигательного аппарата.
Афферентным путем от мышц глазного яблока (проприоцептивная чувствительность) являются, по одним авторам, сами анимальные нервы, иннервирующие данные мышцы (III, IV, VI головные нервы), по другим - n. ophthalmicus (n. trigemini).
Центры иннервации мышц глазного яблока - ядра III, IV и VI пар. Эфферентный путь - III, IV и VI головные нервы. Конвергенция глаза осуществляется, как указывалось, сочетанным сокращением мышц обоих глаз.
Надо иметь в виду, что изолированных движений одного глазного яблока вообще не существует. В любых произвольных и рефлекторных движениях всегда участвуют оба глаза. Эта возможность сочетанного движения глазных яблок (взора) обеспечивается особой системой волокон, связывающей между собой ядра III, IV и VI нервов и носящей название медиального продольного пучка.
Медиальный продольный пучок начинается в ножках мозга от ядра Даркшевича, соединяется с ядрами III, IV, VI нервов при помощи коллатералей и направляется по мозговому стволу вниз в спинной мозг, где заканчивается, по-видимому, в клетках передних рогов верхних шейных сегментов. Благодаря этому движения глаз сочетаются с движениями головы и шеи (Ф. А. Поемный и Е. П. Семенова, 1960).
Иннервация гладких мышц глаза - гл. sphincter pupillae и m. ciliaris, осуществляющих аккомодацию глаза, происходит за счет парасимпатической системы; иннервация m. dilatator pupillae - за счет симпатической. Афферентными путями вегетативной системы является n. oculomotorius и n. ophthalmicus.
Эфферентная парасимпатическая иннервация. Преганглионарные волокна идут из ядра Якубовича (мезенцефалический отдел парасимпатической нервной системы) в составе n. oculomotorius и по его radix oculomotoria достигают ganglion ciliare (рис. 343), где и оканчиваются. В ресничном узле начинаются постганглионарные волокна, которые через nn. ciliares breves доходят до ресничного мускула и круговой мышцы радужной оболочки. Функция: сужение зрачка и аккомодация глаза к дальнему и близкому видению.
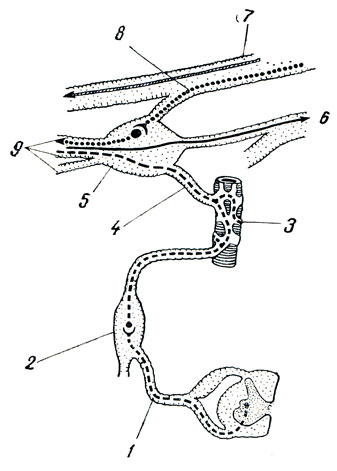
Рис. 343. Ganglion ciliare (схема). 1 - r. communicans albus; 2 - ganglion cervicale superins; 3 - a. ophthalmica; 4 - r. sympathicus ad ganglion ciliare; 5 - ganglion ciliare; 6 - n. nasociliaris; 7 - n. oculomotorius; 8 - radix oculomotoria (парасимпатические преганглионарные волокна); 9 - nn. ciliares breves
Эфферентная симпатическая иннервация. Преганглионарные волокна идут из клеток nucleus intermediolateralis боковых рогов последнего шейного и двух верхних грудных сегментов (CVIII-ThII, centrum ciliospinale), выходят через две верхние грудные rami communicantes albi, проходят в составе шейного отдела симпатического ствола и кончаются в верхнем шейном узле. Постганглионарные волокна идут в составе n. caroticus internus в полость черепа и вступают в plexus caroticus internus и plexus ophtalmicus; после этого часть волокон проникает в ramus communicans, соединяющуюся с n. nasociliaris и nervi ciliares longi, а часть направляется к ресничному узлу, через который проходит, не прерываясь, в nervi ciliares breves. И те и другие симпатические волокна, проходящие через длинные и короткие ресничные нервы, достигают радиальной мышцы радужной оболочки. Функция: расширение зрачка, а также сужение сосудов глаза.
Иннервация желез слезной и слюнных. Афферентным путем для слезной железы является n. lacrimalis (ветвь n. ophthalmicus от n. trigemini), для подчелюстной и подъязычной - n. lingualis (ветвь n. mandibularis от n. trigemini) и chorda tympani (ветвь n. intermedius), для околоушной - n. auriculotemporais и n. glossopharyngeus.
Эфферентная парасимпатическая иннервация слезной железы. Центр лежит в верхнем отделе продолговатого мозга (В. М. Бехтерев) и связан с ядром промежуточного нерва (nucleus salivatorius superior). Преганглионарные волокна идут в составе n. intermedius, далее n. petrosus major до ganglion pterygopalatinum (рис. 344). Отсюда начинаются постганглионарные волокна, которые в составе n. maxillaris и далее его ветви n. zygomatics через связи с n. lacrimals достигают слезной железы.
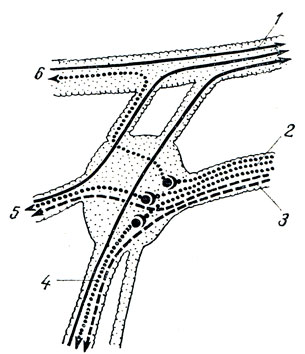
Рис. 344. Ganglion pterygopalatinum (схема). 1 - n. maxillaris; 2 - n. petrosus major; 3 - n. petrosus profundus; 4 - nn. palatini; 5 - nn. nasales posteriores; 6 - n. zygomaticus
Эфферентная парасимпатическая иннервация подчелюстной и подъязычной желез. Преганглионарные волокна идут от nucleus salivatorius superior в составе n. intermedius, далее chorda tympani и n. lingualis до ganglion submandibular, откуда начинаются постганглионарные волокна, достигающие желез в составе язычного нерва.
Эфферентная парасимпатическая иннервация околоушной железы. Преганглионарные волокна идут от nucleus salivatorius inferior в составе n. glossopharyngeus, далее n. tympanicus, n. petrosus minor до ganglion oticum (рис. 345). Отсюда начинаются постганглионарные волокна, идущие к железе в составе n. auriculotemporalis. Функция: усиление секреции слезной и названных слюнных желез; расширение сосудов желез.
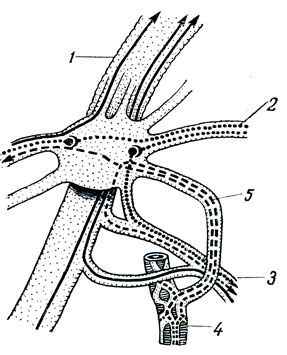
Рис. 345. Ganglion oticum (схема). 1 - n. mandibularis; 2 - n. petrosus minor; 3 - n. auriculotemporalis; 4 - a. meningea media; 5 - симпатические волокна
Эфферентная симпатическая иннервация всех названных желез. Преганглионарные волокна начинаются в боковых рогах верхних грудных сегментов спинного мозга и заканчиваются в верхнем шейном узле. Постганглионарные волокна начинаются в названном узле и доходят до слезной железы в составе plexus caroticus internus, до околоушной - в составе plexus caroticus externus и до подчелюстной и подъязычной желез - через plexus caroticus externus и затем через plexus facialis. Функция: задержка отделения слюны (сухость во рту). Слезотечение (влияние не резкое).
Иннервация сердца (рис. 346). Афферентные пути от сердца идут в составе n. vagus, а также в среднем и нижнем шейных и грудных сердечных симпатических нервах. При этом по симпатическим нервам проводится чувство боли, а по парасимпатическим - все остальные афферентные импульсы (Mitchell, 1957).
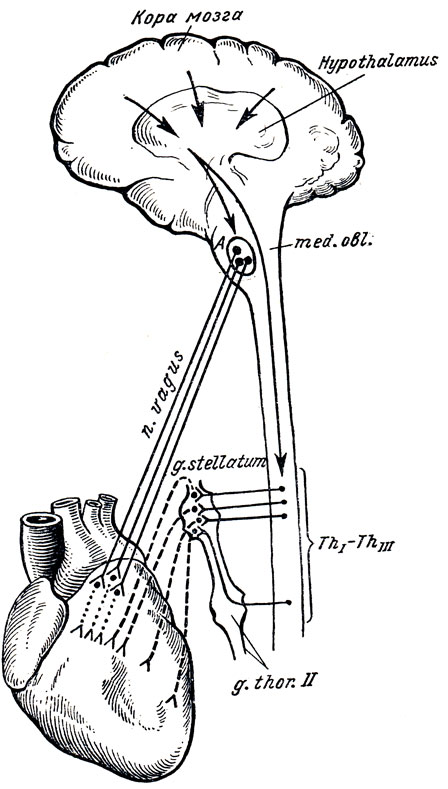
Рис. 346. Схема иннервации сердца. A - ядро сердечных волокон блуждающего нерва в продолговатом мозгу; ТhI-ThIII - сегменты спинного мозга, в которых расположены клетки, отдающие симпатические нервные волокна к сердцу; g. thor. II - второй грудной симпатический ганглий. Сплошные линии - преганглионарные волокна блуждающего и симпатических нервов; пунктир - постганглионарные волокна блуждающего нерва в сердце; прерывистые линии - постганглионарные симпатические волокна, идущие к сердцу. Стрелки - влияние коры мозга, передаваемое через подбугровую область на ядра блуждающего нерва и симпатических сердечных нервных волокон
Эфферентная парасимпатическая иннервация. Преганглионарные волокна начинаются в дорсальном вегетативном ядре блуждающего нерва и идут в составе последнего, его сердечных ветвей (rami cardiaci n. vagi) и сердечных сплетений (см. иннервацию сердца, с. 395) до внутренних узлов сердца (это впервые установил В. В. Николаев в 1893 г.), а также узлов околосердечных полей (Mitchell, 1957). Постганглионарные волокна исходят от этих узлов к мышце сердца. Функция: торможение и угнетение деятельности сердца. Сужение коронарных артерий.
И. Ф. Цион в 1866 г. открыл "сердечночувствующий" нерв, идущий в составе блуждающего нерва центростремительно. С этим нервом связано понижение кровяного давления, отчего он назван n. depressor.
Эфферентная симпатическая иннервация. Преганглионарные волокна начинаются из боковых рогов спинного мозга 4-5 верхних грудных сегментов, выходят в составе соответственных rami communicantes albi и проходят через симпатический ствол до пяти верхних грудных и трех шейных узлов. В этих узлах начинаются постганглионарные волокна, которые в составе сердечных нервов, nn. cardiaci, cervicales superior, medius et inferior и nn. cardiaci thoracici, достигают сердечной мышцы. По данным К. М. Быкова и др. (1955), перерыв осуществляется только в ganglion stellatum. По описанию Г. Ф. Иванова, сердечные нервы содержат в своем составе преганглионарные волокна, которые переключаются на постганглионарные в клетках сердечного сплетения. Функция: усиление работы сердца (это установил И. П. Павлов в 1888 г., назвав симпатический нерв усиливающим) и ускорение ритма (это впервые установил И. Ф. Цион в 1866 г.), расширение венечных сосудов.
Иннервация легких и бронхов. Афферентными путями от висцеральной плевры являются легочные ветви грудного отдела симпатического ствола, от париетальной плевры - nn. intercostales и n. phrenicus, от бронхов - n. vagus.
Эфферентная парасимпатическая иннервация. Преганглионарные волокна начинаются в дорсальном вегетативном ядре блуждающего нерва и идут в составе последнего и его легочных ветвей к узлам plexus pulmonalis, а также к узлам, расположенным по ходу трахеи, бронхов и внутри легких. Постганглионарные волокна направляются от этих узлов к мускулатуре и железам бронхиального дерева. Функция: сужение просвета бронхов и бронхиол и выделение слизи; расширение сосудов.
Эфферентная симпатическая иннервация. Преганглионарные волокна выходят из боковых рогов спинного мозга верхних грудных сегментов (Тh2-Тh6) и проходят через соответствующие rami communicantes albi и симпатический ствол к звездчатому и верхним грудным узлам. От последних начинаются постганглионарные волокна, которые проходят в составе легочного сплетения к бронхиальной мускулатуре и кровеносным сосудам. Функция: расширение просвета бронхов. Сужение и иногда расширение сосудов.
Иннервация желудочно-кишечного тракта (до сигмовидной кишки), поджелудочной железы, печени. Афферентные пути от указанных органов идут в составе n. vagus, n. splanchnicus major et minor, plexus hepaticus, plexus celiacus, грудных и поясничных спинномозговых нервов, а по данным Ф. П. Полякина (1958) и И. И. Шапиро (1961), и в составе n. phrenicus.
По симпатическим нервам передается чувство боли от этих органов, по n. vagus - другие афферентные импульсы, а от желудка - чувство тошноты и голода (Mitchell, 1953).
Эфферентная парасимпатическая иннервация. Преганглионарные волокна из дорсального вегетативного ядра блуждающего нерва проходят в составе последнего до терминальных узлов, находящихся в толще названных органов. В кишечнике - это клетки кишечных сплетений (plexus myentericus, submucosus). Постганглионарные волокна идут от этих узлов к гладким мышцам и железам. Функция: усиление перистальтики желудка, расслабление сфинктера привратника, усиление перистальтики кишок и желчного пузыря. По отношению к секреции в составе блуждающего нерва имеются волокна, возбуждающие и тормозящие ее. Расширение сосудов.
Эфферентная симпатическая иннервация. Преганглионарные волокна выходят из боковых рогов спинного мозга V-XII грудных сегментов, идут по соответствующим rami communicantes albi в симпатический ствол и далее без перерыва в составе nn. splanchnici majores (VI-IX) до промежуточных узлов, участвующих в образовании солнечного и нижнего брыжеечного сплетений (ganglia celiaca и ganglion mesentericum superius et inferius). Отсюда возникают постганглионарные волокна, идущие в составе plexus celiacus и pl. mesenterial superior к печени, pancreas, к тонкой кишке и к толстой до середины colon transversum; левая половина colon transversum и colon descendens иннервируются plexus mesenterial inferior. Указанные сплетения снабжают мускулатуру и железы названных органов. Функция: замедление перистальтики желудка, кишок и желчного пузыря, сужение просвета кровеносных сосудов и угнетение секреции желез.
К этому нужно прибавить, что задержка движений в желудке и кишечнике достигается также и тем, что симпатические нервы вызывают активное сокращение сфинктеров: sphincter pylori, сфинктеры кишечника и др.
Иннервация сигмовидной и прямой кишки и мочевого пузыря. Афферентные пути идут в составе plexus mesentericus inferior, plexus hypogastrics superior и inferior и в составе nn. splanchnici pelvini.
Эфферентная парасимпатическая иннервация. Преганглионарные волокна начинаются в боковых рогах спинного мозга II-IV крестцовых сегментов и выходят в составе соответствующих передних корешков спинномозговых нервов. Далее они идут в виде nn. splanchnici pelvini до внутриорганных узлов названных отделов толстой кишки и околоорганных узлов мочевого пузыря. В этих узлах начинаются постганглионарные волокна, которые достигают гладкой мускулатуры названных органов. Функция: возбуждение перистальтики сигмовидной и прямой кишок, расслабление m. sphincter ani internus, сокращение m. detrusor urinae и расслабление m. sphincter vesicae.
Эфферентная симпатическая иннервация. Преганглионарные волокна идут от боковых рогов поясничного отдела спинного мозга через соответствующие передние корешки в rami communicantes albi, проходят, не прерываясь, через симпатический ствол и достигают ganglion mesentericum inferius. Здесь начинаются постганглионарные волокна, идущие в составе nn. hypogastrici до гладкой мускулатуры названных органов. Функция: задержка перистальтики сигмовидной и прямой кишок и сокращение внутреннего сфинктера прямой кишки. В мочевом пузыре симпатические нервы вызывают расслабление m. detrusor urinae и сокращение сфинктера мочевого пузыря.
Иннервация половых органов: симпатическая, парасимпатическая. Иннервация других внутренних органов приводится после их описания.
Иннервация кровеносных сосудов. Степень иннервации артерий, капилляров и вен неодинакова (Mitchell, 1957). Артерии, у которых более развиты мышечные элементы в tunica media, получают более обильную иннервацию, вены - менее обильную; v. cava inferior и v. portae занимают промежуточное положение.
Более крупные сосуды, расположенные внутри полостей тела, получают иннервацию от ветвей симпатического ствола, ближайших сплетений вегетативной системы и прилежащих спинномозговых нервов; периферические же сосуды стенок полостей и сосуды конечностей получают иннервацию от проходящих поблизости нервов. Нервы, подходящие к сосудам, идут сегментарно и образуют периваскулярные сплетения, от которых отходят волокна, проникающие в стенку и распределяющиеся в адвентиции (tunica externa) и между последней и tunica media. Волокна снабжают мышечные образования стенки, имея различную форму окончаний. В настоящее время доказано наличие рецепторов во всех кровеносных и лимфатических сосудах.
Первый нейрон афферентного пути сосудистой системы лежит в межпозвонковых узлах или узлах вегетативных нервов (nn. splanchnici, n. vagus); далее он идет в составе кондуктора интероцептивного анализатора (с. 609). Сосудодвигательный центр лежит в продолговатом мозгу (Ф. В. Овсянников). К регуляции кровообращения имеют отношение globus pallidus, зрительный бугор, а также серый бугор. Высшие центры кровообращения, как и всех вегетативных функций, заложены в коре моторной зоны головного мозга (лобная доля), а также впереди и сзади нее. По новейшим данным, корковый конец анализатора сосудистых функций располагается, по-видимому, во всех отделах коры. Нисходящие связи головного мозга со стволовыми и спинальными центрами осуществляются, по-видимому, пирамидными и экстрапирамидными трактами.
Замыкание рефлекторной дуги может происходить на всех уровнях центральной нервной системы, а также в узлах вегетативных сплетений (собственная вегетативная рефлекторная дуга).
Эфферентный путь вызывает вазомоторный эффект - расширение или сужение сосудов. Сосудосуживающие волокна проходят в составе симпатических нервов, сосудорасширяющие волокна идут в составе всех парасимпатических нервов краниального отдела вегетативной системы (III, VII, IX, X), в составе задних корешков спинномозговых нервов (признается не всеми) и парасимпатических нервов сакрального отдела (nn. splanchnici pelvini).
При использовании материалов сайта активная ссылка обязательна:
http://anfiz.ru/ 'Анатомия и физиология человека'