Головной мозг
Общий обзор головного мозга
Головной мозг, encephalon, помещается в полости черепа и имеет форму, в общих чертах соответствующую внутренним очертаниям черепной полости. Его верхнебоковая, или дорсальная, поверхность сообразно своду черепа выпукла, а вентральная сторона, или основание мозга, более или менее уплощена и неровна. В головном мозгу можно различить три крупные части: полушария большого мозга (hemispheriae cerebri), мозжечок (cerebellum) и мозговой ствол (truncus cerebri). Наибольшую часть всего головного мозга занимают полушария, за ними по величине следует мозжечок, остальную, сравнительно небольшую часть составляет мозговой ствол.
Верхнелатеральная поверхность головного мозга. Оба полушария разделяются друг от друга щелью, fisstira longitudinalis cerebri, идущей в сагиттальном направлении. В глубине продольной щели полушария связаны между собой спайкой - мозолистым телом, corpus callosum, и другими лежащими под ним образованиями. Спереди от мозолистого тела продольная щель сквозная, а сзади она переходит в поперечную щель мозга, fissiira transversa cerebri, отделяющую задние части полушарий от лежащего под ними мозжечка.
Нижняя поверхность мозга (рис. 272). Со стороны нижней поверхности мозга, fades inferior cerebri, видна не только нижняя сторона полушарий большого мозга и мозжечка, но и вся нижняя поверхность мозгового ствола, а также отходящие от мозга нервы. Здесь мы встречаем следующие части, если идти спереди назад. Передний отдел нижней поверхности головного мозга представлен лобными долями полушарий. На нижней поверхности лобных долей замечаются обонятельные луковицы, btilbi olfactorii, к которым из полости носа через отверстия lamina cribrosa решетчатой кости подходят тонкие нервные нити, fila olfactoria, образующие в своей совокупности I пару головных нервов - обонятельные нервы, nn. olfactorii. Обыкновенно при вынимании мозга из черепа эти нити отрываются от billbus olfactorius. Обонятельные луковицы продолжаются кзади в обонятельные тракты, tractus olfactorii, оканчивающиеся каждый двумя корешками, между которыми находится возвышение, называемое trigonum olfactorium. Непосредственно сзади последнего на той и другой стороне находится переднее продырявленное вещество, substantia perforata anterior, названное так по причине находящихся здесь маленьких дырочек, через которые проходят в мозговое вещество сосуды.
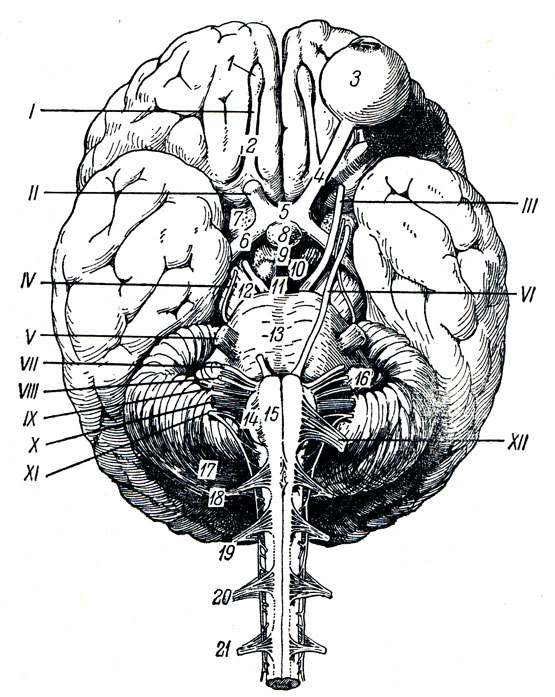
Рис. 272. Нижняя поверхность головного мозга. I - tracius olfactorius; II - n. opticus; III - noculomotorius; IV - n. trochlears; V - n. trigeminus; VI - n. abducens; VII - n. facialis; VIII - n. vestibulocochlearis; IX - n. glossopharyngeus X - n. vagus; XI - n. accessorius; XII - n. hypoglossus; 1 - bulbus olfactorius; 2 - tractus olfactorius; 3 - bulbus oculi sinistri; 4 - n opticus; 5 - chiasma opticum; 6 - tractus opticus; 7 - substantia perforata ant.; 8 - hypophysis; 9 - tuber cinereum; 10 - corpus mamillare; 11 - substantia perforata post.; 12 - pedunculus cerebri; 13 - pons; 14 - oliva; 15 - pyramis; 16 - flocculus; 17 - cerebellum; 18, 19, 20, 21 - корешки четырех верхних спинномозговых нервов
Посередине между обоими передними продырявленными пространствами лежит зрительный перекрест, chiasma opticum, имеющий форму буквы "X". От верхней поверхности хиазмы отходит тоненькая пластинка серого цвета, lamina terminalis, идущая вглубь fissura longitudinalis cerebri. Сзади зрительного перекреста помещается серый бугор, tuber cinereum; верхушка его вытянута в узкую трубку, так называемую воронку, infundibulum, на которой, как ягода на стебельке, висит кругловатое тело - придаток мозга, hypophysis cerebri. При положении мозга на его месте в черепе придаток помещается в углублении турецкого седла. Позади серого бугра находятся два шарообразных белого цвета возвышения - сосцевидные тела, corpora mamillaria. За ними лежит довольно глубокая четырехугольная ямка, fossa interpeduncularis, ограниченная с боков двумя толстыми белыми пучками, сходящимися кзади и называемыми ножками мозга, pedunculi cerebri. Дно ямки пронизано отверстиями для сосудов, а потому носит название заднего продырявленного вещества, substantia perforata posterior. Рядом с этим веществом в бороздке медиального края мозговой ножки на той и другой стороне выходит III пара - глазодвигательный нерв, n. oculomotorius. Сбоку ножек мозга виден самый тонкий из головных нервов блоковый нерв, n. trochlearis - IV пара, который, однако, отходит не на основании мозга, а с его дорсальной стороны, из так называемого верхнего мозгового паруса. Позади ножек мозга находится толстый поперечный вал - мост, pons (Varolii), который, суживаясь с боков, погружается в мозжечок. Боковые части моста, ближайшие к мозжечку, носят название средних ножек мозжечка, pedunculi cerebellares medii; на границе между ними и собственно мостом выходит на той и другой стороне V пара - тройничный нерв, n. trigeminus. Позади варолиева моста лежит продолговатый мозг, medulla oblongata; между ним и задним краем моста по бокам средней линии видно начало VI пары - отводящего нерва, n. abducens; еще далее вбок у заднего края средних ножек мозжечка выходят рядом на той и другой стороне еще два нерва: VII пара - лицевой нерв, n. facialis, и VIII пара - n. vestibulocochlearis.
Между пирамидой и оливой продолговатого мозга выходят корешки XII пары - подъязычного нерва, n. hypoglossus. Корешки IX, X и XI пар - n. glossopharyngeus, n. vagus и n. accessorius (верхней части) - выходят из бороздки позади оливы. Нижние волокна XI пары отходят уже от спинного мозга в шейной его части.
Развитие (эмбриогенез) головного мозга
Мозговая трубка очень рано подразделяется на два отдела, соответствующие головному и спинному мозгу. Передний, расширенный ее отдел, представляющий зачаток головного мозга, как было уже сказано, расчленяется путем перетяжек на три первичных мозговых пузырька, лежащих друг за другом: передний, prosencephalon, средний, mesencephalon, и задний, rhombencephalon. Передний мозговой пузырек замыкается спереди так называемой концевой пластинкой, lamina terminalis. Эта стадия из трех пузырьков при последующей дифференцировке превращается в пять пузырьков, дающих начало пяти главным отделам головного мозга (рис. 273). Одновременно с этим мозговая трубка изгибается в сагиттальном направлении. Прежде всего в области среднего пузырька развивается выпуклый в дорсальную сторону теменной изгиб, а затем на границе с зачатком спинного мозга также выпуклый дорсально затылочный изгиб. Между ними образуется в области заднего пузырька третий изгиб, выпуклый в вентральную сторону (мостовой изгиб).
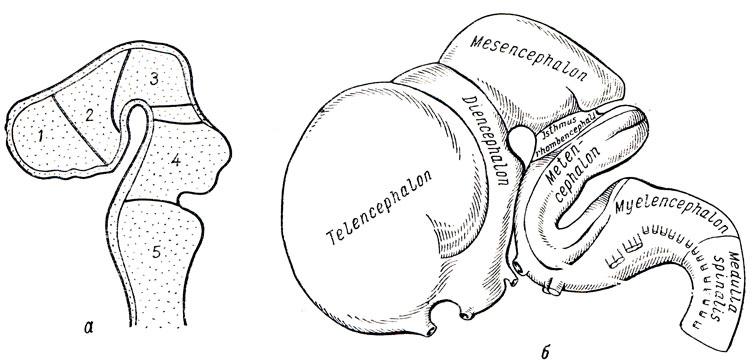
Рис. 273. Развитие головного мозга (схема). а - пять мозговых пузырей: 1 - первый пузырь (telencephalon - конечный мозг); 2 - второй пузырь (diencephalon - промежуточный мозг); 3 - третий пузырь (mesencephalon - средний мозг); 4 - четвертый пузырь (metencephalon - собственно задний мозг как часть ромбовидного мозга); 5 - пятый пузырь (myelencephalon - продолговатый мозг); между третьим и четвертым пузырем - перешеек (isthmus), б - развитие головного мозга (по Р. Д. Синельникову)
Посредством этого последнего изгиба задний мозговой пузырек, rhombencephalon, делится на два отдела. Из них задний, myelencephalon, превращается при окончательном развитии в продолговатый мозг, а из переднего отдела, называемого теfencephalon, развивается с вентральной стороны варолиев мост и с дорсальной стороны мозжечок. Metencephalon отделяется от лежащего впереди него пузырька среднего мозга узкой перетяжкой, isthmus rhombencephali. Общая полость rhombencephalon, имеющая на фронтальном сечении вид ромба, образует IV желудочек, сообщающийся с центральным каналом спинного мозга. Вентральная и боковые стенки ее благодаря развитию в них ядер головных нервов сильно утолщаются, дорсальная же стенка остается тонкой. В области продолговатого мозга большая часть ее состоит только из одного эпителиального слоя, срастающегося" с сосудистой мозговой оболочкой (tela chorioidea inferior). Стенки среднего пузырька, mesencephalon, утолщаются при развитии в них мозгового вещества более равномерно. Вентрально из них возникают ножки мозга, а с дорсальной стороны - пластинка четверохолмия (см. рис. 273). Полость среднего пузырька превращается в узкий канал - водопровод, соединяющийся с IV желудочком.
Более значительной дифференцировке и видоизменениям в форме подвергается передний мозговой пузырек, prosencephalon, который подразделяется на заднюю часть, diencephalon (промежуточный мозг), и переднюю, telencephalon (конечный мозг). Боковые стенки промежуточного мозга, утолщаясь, образуют зрительные бугры (thalami). Кроме того, боковые стенки, выпячиваясь в стороны, образуют два глазных пузырька, из которых впоследствии развиваются сетчатая оболочка глаз и зрительные нервы. Дорсальная стенка промежуточного мозга остается тонкой, в виде эпителиальной пластинки, срастающейся с сосудистой оболочкой (tela chorioidea superior). Сзади из этой стенки возникает выпячивание, за счет которого происходит эпифиз (corpus pineale). Полые ножки глазных пузырьков втягиваются с вентральной стороны в стенку переднего мозгового пузырька, вследствие чего на дне полости последнего образуется углубление, recessus opticus, передняя стенка которого состоит из тонкой lamina terminalis. Позади recessus opticus возникает другое воронкообразное углубление, стенки которого дают tuber cinereum, infundibulum и заднюю (нервную) долю hypophysis cerebri. Еще далее кзади в области diencephalon в виде одиночного возвышения закладываются парные corpora mamillaria. Полость промежуточного мозга образует III желудочек.
Telencephalon разделяется на среднюю, меньшую часть (telencephalon medium) и две большие боковые части - пузырьки полушарий большого мозга (hemispherium dextrum et sinistrum), которые у человека разрастаются очень сильно и в конце развития своей величиной значительно превосходят остальные отделы головного мозга. Полость telencephalon medium, являющаяся передним продолжением полости промежуточного мозга (III желудочка), по бокам сообщается посредством межжелудочковых отверстий с полостями пузырьков полушарий, которые на развитом мозге носят название боковых желудочков. Передняя стенка средней части конечного мозга (telencephalon medium), представляющая непосредственное продолжение lamina terminalis, в начале первого месяца эмбриональной жизни образует утолщение, так называемую комиссуральную пластинку, из которой впоследствии развиваются мозолистое тело и передняя комиссура.
В основании пузырьков полушарий внутри на той и другой стороне образуется выступ, так называемый узловой бугор, из которого развивается полосатое тело, corpus striatum. Часть медиальной стенки пузырька полушарий остается в форме одного эпителиального слоя, который вворачивается внутрь пузырька складкой сосудистой оболочки (plexus chorioideus). На нижней стороне каждого пузырька полушария уже на 5-й неделе эмбриональной жизни происходит выпячивание - зачаток обонятельного мозга, rhinencephalon, которое постепенно отграничивается от стенки полушарий бороздкой, соответствующей fissura rhinalis lateralis. При развитии серого вещества (коры), а затем и белого в стенках полушария последнее увеличивается и образует так называемый плащ, pallium, лежащий над обонятельным мозгом и покрывающий собой не только зрительные бугры, но и дорсальную поверхность среднего мозга и мозжечка.
Полушарие при своем росте увеличивается сначала в области лобной доли, затем теменной и затылочной и, наконец, височной. Благодаря этому создается впечатление, как будто плащ вращается вокруг зрительных бугров сначала спереди назад, затем вниз и, наконец, загибается вперед, к лобной доле. Вследствие этого на боковой поверхности полушария, между лобной долей и приблизившейся к ней височной, образуется яма, fossa cerebri lateralis (Sylvii), которая при полном сближении названных долей большого мозга превращается в щель, sulcus cerebri lateralis (Sylvii); на дне ее образуется островок, insula.
При развитии и росте полушария вместе с ним развиваются и совершают указанное "вращение" и его внутренние камеры, боковые желудочки мозга (остатки полости первичного пузыря), а также часть corpus striatum (хвостатое ядро), чем и объясняется сходство их формы с формой полушария: у желудочков - наличие передней, центральной и задней частей и загибающейся книзу и вперед нижней части (см. рис. 295), у хвостатого ядра - наличие головки, тела и загибающегося книзу и вперед хвоста.
Борозды и извилины (рис. 274, 275, 276) возникают вследствие неравномерного роста самого мозга (Д. Зернов), что связано с развитием отдельных его частей. Так, на месте обонятельного мозга возникает sulcus olfactorius, sulcus hyppocdmpi и sulcus cinguli; на границе корковых концов кожного и двигательного анализатора (понятие анализатора и описание борозд см. ниже) - sulcus centralis; на границе двигательного анализатора и премоторной зоны, получающей импульсы от внутренностей, - sulcus precentralis; на месте слухового анализатора - sulcus temporalis superior; в области зрительного анализатора - sulcus calcarinus и sulcus parieiooccipitdlis.
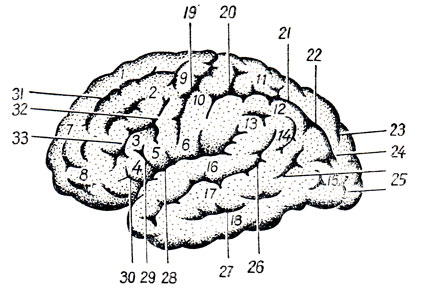
Рис. 274. Верхнелатеральная поверхность левого полушария головного мозга. 1, 7 - gyrus frontalis superior; 2 - gyrus frontalis medius: 3 - gyrus frontalis inferior; 4 - pars triangularis; 5 - pars opercularis; 8 - polus frontalis; 9 - gyrus precentralis; 10, 6 - gyrus postcentralis; 11 - lobulus parietalis superior; 12 - lobulus parietalis inferior; 13 - gyrus supramarginalis; 14 - gyrus angularis; 15 - gyri occipitales laterales; 16 - gyrus temporalis superior; 17 - gyrus temporalis medius; 18 - gyrus temporalis inferior; 19 - sulcus centralis; 20 - sulcus postcentralis; 21 - sulcus intraparietalis; 22 - соединение sulcus intraparietalis с sulcus occipitalis transversa; 23 - sulcus parietooccipitalis; 24 - sulcus occipitalis transversus; 25 - sulci occipitales laterales; 26 - sulcus temporalis superior; 27 - sulcus temporalis inferior; 28 - sulcus cerebri lateralis; 29 - ramus ascendens sulci cerebri lateralis; 30 - ramus anterior sulci cerebri lateralis; 31 - sulcus frontalis superior; 32 - sulcus precentralis; 33 - sulcus frontalis inferior
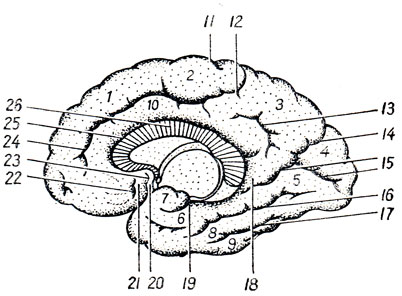
Рис. 275. Медиальная поверхность правого полушария головного мозга. 1 - gyrus frontalis superior; 2 - lobulus paracentralis; 3 - precuneus; 4 - cuneus; 5 - gyrus lingualis; 6 - gyrus parahippocampalis; 7 - uncus; 8 - gyrus occipitotemporal med.; 9 - gyrus occipitotmporalis lat.; 10 - gyrus cinguli; 11 - sulcus centralis; 12, 24 - sulcus cinguli; 13 - sulcus subparietalis; 14 - sulcus parietooccipitalis; 15 - sulcus calcarinus; 16 - sulcus collateralis; 17 - sulcus occipitotemporalis; 18 - isthmus gyri cinguli; 19 - sulcus hippocampi; 20 - gyrus subcallosus; 21 - area olfactoria; 22 - sulcus olfactorius anterior; 23 - sulcus olfactorius posterior; 25 - sulcus corporis callosi; 26 - corpus callosum
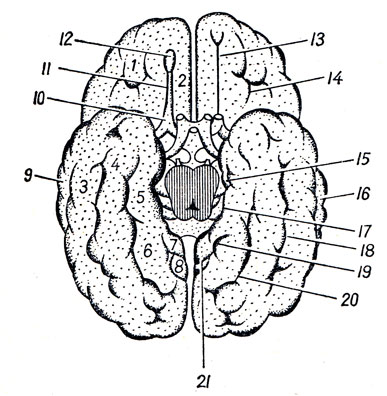
Рис. 276. Нижняя поверхность головного мозга. 1 - gyri orbit ales; 2 - gyrus rectus; 3, 4 - gyrus occipitotemporales medialis et lateralis; 5 - gyrus parahippocampalis; 6 - gyrus occipitotemporalis medialis; 7 - isthmus gyri cinguli; 8 - cuneus; 9 - gyrus temporalis medius; 10 - trigonum olfactorium; 11 - tractus olfactorius; 12 - bulbus olfactorius; 13 - sulcus olfactorius; 14 - sulci orbitales; 15 - uncus gyri parahippocampalis; 16 - sulcus temporalis inferior; 17 - sulcus hippocampi; 18 - sulcus occipitotemporalis; 19 - sulcus calcarinus; 20 - sulcus collateralis; 21 - sulcus parietooccipitalis
Все эти борозды, появляющиеся раньше других и отличающиеся абсолютным постоянством, относятся, по Д. Зернову, к бороздам первой категории. Остальные борозды, имеющие наименования и также возникающие в связи с развитием анализаторов, но появляющиеся несколько позднее и отличающиеся меньшим постоянством, относятся к бороздам второй категории. К моменту рождения имеются все борозды первой и второй категорий. Наконец, многочисленные мелкие бороздки, не имеющие названий, появляются не только в утробной жизни, но и после рождения. Они крайне непостоянны по времени появления, месту и числу; это борозды третьей категории. От степени их развития зависит все разнообразие и сложность мозгового рельефа. Рост человеческого мозга в эмбриональном периоде и в первые годы жизни, пока идет бурный рост организма, его приспособление к новой среде, приобретение способности к прямохождению и становление второй, словесной, сигнальной системы, происходит очень интенсивно и заканчивается к 20 годам. У новорожденных мозг (в среднем) имеет вес 340 г у мальчиков и 330 г у девочек, а у взрослого - 1375 г у мужчин и 1245 г у женщин.
Отдельные части головного мозга
На основании эмбрионального развития, как было уже указано, головной мозг делится на отделы, располагающиеся, начиная с каудального конца, в таком порядке:
1) rhombencephalon, ромбовидный, или задний, мозг, который, в свою очередь, состоит из: a) myelencephalon, продолговатого мозга и б) metencephalon - собственно заднего мозга; 2) mesencephalon - средний мозг; 3) prosencephalon, передний мозг, в котором различают: a) diencephalon - промежуточный мозг и б) telencephalon - конечный мозг.
Кроме этих отделов, выделяют еще перешеек, isthmus rhombencephali, между rhombencephalon и средним мозгом.
Mesencephalon et prosencephalon составляют большой мозг, cerebrum, в отличие от малого мозга, мозжечка - cerebellum.
Rhombencephalon
Myelencephalon, продолговатый мозг
Продолговатый мозг, tnyelencephalon, medulla oblongdta (рис. 277, 278), представляет непосредственное продолжение спинного мозга в ствол головного мозга и является частью ромбовидного, или заднего, мозга. Он сочетает в себе черты строения спинного мозга и начального отдела головного, чем и оправдывается его название - myelencephalon. Medulla oblongata имеет вид луковицы, bulbus cerebri, s. bulbus medullae spinalis (отсюда термин "бульварные расстройства"); верхний расширенный конец граничит с мостом, а нижней ее границей служит место выхода корешков I пары шейных нервов или уровень большого отверстия затылочной кости.
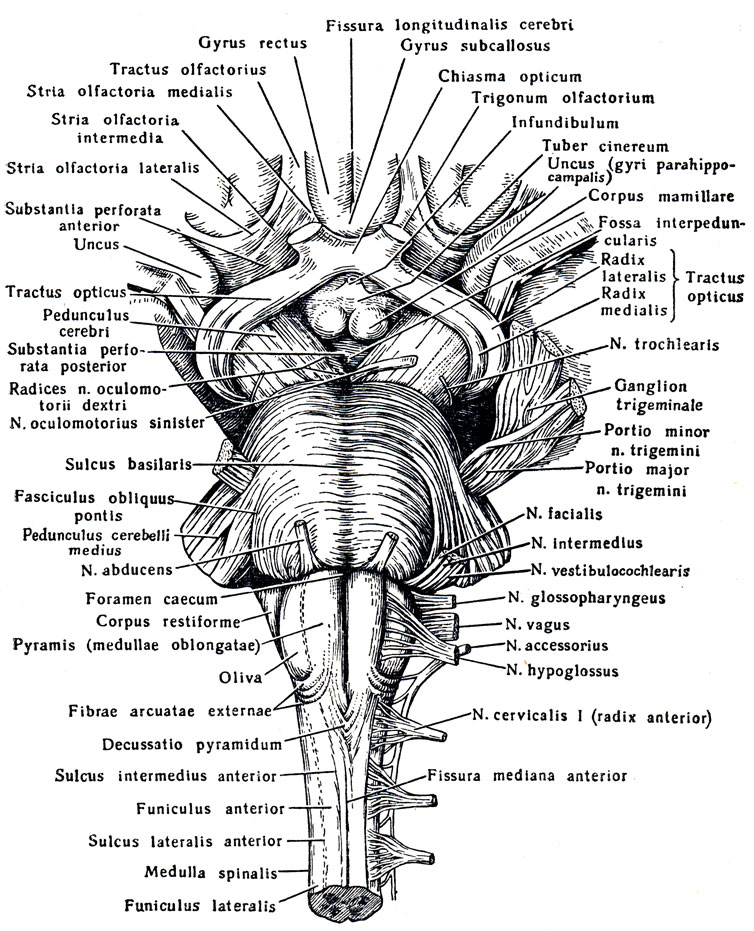
Рис. 277. Вентральная сторона продолговатого мозга, моста и среднего мозга
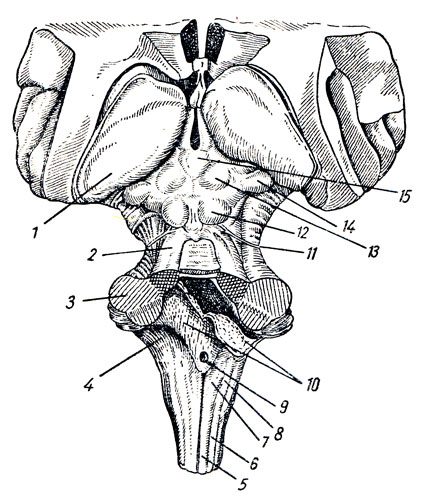
Рис. 278. Ствол головного мозга. 1 - pulvinar (задняя часть thalamus); 2 - pedunculus cerebellaris superior; 3 - pedunculus cerebellaris medius; 4 - pedunculus cerebellaris inferior; 5 - fasciculus gracilis (Goll); 6 - fasciculus cuneatus (Burdach); 7 - tuberculum nuclei gracilis; 8 - tuberculum nuclei cuneati; 9 - apertura mediana ventriculi quarti; 10 - plexus chorioideus, tela chorioidea ventriculi quarti (разрезаны и отвернуты, через разрез видна полость IV желудочка); 11 - n. trochlears; 12 - colliculus inferior четверохолмия; 13 - colliculus superior четверохолмия; 14 - corpus geniculatum mediale; 15 - corpus pineale
1. На передней (вентральной) поверхности продолговатого мозга по средней линии проходит fissura mediana anterior, составляющая продолжение одноименной борозды спинного мозга. По бокам ее на той и другой стороне находятся два продольных тяжа - пирамиды, pyramides medullae oblongatae, которые как бы продолжаются в передние канатики спинного мозга. Составляющие пирамиды пучки нервных волокон частью перекрещиваются в глубине fissura mediana anterior с аналогичными волокнами противоположной стороны - decussdtio pyramidum, после чего спускаются в боковом канатике на другой стороне спинного мозга - tractus corticospinal (pyramidalis) lateralis, частью остаются неперекрещенными и спускаются в переднем канатике спинного мозга на своей стороне - tractus corticospinal (pyramidalis) anterior.
Пирамиды отсутствуют у низших позвоночных и появляются по мере развития новой коры; поэтому они наиболее развиты у человека, так как пирамидные волокна соединяют кору большого мозга, достигшую у человека наивысшего развития, с ядрами головных нервов и передними рогами спинного мозга.
Латерально от пирамиды лежит овальное возвышение - олива, olfva, которая отделена от пирамиды бороздкой, sulcus lateralis anterior.
2. На дорсальной (задней) стороне продолговатого мозга (см. рис. 278) тянется sulcus medidnus posterior - непосредственное продолжение одноименной борозды спинного мозга. По бокам ее лежат задние канатики, ограниченные латерально с той и другой стороны слабо выраженной sulcus lateralis posterior. По направлению кверху задние канатики расходятся в стороны и идут к мозжечку, входя в состав его нижних ножек, pedunculi cerebellares inferiores, окаймляющих снизу ромбовидную ямку. Каждый задний канатик подразделяется при помощи промежуточной борозды - на медиальный, fasciculus gracilis, и латеральный, fasciculus cuneatus. У нижнего угла ромбовидной ямки нежный и клиновидный канатики приобретают утолщения - tuberculum nuclei gracilis и tuberculum nuclei cunedti. Эти утолщения обусловлены соименными с канатиками ядрами серого вещества, nucleus gracilis и nucleus cuneatus. В названных ядрах оканчиваются проходящие в задних канатиках восходящие волокна задних корешков спинного мозга (пучки Голля и Бурдаха). Латеральная поверхность продолговатого мозга, находящаяся между sulcus lateralis posterior et anterior, соответствует боковому канатику. Из sulcus lateralis posterior позади оливы выходят XI, X и IX пары головных нервов. В состав продолговатого мозга входит нижняя часть ромбовидной ямки (рис. 279; см. рис. 283).
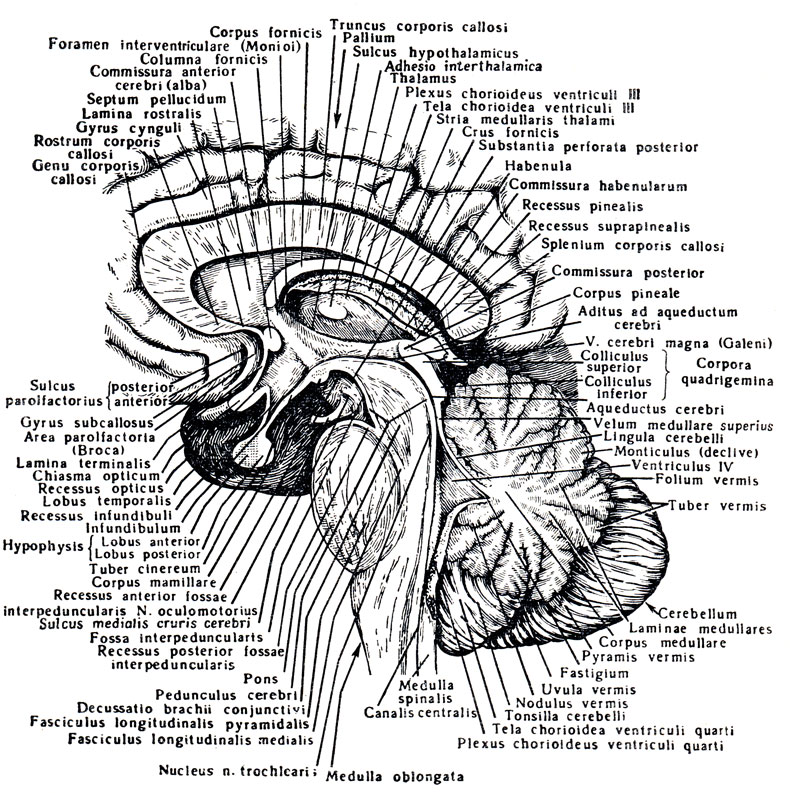
Рис. 279. Ствол головного мозга (сагиттальный разрез)
Внутреннее строение продолговатого мозга. Продолговатый мозг возник в связи с развитием органов статики и акустики, а также в связи с жаберным аппаратом, имеющим отношение к дыханию и кровообращению. Поэтому в нем заложены ядра серого вещества, имеющие отношение к равновесию и координации движений, а также к регуляции обмена веществ (рис. 280).
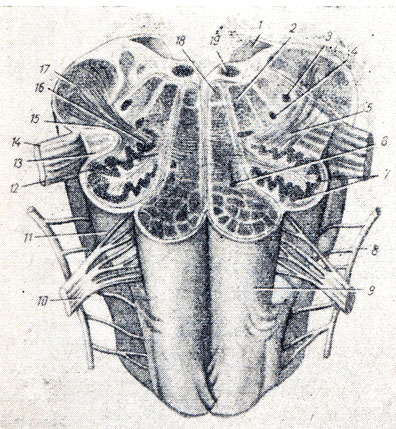
Рис. 280. Продолговатый мозг - горизонтальный разрез на уровне оливы (по Р. Д. Синельникову). 1 - velum medullare posterior; 2 - formatio reticularis; 3 - nucleus tractus spinalis n. trigemini; 4 - nucleus ambiguus; 5 - tractus olivocerebellaris; 6 - nucleus olivaris accessorius medialis; 7, 16 - nucleus olivaris; 8 - n. accessorius; 9 - pyramis; 10 - n. hypoglossus; 11 - oliva; 12 - hilus nuclei olivaris 13 - tractus tectospinal; 14 - n. vagus; 15 - tractus rubrospinalis; 17 - pedunculus cerebellaris inferior; 18 - fasciculus longitudinalis medialis; 19 - nucleus n. hypogllossi
1. Nucleus olivdris, ядро оливы, имеет вид извитой пластинки серого вещества, открытой медиально (hilus), и обусловливает снаружи выпячивание оливы. Оно связано с зубчатым ядром мозжечка и является промежуточным ядром равновесия, наиболее выраженным у человека, вертикальное положение которого нуждается в наиболее совершенном аппарате равновесия. (Встречается еще nucl. olivaris access, med.)
2. Formatio reticularis, сетчатая формация, образующаяся из переплетения нервных волокон и лежащих между ними нервных клеток.
3. Ядра четырех пар нижних головных нервов (XII-IX), имеющие отношение к иннервации производных жаберного аппарата и внутренностей.
4. Жизненно важные центры дыхания и кровообращения, связанные с ядрами блуждающего нерва. Поэтому при повреждении продолговатого мозга может наступить смерть.
Белое вещество продолговатого мозга содержит длинные и короткие волокна. К длинным относятся проходящие транзитно в передние канатики спинного мозга нисходящие пирамидные пути, частью перекрещивающиеся в области пирамид. Кроме того, от ядер задних канатиков (nucleus gracilis et cuneatus) начинаются вторые нейроны восходящих чувствительных путей, идущих от продолговатого мозга к зрительному бугру, trdctus bulboihaldmicus. Волокна этого пучка образуют медиальную петлю, lemniscus medidlis, которая в продолговатом мозгу совершает перекрест, decussdtio lemniscorum, и в виде пучка волокон, расположенных дорсальнее пирамид, между оливами - межоливный петлевой слой - идет далее. Таким образом, в продолговатом мозгу имеются два перекреста длинных проводящих путей: вентральный двигательный, decussatio pyramidum, и дорсальный чувствительный, decussatio lemniscorum.
К коротким путям относятся пучки нервных волокон, соединяющие между собой отдельные ядра серого вещества, а также ядра продолговатого мозга с соседними отделами головного мозга. Среди них следует отметить trdctus olivocerebellars и лежащий дорсально от межоливного слоя fasciculus longitudindlis medidlis.
Топографические взаимоотношения главнейших образований продолговатого мозга видны на поперечном срезе, проведенном на уровне олив (см. рис. 280). Отходящие от ядер подъязычного и блуждающего нервов корешки делят продолговатый мозг на той и другой стороне на три области: заднюю, боковую и переднюю. В задней лежат ядра заднего канатика и нижние ножки мозжечка, в боковой - ядро оливы и formatio reticularis и в передней - пирамиды.
Metencephalon
Metencephalon состоит из двух частей: вентральной - моста и дорсальной - мозжечка.
Мост
Мост, pons (Varolii), представляет собой со стороны основания мозга толстый белый вал, граничащий каудально с верхним концом продолговатого мозга, а краниально - с ножками мозга (см. рис. 277). Латеральной границей моста служит искусственно проводимая линия через корешки тройничного и лицевого нервов, linea trigeminofacidlis. Латерально от этой линии находятся средние ножки мозжечка, pedunculi cerebellares medii, погружающиеся на той и другой стороне в мозжечок. Дорсальная поверхность моста не видна снаружи, так как она скрыта под мозжечком, образуя верхнюю часть ромбовидной ямки (дна IV желудочка). Вентральная поверхность моста имеет волокнистый характер, причем волокна в общем идут поперечно и направляются в pedunculi cerebellares medii. По средней линии вентральной поверхности проходит пологая канавка, sulcus basilaris, в которой лежит a. basilaris.
Внутреннее строение моста. На поперечных разрезах моста можно видеть, что он состоит из большей нижней, или вентральной, части, pars ventralis pontis, и меньшей дорсальной, pars dorsalis pontis. Границей между ними служит толстый слой поперечных волокон - трапециевидное тело, corpus trapezoideum, волокна которого относятся к слуховому пути. В области трапециевидного тела располагается ядро, также имеющее отношение к слуховому пути - nucleus dorsalis corporis trapezoidei, или nucleus olivaris superior (BNA). Последнее название было дано ядру потому, что оно имеет зубчатую форму подобно ядру оливы продолговатого мозга.
Pars ventralis содержит продольные и поперечные волокна, между которыми разбросаны собственные ядра серого вещества, nuclei pontis.
Продольные волокна принадлежат к пирамидным путям, fibrae corticopontinae, которые связаны с собственными ядрами моста, откуда берут начало поперечные волокна, идущие к коре мозжечка, tractus pontocerebellaris. Вся эта система проводящих путей связывает через посредство моста кору полушарий большого мозга с корой полушарий мозжечка. Чем сильнее развита кора большого мозга, тем сильнее развиты мост и мозжечок. Естественно, что мост оказывается наиболее выраженным у человека, что является специфической чертой строения его головного мозга. В pars dorsalis находится formatio reticularis pontis, являющееся продолжением такой же части продолговатого мозга, а поверх сетчатой формации - выстланное эпендимой дно ромбовидной ямки с лежащими под ним ядрами головных нервов (VIII-V пары).
В pars dorsalis продолжаются также проводящие пути продолговатого мозга, располагающиеся между средней линией и nucleus dorsalis corporis trapezoidei и входящие в состав медиальной петли, lemniscus medialis; в последней перекрещиваются восходящие пути продолговатого мозга, tractus bulbothalamicus.
Мозжечок
Мозжечок, cerebellum, является производным заднего мозга, развившегося в связи с рецепторами статики. Поэтому он имеет прямое отношение к координации движений и является органом приспособления организма к преодолению основных свойств массы тела - тяжести и инерции. Его считают также одним из высших центров вегетативной (симпатической) нервной системы (Л. А. Орбели и его школа).
Развитие мозжечка в процессе филогенеза прошло 3 основных этапа соответственно изменению способов передвижения животного.
На первом этапе, когда животные передвигались в воде с помощью змеевидных движений тела (круглоротые), их мозжечок имел вид небольшого образования, auricula, которое у наземных животных развилось в так называемый клочок, flocculus. Это самая древняя часть мозжечка - archicerebellum. На втором этапе, когда у водных животных появляются плавники (рыбы) и меняются туловищные движения, возникает новая часть мозжечка - червь, vermis, которая у человека является старой частью мозжечка, paleocerebellum. Наконец, на третьем этапе у наземных животных возникают полушария мозжечка, которые являются самой новой частью мозжечка, neocerebellum, и постепенно разрастаются по мере отдаления тела животного от земли и развития передвижения при помощи конечностей. У человека в связи с прямохождением при помощи одной пары конечностей (ног) и усовершенствованием хватательных движений руки при трудовых процессах полушария мозжечка достигают наибольшего развития, так что мозжечок у человека развит сильнее, чем у всех животных, что составляет специфическую человеческую черту его строения.
Мозжечок помещается под затылочными долями полушарий большого мозга, дорсально от моста и продолговатого мозга, и лежит в задней черепной ямке. В нем различают объемистые боковые части, или полушария, hemispheria cerebelli, и расположенную между ними среднюю узкую часть - червь, vermis.
На переднем краю мозжечка находится передняя вырезка, которая охватывает прилежащую часть ствола мозга. На заднем краю имеется более узкая задняя вырезка, отделяющая полушария друг от друга.
Поверхность мозжечка покрыта слоем серого вещества, составляющим кору мозжечка, и образует узкие извилины, folia cerebelli, отделенные друг от друга бороздами, fissurae cerebelli. Среди них самая глубокая, fissura horizontdlis cerebelli, проходит по заднему краю мозжечка и отделяет верхнюю поверхность полушарий, fdcies superior, от нижней, fdcies inferior. С помощью горизонтальной и других крупных борозд вся поверхность мозжечка делится на ряд долек, lobuli cerebelli. Среди них необходимо выделить наиболее изолированную маленькую дольку - клочок, flocculus, лежащую на нижней поверхности каждого полушария у средней ножки мозжечка, а также связанную с клочком часть червя - nodulus, узелок. Flocculus соединен с nodulus посредством тонкой полоски - ножки клочка, pedunculus flocculi, которая медиально переходит в тонкую полулунную пластинку - нижний мозговой парус, velum medullare inferius.
Внутреннее строение мозжечка. В толще мозжечка имеются парные ядра серого вещества, заложенные в каждой половине мозжечка, среди белого ее вещества (рис. 281). По бокам от средней линии в области, где в мозжечок вдается шатер, fastigium, лежит самое медиальное ядро - ядро шатра, nucleus fasiigii. Латеральнее от него расположены маленькие островки шаровидного ядра, nucleus globosus, а еще латеральнее - пробковидное ядро, nucleus emboliformis. Наконец, в центре полушария находится зубчатое ядро, nucleus dentdtus, имеющее вид серой извилистой пластинки, похожей на ядро оливы, и открытое медиально - hilus nuclei dentati. Сходство nucleus dentatus мозжечка с имеющим также зубчатую форму ядром оливы не случайно, так как оба ядра связаны проводящими путями, fibrae olivocerebellares, и каждая извилина одного ядра аналогична извилине другого. Таким образом, оба ядра вместе участвуют в осуществлении функции равновесия (см. рис. 280, 281).
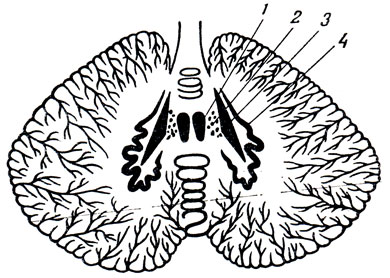
Рис. 281. Ядра мозжечка (схема). 1 - nucleus fastigii; 2 - nucleus globosus; 3 - nucleus emboliformis; 4 - nucleus dentatus
Названные ядра мозжечка имеют различный филогенетический возраст: nucleus fastigii относится к самой древней части мозжечка - flocculus (archicerebellum), связанной с вестибулярным аппаратом, nuclei emboliformis et globosus - к старой части (paleocerebellum), возникшей в связи с движениями туловища, и nucleus dentatus - к самой молодой (neocerebellum), развившейся в связи с передвижением при помощи конечностей. Поэтому при поражении каждой из этих частей нарушаются различные стороны двигательной функции, соответствующие различным стадиям филогенеза, а именно: при повреждении флоккуло-нодулярной системы и ее ядра шатра нарушается равновесие тела. При поражении червячка и соответствующих ему пробковидного и шаровидного ядер нарушается работа мускулатуры шеи и туловища. При поражении полушарий и зубчатого ядра- работа мускулатуры конечностей.
Белое вещество мозжечка на разрезе имеет вид мелких листочков растения, соответствующих каждой извилине, покрытой с периферии корой серого вещества. В результате общая картина белого и серого вещества на разрезе мозжечка напоминает дерево, arbor viiae cerebelli (древо жизни; название дано по внешнему виду, поскольку повреждение мозжечка не является непосредственной угрозой жизни). Белое вещество мозжечка слагается из различного рода нервных волокон. Одни из них связывают извилины и дольки, другие идут от коры к внутренним ядрам мозжечка и, наконец, третьи связывают мозжечок с соседними отделами мозга. Эти последние волокна идут в составе трех пар ножек мозжечка:
1. Нижние ножки, pedunculi cerebellares inferiores (к продолговатому мозгу).
В их составе идут к мозжечку trdctus spinocerebelldris posterior (Flechsig), fibrae arcudtae externae - от ядер задних канатиков продолговатого мозга, и fibrae olivocerebellars - от оливы. Все эти волокна оканчиваются в коре червячка и полушарий. Кроме того, здесь идут волокна от ядер вестибулярного нерва, заканчивающиеся в nucleus fastigii. Благодаря всем этим волокнам мозжечок получает импульсы от вестибулярного аппарата и проприоцептивного поля, вследствие чего становится ядром проприоцептивной чувствительности, совершающим автоматическую поправку на двигательную деятельность остальных отделов мозга. В составе нижних ножек идут также нисходящие пути в обратном направлении, а именно: от nucleus fastigii к латеральному вестибулярному ядру, а от него - к передним рогам спинного мозга, tractus vestibulospinalis. При посредстве этого пути мозжечок оказывает влияние на спинной мозг.
2. Средние ножки, pedunculi cerebellares medii (к мосту).
В их составе идут нервные волокна от ядер моста к коре мозжечка. Возникающие в ядрах моста проводящие пути к коре мозжечка, trdctus pontocerebellars, находятся на продолжении корково-мостовых путей, fibrae corticopontinae, оканчивающихся в ядрах моста после перекреста. Эти пути связывают кору большого мозга с корой мозжечка, чем и объясняется тот факт, что чем более развита кора большого мозга, тем более развиты мост и полушария мозжечка, что наблюдается у человека.
3. Верхние ножки, pedunculi cerebellares superiores (к четверохолмию). Они состоят из нервных волокон, идущих в обоих направлениях: 1) к мозжечку - trdctus spinocerebelldris anterior (Gowers), и 2) от nucleus dentatus мозжечка к крыше четверохолмия - trdctus cerebellotegmentdlis, который после перекреста заканчивается в красном ядре и в зрительном бугре. Через первые пути мозжечок получает импульсы от спинного мозга, а по вторым он посылает импульсы в экстрапирамидную систему, через которую сам влияет на спинной мозг.
Перешеек
Перешеек, isthmus rhombencephali, представляет переход от rhombencephalon к mesencephalon. В его состав входят: 1) верхние ножки мозжечка, pedunculi cerebellares superiores; 2) натянутый между ними и мозжечком верхний мозговой парус, velum medullare superius, который прикрепляется к срединной бороздке между бугорками четверохолмия; 3) треугольник петли, trigonum lemnisci, обусловленный ходом слуховых волокон латеральной петли, lemniscus lateralis. Этот треугольник серого цвета ограничен спереди нижней ручкой четверохолмия, сзади - верхней ножкой мозжечка и латерально - ножкой мозга. Последняя отделена от перешейка и среднего мозга ясно выраженной бороздой, sulcus lateralis mesencephali. Внутрь перешейка вдается верхний конец IV желудочка, переходящий в среднем мозгу в водопровод.
IV желудочек
IV желудочек, ventricuius quartus, представляет собой остаток полости заднего мозгового пузыря и поэтому является общей полостью для всех отделов заднего мозга, составляющих rhombencephalon (продолговатый мозг, мозжечок, мост и перешеек). IV желудочек напоминает палатку, в которой различают дно и крышу (см. рис. 279; рис. 282).
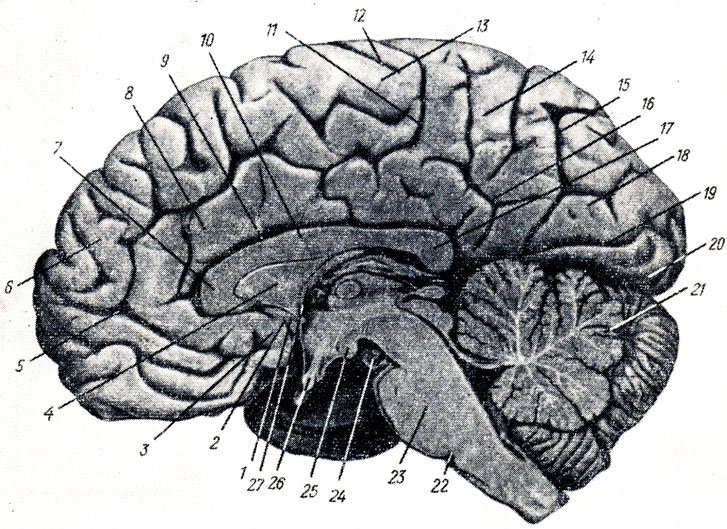
Рис. 282. Внутренняя поверхность правого полушария мозга, стволовой части и мозжечка. Водопровод мозга; IV желудочек. 1 - gyrus subcallosus; 2 - sulcus olfactorius posterior; 3 - area olfactoria; 4 - septum pellucidum; 5, 11 - sulcus cinguli; 6 - gyrus frontalis superior; 7 - genu corporis callosi; 8 - gyrus cinguli; 9 - sulcus corporis callosi; 10 - truncus corporis callosi; 12 - sulcus centralis (Rolandi); 13 - lobulus paracentralis; 14 - precuneus; 15 - sulcus parietooccipitalis; 16 - sulcus subparietalis; 17 - splenium corporis callosi; 18 - cuneus; 19 - sulcus calcarinus; 20 - gyrus occipitotemporal medialis; 21 - долька нижнего червя; 22 - задняя граница моста; 23 - pons; 24 - n. oculomotorius; 25 - corpus mamillare; 26 - chiasma opticum; 27 - fornix (columna)
Дно, или основание, желудочка имеет форму ромба, как бы вдавленного в заднюю поверхность продолговатого мозга и моста. Поэтому его называют ромбовидной ямкой, fossa rhomboidea. В задненижний угол ромбовидной ямки открывается центральный канал спинного мозга, а в передневерхнем углу IV желудочек сообщается с водопроводом. Латеральные углы заканчиваются слепо в виде двух карманов, recessus laterdles ventriculi qudrti, загибающихся вентрально вокруг нижних ножек мозжечка.
Крыша IV желудочка, tegmen ventriculi quarti, имеет форму шатра, fastigium (BNA), и составлена двумя мозговыми парусами: верхним, velum medullare superius, натянутым между верхними ножками мозжечка, и нижним, velum medullare inferius, парным образованием, примыкающим к ножкам клочка. Часть крыши между парусами образована веществом мозжечка. Нижний мозговой парус дополняется листком сосудистой мозговой оболочки, tela chorioidea ventriculi qudrti, покрытой изнутри слоем эпителия, lamina chorioidea epithelialis, представляющим рудимент задней стенки заднего мозгового пузыря (с ней связано сплетение - pl. chorioid. ventr. IV).
Tela chorioidea первоначально вполне замыкает полость желудочка, но затем в процессе развития в ней появляются три отверстия: одно - в области нижнего угла ромбовидной ямки, apertura medidna ventriculi qudrti (foramen Magendi) (самое большое), и два - в области боковых карманов желудочка, aperturae laterdles ventriculi qudrti (foramina Luschka). При посредстве этих отверстий IV желудочек сообщается с подпаутинным пространством головного мозга, благодаря чему спинномозговая жидкость поступает из мозговых желудочков в межоболочечные пространства. В случае сужения или заращения этих отверстий на почве воспаления мозговых оболочек (менингита) накапливающаяся в мозговых желудочках спинномозговая жидкость не находит себе выхода в подпаутинное пространство, и возникает водянка головного мозга.
Ромбовидная ямка, fossa rhomboidea (рис. 283), имеет соответственно ромбовидной форме четыре стороны - две верхние и две нижние. Верхние стороны ромба ограничены двумя верхними ножками мозжечка, а нижние стороны - двумя нижними ножками. Вдоль ромба, по средней линии, от верхнего угла к нижнему тянется срединная борозда, sulcus medidnus, которая делит ромбовидную ямку на правую и левую половины. По сторонам борозды расположено парное возвышение, eminentia medialis, обусловленное скоплением серого вещества.
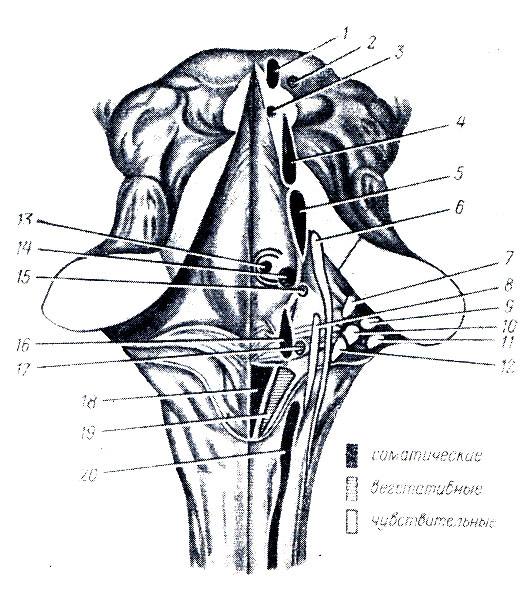
Рис. 283. Ядра головных нервов в ромбовидной ямке. 1 - nucleus n. oculomotorii; 2 - nucleus accessorius n. oculornotorii; 3 - nucleus n. trochlearis; 4 - nucleus tractus mesencephali n. trigemini; 5 - nucleus motorius n. trigemini; 6 - nucleus sensorius principalis n. trigemini; 7 - nucleus vestibularis superior; 8 - nucleus tr. solitarii (X, IX); 9 - nucleus cochlearis ventralis; 10 - nucleus vestibularis lateralis; 11 - nucleus cochlearis dorsalis; 12 - nucleus ves tibularis medialis; 13 - nucleus n. abducentis; 14 - nucleus n. facialis; 15 - nucleus salivatorius superior; 16 - nucleus ambiguus; 17 - nucleus salivatorius inferior; 18 - nucleus n. hypoglossi; 19 - nucleus dorsalis n. vagi; 20 - nucleus n. accessorii
Книзу eminentia medialis постепенно суживается, переходя в треугольник, на который проецируется ядро подъязычного нерва, trigonum nervi hypoglossi. Латеральнее нижней части этого треугольника лежит меньший треугольник, заметный по своей серой окраске, trigonum n. vagi, в котором заложено вегетативное ядро блуждающего нерва, nucleus dorsdlis nervi vagi. Вверху eminentia medialis имеет возвышение - лицевой бугорок, collicuius facialis, обусловленный прохождением корешка лицевого и проекцией ядра отводящего нервов.
В области латеральных углов располагается с обеих сторон вестибулярное поле, area vestibularis. Здесь помещаются ядра VIII пары. Часть выходящих из них волокон идет поперек ромбовидной ямки от латеральных углов к срединной борозде в виде горизонтальных полосок, striae medulldres ventriculi qudrti. Эти полоски делят ромбовидную ямку на верхнюю и нижнюю половины и соответствуют границе между продолговатым мозгом и мостом.
Топография серого вещества ромбовидной ямки. Серое вещество спинного мозга непосредственно переходит в серое вещество мозгового ствола и частью расстилается по ромбовидной ямке и стенкам водопровода (см. средний мозг), а частью разбивается на отдельные ядра головных нервов или ядра пучков проводящих путей.
Чтобы понять расположение этих ядер, нужно учитывать, как уже говорилось выше, что замкнутая мозговая трубка спинного мозга при переходе в продолговатый мозг раскрылась на своей задней стороне и развернулась в ромбовидную ямку. Вследствие этого задние рога серого вещества спинного мозга как бы разошлись в стороны. Заложенные в задних рогах соматически-чувствительные ядра расположились в ромбовидной ямке латерально, а соответствующие передним рогам соматически-двигательные ядра остались лежать медиально. Что касается вегетативных ядер, заложенных в боковых рогах спинного мозга, то соответственно положению боковых рогов между задними и передними эти ядра при развертывании мозговой трубки оказались лежащими в ромбовидной ямке между соматически-чувствительными и соматически-двигательными ядрами. В результате в стенке ромбовидной ямки в отличие от спинного мозга ядра серого вещества расположены не в передне-заднем направлении, а лежат рядами - медиально и латерально.
Так, например, соматически-двигательные ядра XII и VI пар лежат в медиальном ряду, вегетативные ядра X, IX, VII пар - в среднем ряду и соматически-чувствительные ядра VIII пары - латерально.
Проекция ядер головных нервов на ромбовидную ямку:
XII пара - подъязычный нерв, n. hypoglossus, имеет единственное двигательное ядро, заложенное в самой нижней части ромбовидной ямки, в глубине trigonum n. hypoglossi.
XI пара - добавочный нерв, n. accessorius, имеет два ядра (оба двигательные): одно заложено в спинном мозгу и называется nucleus spinalis n. accessorii, другое является каудальным продолжением ядер X и IX пар нервов и называется nucleus ambiguus. Оно лежит в продолговатом мозгу дорсо-латерально от ядра оливы.
X пара - блуждающий нерв, n. vagus, имеет три ядра:
1. Чувствительное ядро, nucleus trdctus solitdrii, расположено рядом с ядром подъязычного нерва, в глубине trigonum n. vagi.
2. Вегетативное ядро, nucleus dorsdlis я. vagi, лежит в той же области.
3. Двигательное ядро, nucleus ambiguus (двойное) общее с ядром IX пары, заложено в formatio reticularis, глубже nucleus dorsalis.
IX пара - языкоглоточный нерв, n. glossopharyngeus, также содержит три ядра:
1. Чувствительное ядро, nucleus trdctus solitdrii, лежит латеральнее ядра подъязычного нерва.
2. Вегетативное (секреторное) ядро, nucleus salivatorius inferior, нижнее слюноотделительное ядро; клетки его рассеяны в formatio reticularis продолговатого мозга между n. ambiguus и ядром оливы.
3. Двигательное ядро, общее с n. vagus и n. accessorius, nucleus ambiguus.
VIII пара - статически-слуховой нерв, n. vestibulocochlearis, имеет множественные ядра, проецирующиеся на латеральные углы ромбовидной ямки, в области area vestibularis. Ядра делятся на две группы соответственно двум частям нерва; одна часть нерва, pars cochlearis - нерв улитки, или собственно слуховой нерв, имеет два ядра: дорсальное, nucleus cochleans dorsdlis, и вентральное, nucleus cochlearis ventrdlis, расположенное латеральнее и кпереди от предыдущего. Другая часть нерва, pars vestibularis - нерв преддверия, или статический нерв, имеет четыре ядра (nuclei vestibulares):
1. Медиальное - главное (ядро Швальбе).
2. Латеральное - ядро Дейтерса.
3. Верхнее - ядро Бехтерева.
4. Нижнее.
Наличие у человека четырех ядер отражает ранние стадии филогенеза, когда у рыб имелось несколько отдельных воспринимающих статических аппаратов.
VII пара - лицевой нерв, n. facialis, имеет одно двигательное ядро, расположенное в formatio reticularis partis dorsalis моста. Отходящие от него нервные волокна на своем пути в толще моста образуют петлю, выпячивающуюся на ромбовидной ямке в виде colliculus facialis.
Промежуточный нерв, n. intermedius, тесно связанный в своем ходе с лицевым нервом, имеет два ядра:
1. Вегетативное (секреторное), nucleus salivatorius superior (верхнее слюноотделительное ядро), заложено в formatio reticularis моста, дорсальнее ядра лицевого нерва.
2. Чувствительное, nucleus trdctus solitdrii.
VI пара - отводящий нерв, n. abdiicens, имеет одно двигательное ядро, заложенное в петле лицевого нерва, поэтому colliculus facialis на поверхности ромбовидной ямки соответствует этому ядру
V пара - тройничный нерв, n. trigeminus, имеет четыре ядра:
1. Чувствительное, nucleus sensorius principalis n. trigemini, проецируется в дорсо-латеральной части верхнего отдела моста.
2. Ядро спинномозгового тракта, nucleus tractus spinalis n. trigemini, является продолжением предыдущего по всему протяжению продолговатого мозга до шейного отдела спинного мозга, где соприкасается с substantia gelatinosa задних рогов.
3. Двигательное ядро, nucleus motorius n. trigemini (жевательное), расположено медиальнее чувствительного.
4. Ядро среднемозгового тракта, nucleus tractus mesencephalici n. trigemini, лежит латеральнее водопровода. Оно представляет ядро проприоцептивной чувствительности для жевательных мышц и для мышц глазного яблока.
Возможно, что это ядро отражает самостоятельное развитие первой ветви тройничного нерва (n. ophthalmicus), называемого у животных n. ophthalmicus profundus и имеющего отношение к органу зрения, чем и объясняется расположение ядра в среднем мозгу.
Mesencephalon
Средний мозг, mesencephalon, развивается в процессе филогенеза под преимущественным влиянием зрительного рецептора, поэтому важнейшие его образования имеют отношение к иннервации глаза. Здесь же образовались центры слуха, которые вместе с центрами зрения в дальнейшем разрослись в виде четырех бугорков - четверохолмие. С появлением у высших животных и человека слухового и зрительного анализаторов в коре переднего мозга слуховые и зрительные центры среднего мозга сами попали в подчиненное положение и стали промежуточными, подкорковыми. С развитием у высших млекопитающих и человека переднего мозга через средний мозг стали проходить проводящие пути, связывающие кору концевого мозга со спинным (ножки мозга) (см. рис. 277, 278).
В результате в среднем мозгу человека имеются: 1) подкорковые центры зрения и ядра нервов, иннервирующие мышцы глаза; 2) подкорковые слуховые центры; 3) все восходящие и нисходящие проводящие пути, связывающие кору головного мозга со спинным и идущие транзитно через средний мозг; 4) пучки белого вещества, связывающие средний мозг с другими отделами центральной нервной системы. Соответственно этому средний мозг, являющийся у человека наименьшим и наиболее просто устроенным отделом головного мозга, имеет две основные части: крышу, где располагаются подкорковые центры слуха и зрения, и ножки мозга, где преимущественно проходят проводящие пути.
1. Дорсальная часть, крыша среднего мозга, или пластинка четверохолмия, tectum mesencephali s. lamina quadrigemina (BNA) (см. рис. 278).
Она скрыта под задним концом мозолистого тела и подразделяется посредством двух идущих крест-накрест канавок - продольной и поперечной - на четыре бугорка белого цвета, располагающихся попарно.
Верхние два бугорка, colliculi superiores, являются подкорковыми центрами зрения; оба нижних, colliculi inferiores, -подкорковыми центрами слуха. В плоской канавке между верхними бугорками лежит шишковидное тело. Каждый бугорок переходит в так называемую ручку бугорка, brachium colliculi, направляющуюся латерально, кпереди и кверху, к промежуточному мозгу. Ручка верхнего бугорка, brdchium colllculi superioris, идет под подушкой, pulvinar, зрительного бугра к боковому коленчатому телу, corpus geniculatum laterale. Ручка нижнего бугорка, brdchium colllculi inferioris, проходя вдоль верхнего края trigonum lemnisci до sulcus lateralis mesencephali, исчезает под медиальным коленчатым телом, corpus geniculatum mediale. Названные коленчатые тела относятся уже к промежуточному мозгу.
2. Вентральная часть, ножки мозга, pedunculi cerebri, содержит все проводящие пути к переднему мозгу.
Ножки мозга, имеют вид двух толстых полуцилиндрических белых тяжей, которые расходятся от края моста под углом и погружаются в толщу полушарий большого мозга.
3. Полость среднего мозга, являющаяся остатком первичной полости среднего мозгового пузыря, имеет вид узкого канала и называется водопроводом,aqueductus cerebri (Sylvii). Он представляет узкий, выстланный эпендимой канал 1,5-2,0 см длиной, соединяющий IV желудочек с III. Дорсально водопровод ограничивается крышей среднего мозга, а вентрально покрышкой ножек мозга.
Внутреннее строение среднего мозга. На поперечном разрезе среднего мозга различают три основные части мозгового ствола: 1) пластинку крыши, lamina tecti, образованную четверохолмием; 2) покрышку, tegmentum, представляющую верхний отдел pedunculi cerebri; 3) вентральный отдел pedunculi cerebri или собственно ножку мозга, crus cerebri. Соответственно развитию среднего мозга под влиянием зрительного рецептора (Е. К. Сепп) в нем заложены различные ядра, имеющие отношение к иннервации глаза.
У низших позвоночных верхнее двухолмие служит главным местом окончания зрительного нерва и является зрительным центром. У млекопитающих и у человека с переносом зрительных центров в передний мозг остающаяся связь зрительного нерва с верхним холмиком имеет значение только для рефлексов. В ядре нижнего холмика, а также в медиальном коленчатом теле оканчиваются волокна слуховой петли (lemniscus lateralis). Крыша четверохолмия имеет двустороннюю связь со спинным мозгом - trdctus spinotectdlis и trdctus tectobulbdris et tectospindlis. Последние после перекреста в покрышке (дорсальный фонтановидный перекрест Мейнерта) идут к мышечным ядрам в продолговатом и спинном мозгу. Это так называемый зрительно-звуковой рефлекторный путь, о котором говорилось при описании спинного мозга. Таким образом, четверохолмие можно рассматривать как рефлекторный центр для различного рода движений, возникающих главным образом под влиянием зрительных и слуховых раздражений.
Сильвиев водопровод окружен центральным серым веществом, имеющим по своей функции отношение к вегетативной системе. В нем, под вентральной стенкой водопровода, в покрышке ножки мозга заложены ядра двух двигательных головных нервов - n. oculomotorius (III пара) на уровне верхнего двухолмия и n. trochlearis (IV пара) на уровне нижнего двухолмия. Ядро глазодвигательного нерва состоит из нескольких отделов соответственно иннервации нескольких мышц глазного яблока. Медиально и кзади от него помещается еще небольшое, тоже парное, вегетативное ядро, nucleus accessorius или ядро Якубовича (Якубович описал его в 1857 r., ранее Вестфаля и Эдингера, именем которых оно неправильно называлось) и непарное срединное ядро. Ядро Якубовича и непарное срединное ядро иннервируют гладкие мышцы глаза, m. ciliaris и m. sphincter pupillae. Эта часть глазодвигательного нерва относится к парасимпатической системе. Выше (оральнее) ядра глазодвигательного нерва в покрышке ножки мозга располагается ядро медиального продольного пучка (ядро Даркшевича).
Латерально от водопровода находится ядро среднемозгового тракта тройничного нерва, nucleus tractus mesencephalici n. trigemini.
Ножки мозга делятся, как уже говорилось, на вентральную часть, или собственно ножку мозга, crus cerebri, и покрышку, tegmentum. Границей между ними служит черное вещество, substantia nigra, обязанное своим цветом содержащемуся в составляющих его нервных клетках черному пигменту - меланину (рис. 284).
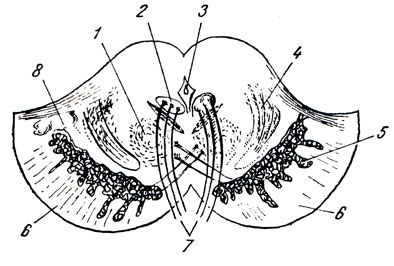
Рис. 284. Поперечный разрез через ножки мозга. 1 - nucleus ruber; 2 - nucleus n. oculomotorii; 3 - aqueductus cerebri; 4 - lemniscus medialis; 5 - substantia nigra; 6 - crus cerebri; 7 - n. oculomotorius, 8 - tegmentum pedunculi
Substantia nigra простирается на всем протяжении ножки мозга от моста до промежуточного мозга; по своей функции относится к экстрапирамидной системе.
Расположенная вентрально от substantia nigra, crus cerebri содержит продольные нервные волокна, спускающиеся от коры полушария большого мозга ко всем нижележащим отделам центральной нервной системы (tractus corticopontine, corticonuclearis, corticospinal и др.). Tegmentum, находящаяся дорсально от substantia nigra, содержит преимущественно различные восходящие волокна, а также ядра серого вещества. Среди них самое значительное - красное ядро, nucleus ruber.
Это удлиненное колбасовидное образование простирается в покрышке ножки мозга от подбугорной области промежуточного мозга до нижнего двухолмия, где от него начинается важный нисходящий тракт, tractus rubrospinalis, соединяющий красное ядро с передними рогами спинного мозга. Пучок этот после выхода из красного ядра перекрещивается с аналогичным пучком противоположной стороны в вентральной части срединного шва - вентральный перекрест покрышки (Фореля). Nucleus ruber является весьма важным координационным центром экстрапирамидной системы, связанным с остальными ее частями. К нему проходят волокна от мозжечка в составе верхних ножек последнего после их перекреста под четверохолмием, вентрально от aqueductus cerebri, а также от pallidum - этого самого нижнего и самого древнего из подкорковых узлов головного мозга, входящих в состав экстрапирамидной системы. Благодаря этим связям мозжечок и экстрапирамидная система через посредство красного ядра, и отходящего от него tractus rubrospinalis оказывают влияние на всю скелетную мускулатуру в смысле бессознательных автоматических движений. Кроме нисходящих продольных волокон, в составе tegmentum идут восходящие, образующие продолжение в среднем мозгу медиальной (lemniscus medialis) и латеральной (lemniscus lateralis) петель. В составе этих петель восходят к большому мозгу все чувствительные пути, за исключением зрительного и обонятельного.
В покрышку среднего мозга продолжается также сетчатая формация, formatio reticularis, и fasciculus longitudindlis medialis. Последний берет начало в различных местах. Одна из его частей начинается из вестибулярных ядер, проходит на той и другой стороне по бокам средней линии, непосредственно под серым веществом дна сильвиева водопровода и IV желудочка, и состоит из восходящих и нисходящих волокон, идущих к ядрам III, IV, VI и XI головных нервов. Медиальный продольный пучок является важным ассоциационным путем, связующим различные ядра нервов глазных мышц между собой, чем обусловливаются сочетанные движения глаз при отклонении их в ту или другую сторону. Функция его связана также с движениями глаз и головы, возникающими при раздражении аппарата равновесия.
Prosencephalon
Передний мозг, prosencephalon, развивается в связи с обонятельным рецептором и вначале (у водных животных) является чисто обонятельным мозгом, rhinencephalon. С переходом животных из водной среды в воздушную роль обонятельного рецептора возрастает, так как с его помощью определяются содержащиеся в воздухе химические вещества, сигнализирующие животному о добыче, опасности и других жизненно важных явлениях природы с далекого расстояния - дистантный рецептор.
Поэтому, а также благодаря развитию и совершенствованию других анализаторов передний мозг у наземных животных сильно разрастается и превосходит другие отделы центральной нервной системы, превращаясь из обонятельного мозга в орган, управляющий всем поведением животного. Соответственно двум основным формам поведения: 1) инстинктивное поведение, основанное на опыте вида (безусловные рефлексы), и 2) индивидуальное поведение, основанное на опыте индивида (условные рефлексы) - в переднем мозгу развиваются две группы центров: 1) базальные, или центральные, узлы полушарий большого мозга ("подкорка"); 2) кора большого мозга. В эти две группы центров переднего мозга поступают все нервные импульсы и к ним протягиваются все афферентные чувствительные пути, которые (за немногими исключениями) предварительно проходят через один общий центр - зрительный бугор, или чертог, thalamus. Приспособление организма к среде путем изменения обмена веществ обусловило возникновение в переднем мозгу высших центров, ведающих вегетативными процессами (подбугорная область, hypothalamus).
Из двух частей переднего мозга, промежуточного мозга, diencephalon, и конечного, telencephalon, кора и подкорковые узлы относятся к конечному мозгу, а зрительный бугор и подбугорная область - к промежуточному. По Е. К. Сеппу (1959), промежуточный мозг развился в процессе филогенеза не из переднего мозга в результате деления последнего, а вместе со средним мозгом. Этим объясняется связь зрительного рецептора не только со средним мозгом, но и с промежуточным, и его зрительным бугром, получившим поэтому соответственное название.
Diencephalon
Промежуточный мозг, diencephalon, залегает под мозолистым телом и сводом, срастаясь по бокам с полушариями конечного мозга. Соответственно сказанному выше о функции и развитии переднего мозга в промежуточном мозгу различают две основные части: 1) дорсальную (филогенетически более молодую) - thalamencephalon - центр афферентных путей, и 2) вентральную (филогенетически более старую) - hypothalamus - высший вегетативный центр. Полостью diencephalon является III желудочек.
Thalamencephalon
Thalamencephalon в свою очередь состоит из трех частей: thalamus- чертог, epithalamus - надталамическая область, и metathalamus - заталамическая область (рис. 285).
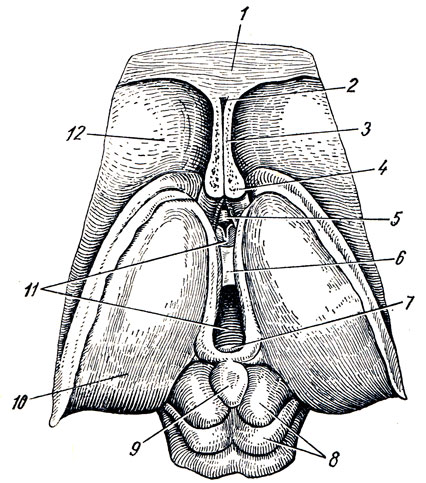
Рис. 285. Промежуточный и средний мозг сверху. 1 - corpus callosum; 2 - cavum septi pellucidi; 3 - septum pellucidum; 4 - fornix (поперечный разрез столбиков); 5 - comissura anterior; 6 - adhesio interthala mica; 7 - comissura posterior; 8 - tectum raesencephali (lamina tecti); 9 - corpus pineale; 10 - thalamus; 11 - ventriculus tertius; 12 - nucleus caudatus (caput)
A. Thalamus, зрительный бугор, представляет большое парное скопление серого вещества в боковых стенках промежуточного мозга по бокам III желудочка, имеющее яйцевидную форму, причем передний его конец заострен в виде tuberculum anterius, а задний расширен и утолщен в виде подушки, pulvinar. Деление на передний конец и подушку соответствует функциональному делению thalamus на центры афферентных путей (передний конец) и на зрительный центр (задний). Дорсальная поверхность покрыта тонким слоем белого вещества - stratum zonule. В латеральном своем отделе она обращена в полость бокового желудочка, отделяясь от соседнего с ней хвостатого ядра пограничной бороздкой, sulcus termindlis, являющейся границей между telencephalon, к которому принадлежит хвостатое ядро, и diencephalon, к которому относится зрительный бугор. По этой бороздке проходит полоска мозгового вещества, stria termindlis.
Медиальная поверхность зрительного бугра, покрытая слоем центрального серого вещества, стоит вертикально и обращена в полость III желудочка, образуя его латеральную стенку. Сверху она отграничивается от дорсальной поверхности посредством белой мозговой полоски, stria medulldris thalami. Обе медиальные поверхности зрительных бугров соединены между собой серой спайкой - adhesio interthaldmica, лежащей почти по середине. Латеральная поверхность зрительного бугра граничит с внутренней сумкой, capsula interna. Нижней своей поверхностью зрительный бугор располагается над ножкой мозга, срастаясь с ее покрышкой (рис. 286). Как видно на разрезах, серая масса зрительного бугра белыми прослойками, laminae medullares thalami, разделяется на отдельные ядра, носящие названия в зависимости от их топографии, - переднее, центральное, медиальное, латеральное и ряд вентральных.
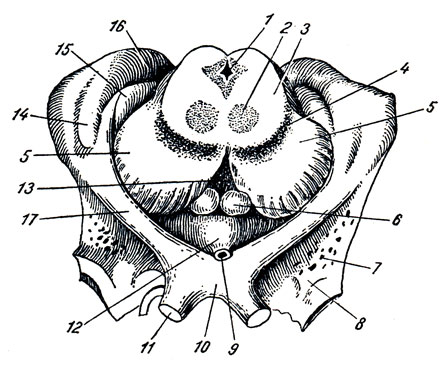
Рис. 286. Metathalamus и hypothalamus. 1 - aqueductus cerebri; 2 - nucleus ruber; 3 - tegmentum; 4 - substantia nigra; 5 - crus cerebri; 6 - corpus mamillare; 7 - substantia perforata anterior; 8 - trigonum olfactorium; 9 - infundibulum; 10 - chiasma opticum; 11 - n. opticus; 12 - tuber cinereum; 13 - substantia perforata posterior; 14 - corpus geniculatum laterale; 15 - corpus geniculatum mediale; 16 - pulvinar; 17 - tractus opticus
Функциональное значение зрительного бугра очень велико. В нем переключаются афферентные пути: в его подушке, pulvinar, оканчивается часть волокон зрительного тракта (подкорковый центр зрения), в переднем ядре - пучок Вик-д'Азира, идущий от corpora mamillaria и связывающий зрительный бугор с обонятельной сферой, и, наконец, все остальные афферентные чувствительные пути от нижележащих отделов центральной нервной системы в остальных его ядрах, причем lemniscus medialis заканчивается в латеральном ядре. Таким образом thalamus является подкорковым центром всех видов чувствительности. Отсюда чувствительные пути идут частью в подкорковые узлы (благодаря чему зрительный бугор становится чувствительным центром экстрапирамидной системы), частью непосредственно в кору (tractus thalamocorticalis).
Б. Epithalamus. Striae medullares обоих зрительных бугров направляются кзади (каудально) и образуют на той и другой стороне треугольное расширение, называемое trigonum habenulae. От последнего отходит так называемый поводок, habenula, который вместе с таким же поводком противоположной стороны соединяется с шишковидным телом, corpus pineale (pinus - сосна) s. epiphysis (см. рис. 285). Спереди от corpus pineale оба поводка связаны вместе посредством commissura habenuldrum. Само шишковидное тело, напоминающее несколько сосновую шишку (отчего и происходит его название), по своему строению и функции относится к железам внутренней секреции. Выдаваясь кзади в область среднего мозга, шишковидное тело располагается в бороздке между верхними бугорками четверохолмия, образуя как бы пятый бугорок.
В. Metathalamus. Позади зрительного бугра находятся два небольших возвышения - коленчатые тела, corpus geniculdtum laterale et mediate (см. рис. 278, 286).
Медиальное коленчатое тело, меньшее по размерам, но более выраженное, лежит спереди нижней ручки четверохолмия под pulvinar зрительного бугра, отделенное от него ясной бороздкой. В нем заканчиваются волокна слуховой петли, lemniscus lateralis, вследствие чего оно является вместе с нижними бугорками четверохолмия подкорковым центром слуха. Латеральное коленчатое тело, большее, в виде плоского бугорка, помещается на нижней латеральной стороне pulvinar. В нем оканчивается большей своей частью латеральная часть зрительного тракта (другая часть тракта оканчивается в pulvinar). Поэтому вместе с pulvinar и верхними бугорками четверохолмия латеральное коленчатое тело является подкорковым центром зрения. Ядра обоих коленчатых тел центральными путями связаны с корковыми концами соответственных анализаторов.
Hypothalamus
Подталамаческая область, hypothdlamus, в широком смысле слова, объединяет образования, расположенные вентрально под дном III желудочка, впереди substantia perforata posterior, включая и подбугорную область в узком смысле, regio subthalamica. Соответственно эмбриональному развитию hypothalamus делится на два отдела: передний - зрительный, pars optica hypothalami, под именем которого объединяют tuber cinereum с infundibulum и hypophysis, а также chiasma opticum с tractus opticus, и задний - обонятельный: corpora mamillaria и regio subthalamica.
A. Tuber cinereum, серый бугор, находящийся спереди от corpora mamillaria, представляет непарный полый выступ нижней стенки III желудочка, состоящий из тонкой пластинки серого вещества. Верхушка бугра вытянута в узкую полую воронку, infundibulum, на слепом конце которой находится мозговой придаток, hypophysis (glandula pituitaria), лежащий в углублении турецкого седла (описание его см. в разделе "Органы внутренней секреции"). В tuber cinereum заложены ядра серого вещества, являющиеся высшими вегетативными центрами, влияющими, в частности, на обмен веществ и теплорегуляцию.
Б. Chiasma opticum, зрительный перекрест, лежит впереди серого бугра, образован перекрестом зрительных нервов, nn. optici.
В. Corpora mamillaria, сосцевидные тела, два небольших белого цвета возвышения неправильно шаровидной формы, лежащих симметрично по бокам средней линии, спереди от substantia perforata posterior. Под поверхностным слоем белого вещества внутри каждого из тел находятся два серых ядра.
По своей функции corpora mamillaria относятся к подкорковым обонятельным центрам.
Г. Regio subthalamica, подбугорная область (в узком смысле слова); это небольшой участок мозгового вещества, расположенный под зрительным бугром. В нем латеральнее substantia nigra залегает принадлежащее промежуточному мозгу овальное тело, corpus subthalamicum Luysi. Люисово тело является одним из звеньев экстрапирамидной системы; ему также приписывают и вегетативные функции.
III желудочек
III желудочек, ventriculus tertlus, расположен как раз по средней линии и на фронтальном разрезе мозга имеет вид узкой вертикальной щели. Боковые стенки III желудочка образованы медиальными поверхностями зрительных бугров, между которыми почти по середине перекидывается adhesio interthalamica (см. рис. 285). Переднюю стенку желудочка составляет снизу тонкая пластинка, lamina terminalis, а дальше кверху - столбики свода (columnae fornicis) с лежащей поперек белой передней спайкой, commissura cerebri anterior. По бокам у передней стенки желудочка столбики свода вместе с передними концами зрительных бугров ограничивают межжелудочковые отверстия, foramina interventricularia (Monroi), соединяющие полость III желудочка с боковыми желудочками, залегающими в полушариях конечного мозга. Верхняя стенка III желудочка, лежащая под сводом и мозолистым телом, представляет собой tela chorioidea ventriculi tertii (рис. 287), в состав последней входят недоразвитая стенка мозгового пузыря в виде эпителиальной пластинки, lamina epitelialis и сросшаяся с ней сосудистая оболочка. По бокам от средней линии в tela chorioidea заложено ворсинчатое сплетение, plexus chorioideus ventriculi tertii. В области задней стенки желудочка находятся commissura habeniilarum et commissura cerebri posterior, между которыми вдается в каудальную сторону слепой выступ желудочка, recessus pinealis. Вентрально от commissura posterior открывается в III желудочек воронкообразным отверстием водопровод. Нижняя узкая стенка III желудочка, отграниченная изнутри от боковых стенок бороздками (sulci hypothalamici), со стороны основания мозга соответствует substantia perforata posterior, corpora marnillaria, tuber cinereum с chiasma opticum. В области дна полость желудочка образует два углубления: recessus infundibuli, вдающийся в серый бугор и в воронку, и recessus opticus, лежащий кпереди хиазмы. Внутренняя поверхность стенок III желудочка покрыта эпендимой.
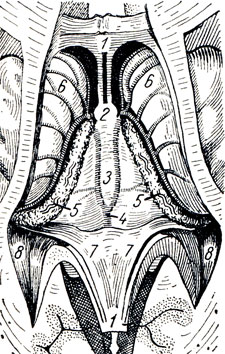
Рис. 287. Часть головного мозга со вскрытыми боковыми желудочками. Мозолистое тело перерезано и вместе со сводом отвернуто кзади для демонстрации tela chorioidea. 1 - corpus callosum; 2 - columnae fornicis (перерезаны); 3 - tela chorioidea ventriculi tertii; 4 - v. cerebri magna; 5 - plexus chorioideus ventriculi lat.; 6 - nucleus caudatus; 7 - crus fornicis; 8 - cornu posterius ventriculi lat
Рассмотренные отделы мозга - ромбовидный (кроме мозжечка), средний и промежуточный объединяются под названием мозгового ствола. Клиницисты иногда к мозговому стволу относят только задний и средний мозг (Е. К. Сепп, М. Б. Цукер, Е. В. Шмидт, 1950). Мозговой ствол, филогенетически более старое образование, существенно отличается по строению и функции от более молодой части головного мозга - конечного мозга, telencephalon.
Telencephalon
Как уже говорилось выше, конечный мозг, telencephalon, представлен двумя полушариями, hemispheria cerebri. В состав каждого полушария входят: плащ, или мантия, pallium, обонятельный мозг, rhinencephalon, и узлы основания. Остатком первоначальных полостей обоих пузырей конечного мозга являются боковые желудочки, ventriculi laterales. Передний мозг, из которого выделяется конечный, вначале возникает в связи с обонятельным рецептором (обонятельный мозг), а затем он становится органом управления поведения животного, причем в нем возникают центры инстинктивного поведения, основанного на видовых реакциях (безусловные рефлексы), - подкорковые узлы, и центры индивидуального поведения, основанного на индивидуальном опыте (условные рефлексы), - кора большого мозга. Соответственно этому в концевом мозгу различают в порядке исторического развития следующие группы центров:
1. Обонятельный мозг, rhinencephalon, самая древняя и вместе с тем самая меньшая часть, расположенная вентрально.
2. Базальные, или центральные, узлы полушарий, "подкорка", старая часть конечного мозга, paleencephalon, скрытая в глубине.
3. Серое вещество коры, cortex, самая молодая часть, neencephalon, и вместе с тем самая большая часть, покрывающая остальные как бы плащом, откуда и ее название - плащ, или мантия, pallium.
Кроме отмеченных для животных двух форм поведения, у человека возникает третья форма - коллективное поведение, основанное на опыте человеческого коллектива, создающегося в процессе трудовой деятельности человека и общения людей с помощью речи. Эта форма поведения связана с развитием самых молодых поверхностных слоев мозговой коры, составляющих материальный субстрат так называемой второй сигнальной (словесной) системы действительности (И. П. Павлов).
Так как в процессе эволюции из всех отделов центральной нервной системы быстрее и сильнее всего растет конечный мозг, то он у человека становится самой большой частью головного мозга и приобретает вид двух объемистых полушарий - правого и левого, hemispherium dextrum et sinistrum. В глубине продольной щели мозга оба полушария соединены между собой толстой горизонтальной пластинкой - мозолистым телом, corpus callosum, которое состоит из нервных волокон, идущих поперечно из одного полушария в другое. В мозолистом теле различают передний загибающийся книзу конец, или колено, genu corporis calldsi, среднюю часть, truncus corporis calldsi, и затем задний конец, утолщенный в форме валика, spleniutn corporis calldsi. Все эти части хорошо видны на продольном разрезе мозга между обоими полушариями (см. рис. 282). Колено мозолистого тела, загибаясь книзу, заостряется и образует клюв, rostrum corporis callosi, который переходит в тонкую пластинку, lamina rostralis, продолжающуюся в свою очередь в lamina terminalis.
Под мозолистым телом находится так называемый свод, fornix (см. рис. 282; рис. 288), представляющий два дугообразных белых тяжа, которые в средней своей части, corpus fornicis, соединены между собой, а спереди и сзади расходятся, образуя впереди столбики свода, соlumnae fornicis, позади же - ножки свода, crura fornicis. Crura fornicis, направляясь назад, спускаются в нижние рога боковых желудочков и переходят там в fimbria hippocampi. Между crura fornicis под splenium corporis callosi протягиваются поперечные пучки нервных волокон, образующие commissura fornicis. Передние концы свода, columnae fornicis, продолжаются вниз до основания мозга, где оканчиваются в corpora mamillares, проходя через серое вещество hypothalamus. Columnae fornicis ограничивают лежащие позади них межжелудочковые отверстия, соединяющие III желудочек с боковыми желудочками. Впереди столбиков свода находится передняя спайка, commissura anterior, имеющая вид белой поперечной перекладины, состоящей из нервных волокон. Между передней частью свода и genu corporis callosi натянута тонкая вертикальная пластинка мозговой ткани - прозрачная перегородка, septum pellucidum, в толще которой находится небольшая щелевидная полость, cavum septi pellucidi (см. рис. 282, 293).
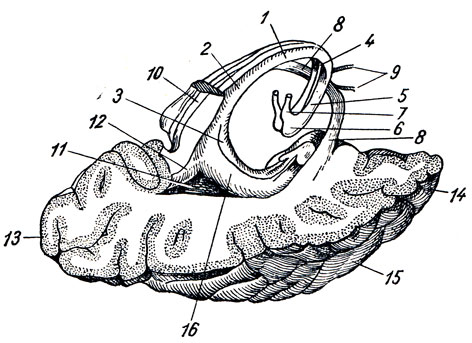
Рис. 288. Свод, гиппокамп и передняя комиссура. 1 - corpus fornicis; 2 - cms fornicis; 3 - fimbria hippocampi; 4, 5 - columna fornicis; 6 - corpus mamillare; 7 - fasciculus thalamomamillaris (Vicq - d'Azyr); 8, 9 - comissura anterior; 10 - splenium corporis callosi; 11 - trigonum collaterale; 12 - calcar avis; 13 - polus occipitalis; 14 - polus temporalis; 15 - gyrus temporalis medius; 16 - hippocampus
Для удобства изучения начнем описание частей в порядке, обратном историческому развитию, т. е. с плаща, который закрывает остальные части.
Pallium
В каждом полушарии можно различать три поверхности: верхнелатеральную, медиальную и нижнюю, и три края: верхний, нижний и медиальный, три конца, или полюса: передний полюс, polus frontalis, задний, polus occipitalis, и затем polus temporalis, соответствующий выступу нижней поверхности и отделенный на ней ямкой, fossa lateralis cerebri (Sylvii).
Поверхность полушария (плащ), образована равномерным слоем серого вещества, 1,3-4,5 мм толщины, содержащего нервные клетки. Слой этот, называемый также корой большого мозга, cortex cerebri, представляется как бы сложенным в складки, благодаря чему поверхность плаща имеет в высшей степени сложный рисунок, состоящий из чередующихся между собой в различных направлениях борозд и валиков между ними, называемых извилинами*, gyri. Величина и форма борозд подвержены значительным индивидуальным колебаниям, вследствие чего не только мозг различных людей, но даже полушария одной и той же особи по рисунку борозд не вполне похожи (см. классификацию борозд).
* (Общая площадь коры взрослого человека около 220000 мм2 (Генеберг), причем 2/3 залегают в глубине между извилинами и только 1/3 лежит на поверхности.)
Глубокими постоянными бороздами пользуются для разделения каждого полушария на большие участки, называемые долями, lobi; последние в свою очередь разделяются на дольки и извилины. Долей каждого полушария пять: лобная (lobus frontalis), теменная (lobus parietalis), височная (lobus temporalis), затылочная (lobus occipitalis) и долька, скрытая на дне боковой (сильвиевой) борозды, так называемый островок (insula).
Верхнелатеральная поверхность полушария разграничена на доли посредством трех борозд (см. рис. 274; рис. 289): латеральной, центральной и верхнего конца теменно-затылочной борозды, которая, находясь на медиальной стороне полушария, образует зарубку на его верхнем краю. Латеральная борозда, sulcus cerebri lateralis (Sylvii), начинается на базальной поверхности полушария из сильвиевой ямы и затем переходит на верхнелатеральную поверхность, направляясь назад и несколько вверх. Она оканчивается приблизительно на границе средней и задней трети верхнелатеральной поверхности полушария. В передней части латеральной борозды от нее отходят две небольшие ветви: одна, ramus ascendens, и другая, ramus anterior, направляющиеся в лобную долю.
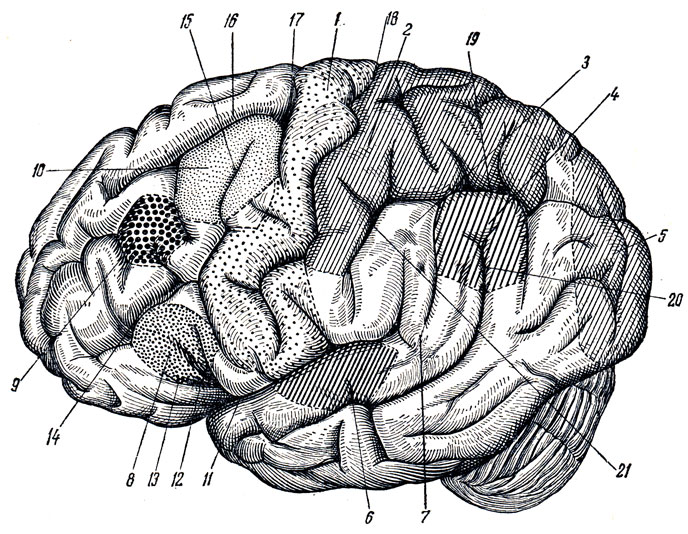
Рис. 289. Ядра анализаторов в коре большого мозга (головной мозг сбоку). 1 - ядро двигательного анализатора; 2 - ядро кожного анализатора; 3 - ядро двигательного анализатора, посредством которого синтезируются привычные целенаправленные движения; 4 - ядро зрительного анализатора письменной речи; 5 - ядро зрительного анализатора (зрительная память); 6 - ядро слухового анализатора; 7 - ядро слухового анализатора речи; 8 - ядро двигательного анализатора артикуляции речи; 9 - ядро двигательного анализатора, имеющего отношение к сочетанному повороту головы и глаз; 10 - ядро двигательного анализатора письменной речи; 11, 12, 13 - sulcus cerebri lateralis (11 - сама борозда; 12 - ramus ascendens; 13 - ramus anterior); 14 - sulcus frontalis inferior; 15 - sulcus precentralis inferior; 16 - sulcus frontalis superior; 17 - sulcus precentralis superior; 18 - sulcus centralis (Rolandi); 19 - sulcus intraparietalis; 20 - sulcus temporalis superior; 21 - sulcus postcentralis
Центральная, или роландова, борозда, sulcus centralis (Rolandi), начинается на верхнем краю полушария, несколько кзади от его середины, и идет вперед и вниз. Нижний конец центральной борозды не доходит до латеральной борозды. Участок полушария, находящийся впереди центральной борозды, относится к лобной доле; часть мозговой поверхности, лежащая сзади от центральной борозды, составляет теменную долю, которая посредством задней части латеральной борозды отграничивается от лежащей ниже височной доли. Задней границей теменной доли служит конец вышеупомянутой теменно-затылочной борозды, sulcus parietooccipitalis, расположенной на медиальной поверхности полушария, но эта граница неполная, ибо названная борозда не заходит далеко на верхнелатеральную поверхность, вследствие чего теменная доля непосредственно переходит в затылочную. Эта последняя также не имеет резкой границы, которая отделяла бы ее от впереди лежащей височной доли. Вследствие этого граница между только что упомянутыми долями проводится искусственно посредством линии, идущей от теменно-затылочной борозды к нижнему краю полушария.
Каждая доля состоит из ряда извилин, называемых местами дольками, которые ограничиваются бороздами мозговой поверхности.
Лобная доля. В заднем отделе наружной поверхности этой доли проходит sulcus precentralis почти в параллельном направлении к sulcus centralis. От нее в продольном направлении отходят две борозды: sulcus frontalis superior et sulcus frontalis inferior. Благодаря этому лобная доля разделяется на четыре извилины - одну вертикальную и три горизонтальные. Вертикальная извилина, gyrus precentralis, находится между sulcus centralis и sulcus precentralis.
Горизонтальные извилины лобной доли следующие: 1) верхняя лобная, gyrus frontalis superior, которая идет выше sulcus frontalis superior, параллельно верхнему краю полушария, заходя и на его медиальную поверхность; 2) средняя лобная извилина, gyrus frontalis medius, тянется между верхней и нижней лобными бороздами и 3) нижняя лобная извилина, gyrus frontalis inferior, помещается между sulcus frontalis inferior и латеральной бороздой. Ветви латеральной борозды, вдающиеся в нижнюю лобную извилину, делят последнюю на три части: pars opercularis, лежащую между нижним концом sulcus precentralis и ramus ascendens sulci lateralis, pars triangularis, находящуюся между обеими ветвями латеральной борозды, и, наконец, pars orbitalis, помещающуюся впереди от ramus anterior sulci lateralis.
Теменная доля. На ней приблизительно параллельно центральной борозде располагается sulcus postcentralis, сливающаяся обычно с sulcus intraparietalis, идущей в горизонтальном направлении. В зависимости от расположения этих борозд теменная доля разделяется на три извилины, из которых одна вертикальная, а две другие горизонтальные. Вертикальная извилина, gyrus postcentralis, идет позади sulcus centralis в одном направлении с gyrus precentralis, отделенная от нее центральной бороздой. Выше sulcus intraparietalis помещается верхняя теменная извилина, или долька, lobulus parietalis superior, которая распространяется и на медиальную поверхность полушария. Ниже sulcus intraparietalis лежит lobulus parietalis inferior, которая, направляясь назад, огибает концы латеральной борозды и sulcus temporalis superior и теряется в области затылочной доли. Часть lobulus parietalis inferior, огибающая латеральную борозду, называется gyrus supramargindlis; другая часть, которая огибает sulcus temporalis superior, носит название gyrus anguldris.
Височная доля. Латеральная поверхность этой доли имеет три продольные извилины, отграниченные друг от друга sulcus temporalis superior и sulcus temporalis inferior. Верхняя из извилин, gyrus temporalis superior, находится между латеральной бороздой и sulcus temporalis superior. Верхняя ее поверхность, скрытая в глубине латеральной борозды, несет 2-3 короткие извил инки, называемые gyri tempordles transversi (извилины Гешля). Между верхней и нижней височными бороздами протягивается gyrus temporalis medius. Ниже последней, отделяясь от нее sulcus temporalis inferior, проходит gyrus temporalis inferior, которая посредством нижнего края отделена от лежащей на нижней поверхности gyrus occipitotemporalis lateralis.
Затылочная доля. Борозды латеральной поверхности этой доли изменчивы и непостоянны. Из них выделяют идущую поперечно sulcus occipitalis transversus, соединяющуюся обыкновенно с концом sulcus intraparietalis.
Островок, insula. Чтобы увидеть эту дольку, надо раздвинуть или удалить нависающие над ней края латеральной борозды. Эти края, относясь к лобной, теменной и височной долям, носят название покрышки, operculum. Островок имеет форму треугольника, верхушка которого обращена вперед и вниз. Спереди, сверху и сзади островок отграничивается от соседних с ним частей посредством глубокой борозды, sulcus circularis. Поверхность островка покрыта короткими извилинами.
Нижняя поверхность полушария (см. рис. 276) в той ее части, которая лежит кпереди от сильвиевой ямы, относится к лобной доле. Здесь параллельно медиальному краю полушария проходит sulcus olfactorius, в которой лежат bulbus et tractus olfactorius. Между этой бороздой и медиальным краем полушария протягивается прямая извилина, gyrus rectus, представляющая собой продолжение верхней лобной извилины. Латерально от sulcus olfactorius на нижней поверхности находится несколько непостоянных бороздок, sulci orbitales, ограничивающих gyri orbitales, которые можно рассматривать как продолжение средней и нижней лобных извилин. Задний участок базальной поверхности полушария образован нижней поверхностью височной и затылочной долей, которые здесь не имеют определенных границ. На этом участке видны две борозды: sulcus occipitotemporalis, проходящая в направлении от затылочного полюса к височному и ограничивающая gyrus occipitotemporalis lateralis и идущая параллельно ей sulcus collaterals (продолжением ее кпереди является sulcus rhindlis). Между ними располагается gyrus occipitotemporalis medialis. Медиально от sulcus collateralis расположены две извилины: между задним отделом этой борозды и sulcus calcarinus лежит gyrus lingualis; между передним отделом этой борозды и sulcus rhinalis с одной стороны и глубокой sulcus hippocampi, огибающей ствол мозга - с другой лежит gyrus parahippocampalis. Эта извилина, примыкающая к стволу мозга, находится уже на медиальной поверхности полушария.
Медиальная поверхность полушария. На этой поверхности (см. рис. 282) находится борозда мозолистого тела, sulcus corporis callosi, идущая непосредственно над мозолистым телом и продолжающаяся своим задним концом в глубокую sulcus hippocampi, которая направляется вперед и книзу. Параллельно и выше этой борозды проходит по медиальной поверхности полушария sulcus cinguli, которая начинается спереди под клювом мозолистого тела, затем идет назад и оканчивается своим задним концом на верхнем краю полушария. Пространство, располагающееся между этим краем полушария и sulcus cinguli, относится к лобной доле, к верхней лобной извилине. Небольшой участок над sulcus cinguli, ограниченный сзади задним концом sulcus cinguli, а спереди маленькой бороздкой, sulcus paracentrdlis, называется околоцентральной долькой, lobulus paracentralis, так как он соответствует медиальной поверхности верхних концов обеих центральных извилин, переходящих здесь друг в друга.
Кзади от lobulus paracentralis находится четырехугольная поверхность (так, называемое предклинье, precuneus), ограниченная спереди концом sulcus cinguli, снизу небольшой sulcus subparietdlis, а сзади глубокой sulcus parietooccipitalis. Precuneus относится к теменной доле. Позади precuneus лежит резко обособленный участок коры, относящийся к затылочной доле, клин, cuneus, который ограничен спереди sulcus parietooccipitalis, а сзади sulcus calcarfnus, сходящимися под углом. Книзу и кзади клин соприкасается с gyrus lingualis. Между sulcus cinguli и бороздкой мозолистого тела протягивается поясная извилина, gyrus cinguli, которая при посредстве перешейка, isthmus, продолжается в gyrus parahippocampalis, заканчивающуюся крючком, uncus. Парагиппокампова извилина ограничивается с одной стороны sulcus hippocampi, огибающей ствол мозга, а с другой - sulcus collateralis и ее продолжением кпереди, носящим название sulcus rhinalis. Isthmus - суженное место перехода поясной извилины в парагиппокампову, находится позади splenium corporis callosi, у конца борозды, образовавшейся от слияния sulcus parietooccipitalis с sulcus calcaneus. Gyrus cinguli, isthmus и gyrus parahippocampalis образуют вместе сводчатую извилину, gyrus fornicatus, которая описывает почти полный круг, открытый только снизу и спереди. Сводчатая извилина не имеет отношения ни к одной из долей плаща.
Раздвигая край sulcus hippocampi, можно видеть узкую зазубренную серую полоску, представляющую собой рудиментарную извилину gyrus dentatus.
Строение мозговой коры. Кора полушарий большого мозга в основном своем типе состоит из шести слоев, различающихся между собой главным образом по форме входящих в них нервных клеток (рис. 290): 1) молекулярный слой лежит непосредственно под pia mater и содержит концевые разветвления отростков нервных клеток, переплетающихся сетеобразно; 2) наружный зернистый слой, называющийся так потому, что в его состав входят многочисленные маленькие клетки, похожие на зерна; 3) слой пирамид состоит из малых и средних пирамидальных нервных клеток; 4) внутренний зернистый слой слагается, так же как и наружный такой же слой, из маленьких клеток - зерен; 5) ганглионарный слой содержит большие пирамидальные клетки (клетки Беца); 6) слой полиморфных клеток граничит с белым веществом. Из этих 6 слоев нижние (5-й и 6-й) являются преимущественно началом эфферентных путей; в частности, 5-й слой состоит из пирамидных клеток Беца, аксоны которых составляют пирамидную систему. Средние слои (3-й и 4-й) связаны преимущественно с афферентными путями, а верхние (1-й и 2-й) относятся к ассоциативным путям коры. Шестислойный тип коры видоизменяется в различных областях как в смысле толщины и расположения слоев, так и состава клеток (о чем подробно излагается в курсе гистологии).
Рис. 290. Схема строения коры головного мозга. 1 - молекулярный слой; 2 - наружный зернистый слой; 3 - слой малых и средних пирамид; 4 - внутренний зернистый слой; 5 - ганглиозный слой; 6 - слой полиморфных клеток; 7 - белое вещество
Обонятельный мозг (rhinencephalon)
Обонятельный мозг, rhinencephaton (рис. 291, 292), есть филогенетически самая древняя часть переднего мозга, возникающая в связи с рецептором обоняния, когда передний мозг не стал еще органом поведения животного. Поэтому все компоненты его являются различными частями обонятельного анализатора (понятие об анализаторе.
Рис. 291. Развитие neopallium (по Эдингеру). Слева - змеи (Python), справа - сумчатого млекопитающего (Hypsiprimnus). 1 и 3 - neopallium; 2 - archipallium; 4 - hippocampus
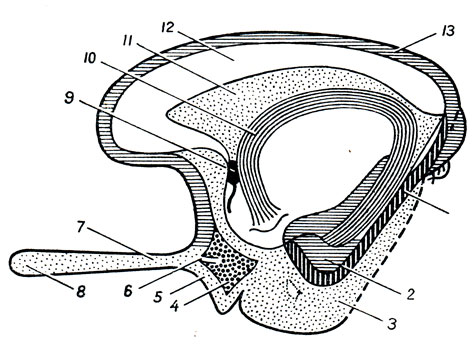
Рис. 292. Обонятельный мозг (схема). 1 - gyrus dentatus; 2 - gyrus parahippocampalis; 3 - uncus; 4 - substantia perforata anterior; 5, 6 - stria olfactoria; 7 - tractus olfactorius; 8 - bulbus olfactorius; 9 - commissura anterior; 10 - fornix; 11 - septum pellucidum; 12 - corpus callosum; 13 - gyrus fornicatus
У рыб почти весь передний мозг является органом обоняния. С развитием новой коры, что наблюдается у млекопитающих и человека, развивается новая часть переднего мозга (neencephalon) - плащ, pallium. Но и плащ проходит свой длинный путь развития и содержит три части различной филогенетической давности. Более старые части:
1. Archipallium, входящий в состав височной доли. Вначале этот отдел располагался на латеральной поверхности полушария, но в дальнейшем, под влиянием сильно увеличивающегося neopallium, он свернулся в колбасовидное образование - гиппокамп, или аммонов рог, и сместился медиально в полость бокового желудочка концевого мозга в виде выпячивания его нижнего рога. Аммонов рог покрыт древней корой, archicortex.
2. Paleopallium, небольшой участок коры на вентральной поверхности лобной доли, лежащий вблизи bulbus olfactorius и покрытый старой корой, paleocortex.
3. Neopallium, новый плащ, в коре которого, neocortex, появились высшие центры обоняния - корковые концы анализатора. Это - uncus, являющийся частью сводчатой извилины.
В результате обонятельный мозг человека содержит ряд образований различного происхождения, которые топографически можно разделить на два отдела. Периферический отдел - это обонятельная доля, lobus olfactorius, под которой разумеется ряд образований, лежащих на основании мозга: 1) bulbus olfactorius; 2) tractus olfactorius; 3) trigonum olfactonum и 4) substantia perforata anterior. Центральный отдел - это извилины мозга: 1) извилина морского коня, hippocampus; 2) зубчатая извилина, gyrus dentatus; 3) сводчатая извилина, gyrus fornicatus, с расположенной вблизи височного полюса передней ее частью - крючком, uncus.
Боковые желудочки
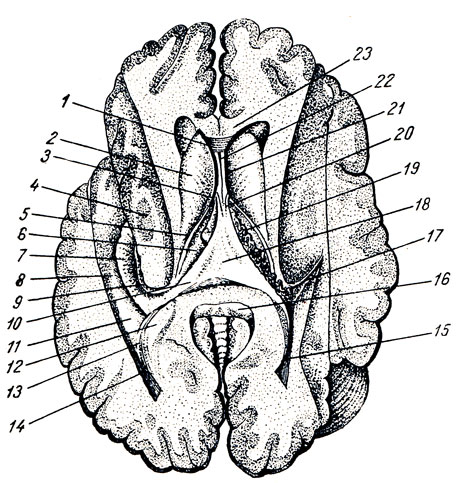
Рис. 293. Боковые желудочки, вскрытые сверху путем удаления части полушарий вместе с мозолистым телом. 1 - cornu anterius; 2 - nucleus caudatus (caput); 3 - foramen interventricular; 4 - nucleus lentiformis (в разрезе); 5 - stria terminalis; 6 - верхняя поверхность thalamus; 7 - hippocampus; 8 - eminentia collateralis; 9 - fimbria hippocampi; 10 - crus fornicis; 11 - cornu posterior ventr. lat.; 12 - медиальная стенка заднего рога; 13 - calcar avis; 14, 15 - cornu posterius; 16 - splenium corporis callosi; 17, 19 - plexus chorioideus в центральной части бокового желудочка и продолжение его в нижний рог; 18 - commissura fornicis; 20 - columnae fornicis; 21 - septum pellucidum; 22 - cavum septi pellucidi; 23 - corpus callosum
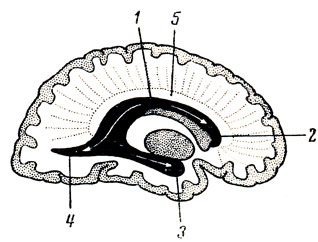
Рис. 294. Сагиттальный разрез левого полушария (немного латеральнее срединной плоскости для демонстрации отделов бокового желудочка). 1 - pars centralis; 2 - cornu anterius; 3 - cornu inferius; 4 - cornu posterius; 5 - corpus callosum
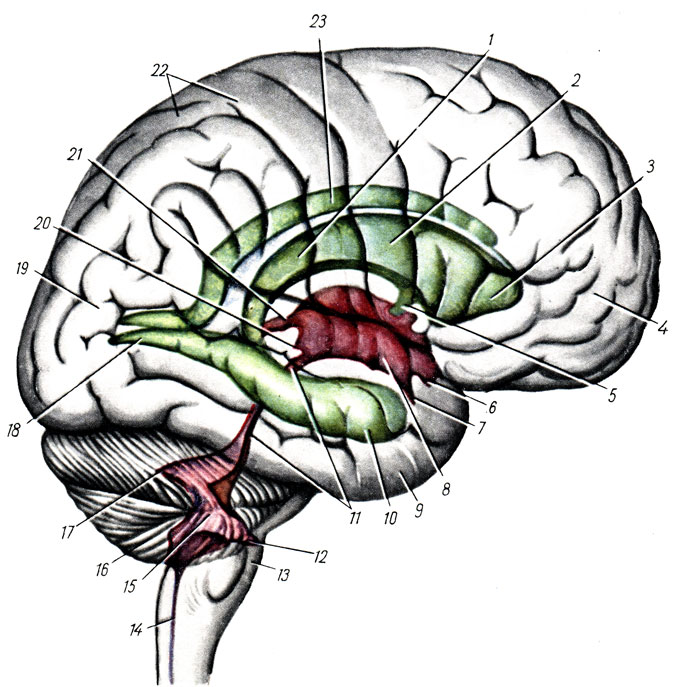
Рис. 295. Желудочки мозга, ventriculi cerebri сбоку (схема). Пространственные взаимоотношения между полушариями головного мозга, мозжечка, мозговым стволом, представленными как бы прозрачными, и желудочками мозга (по Р. Д. Синельникову). 1 - ventriculus lateralis dexter; 2 - pars centralis ventriculi lateralis; 3 - cornu anterius ventriculi lateralis; 4 - lobus frontalis; 5 - foramen interventriculare; 6 - recessus opticus; 7 - recessus infundibuli; 8 - ventriculus tertius; 9 - lobus temporalis; 10 - cornu inferius ventriculi lateralis; 11 - aqueductus cerebri; 12 - recessus lateralis ventriculi quarti; 13 - medulla oblongata; 14 - canalis centralis; 15 - ventriculus quartus; 16 - cerebellum; 17 - fastigium; 18 - cornu posterius ventriculi lateralis; 19 - lobus occipitalis; 20 - recessus pinealis; 21 - заднее углубление третьего желудочка; 22 - lobus parietalis; 23 - ventriculus lateralis sinister
В полушариях как остаток первоначальных полостей обоих пузырей конечного мозга залегают ниже уровня мозолистого тела симметрично по сторонам средней линии два боковых желудочка, ventriculi laterdles, отделенные от верхнелатеральной поверхности полушарий всей толщей мозгового вещества. Полость каждого бокового желудочка (см. рис. 294) соответствует форме полушария: она начинается в лобной доле в виде загнутого вниз и в латеральную сторону переднего рога, cornu anterius, отсюда она через область теменной доли тянется под названием центральной части, pars centralis, а затем на уровне заднего края мозолистого тела поворачивает вниз и идет вперед в толще височной доли в виде нижнего рога, cornu inferius, где и оканчивается. В том месте, где полость желудочка спускается вниз, она дает от себя выступ кзади в затылочную долю - задний рог, cornu posterius.
Медиальная стенка переднего рога образована septum pellucidum, которая отделяет передний рог от такого же рога другого полушария (см. рис. 293). Латеральная стенка и отчасти дно переднего рога заняты возвышением серого цвета, головкой хвостатого ядра, caput nuclei caudati, а верхняя стенка образуется волокнами мозолистого тела. Крыша центральной, наиболее узкой части бокового желудочка также состоит из волокон мозолистого тела, дно же составляется из продолжения хвостатого ядра, corpus nuclei caudati, и части верхней поверхности зрительного бугра. Задний рог окружен слоем белых нервных волокон, происходящих из мозолистого тела, так называемого tapetum (ковер); на его медиальной стенке заметен валик - птичья шпора, calcar avis, образованная вдавлением со стороны sulcus calcarinus, находящейся на медиальной поверхности полушария. Верхнелатеральная стенка нижнего рога образуется tapetum, составляющим продолжение такого же образования, окружающего задний рог. С медиальной стороны на верхней стенке проходит загибающаяся книзу и кпереди утонченная часть хвостатого ядра - cauda nuclei caudati.
По медиальной стенке нижнего рога на всем его протяжении тянется белого цвета возвышение - аммонов рог, cornu Ammonis, или морской конь, hippocampus, который образуется вследствие вдавления от глубоко врезывающейся снаружи sulcus hippocampi. Передний конец hippocampus разделяется бороздками на несколько небольших бугорков. По медиальному краю аммонова рога идет так называемая бахромка, fimbria hippocampi, представляющая продолжение ножки свода (crus fornicis). На дне нижнего рога находится валик, eminentia collaterdlis, происходящий от вдавления снаружи одноименной борозды. С медиальной стороны бокового желудочка в его центральную часть и нижний рог вдается мягкая мозговая оболочка, образующая в этом месте сосудистое сплетение, plexus chorioideus ventrlculi lateralis. Сплетение покрыто эпителием, представляющим остаток неразвитой медиальной стенки желудочка. Plexus chorioideus ventriculi lateralis является латеральным краем tela chorioidea ventriculi tertii.
Базальные, или центральные, узлы или ядра, полушарий
Кроме серой коры на поверхности полушария, имеются еще скопления серого вещества в его толще, именуемые базальными, центральными, или подкорковыми, ядрами и составляющие то, что для краткости называют "подкоркой". В отличие от коры, имеющей строение экранных центров, подкорковые ядра имеют строение ядерных центров. Различают три скопления подкорковых ядер: corpus striatum, claustrum и corpus amygdaloideum (рис. 296, 297).
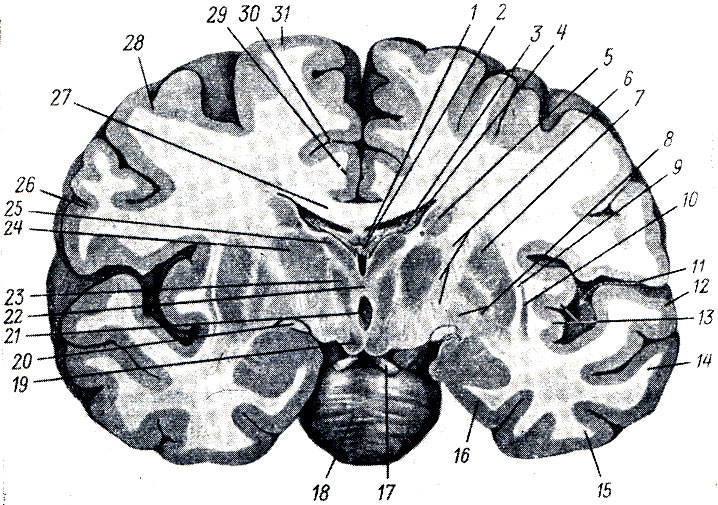
Рис. 296. Фронтальный разрез полушарий, проходящий через corpus striatum и thalamus. 1 - fornix; 2 - plexus chorioideus ventriculi tertii; 3 - plexus chorioideus ventriculi lateralis; 4 - ventriculus lateralis; 5 - nucleus caudatus; 6 - crus posterius внутренней капсулы; 7 - putamen; 8 - globus pallidus; 9 - capsula externa; 10 - claustrum; 11 - sulcus cerebri lateralis; 12, 14, 15 - gyri temporales superior, medius et inferior; 13 - insula; 16 - gyrus parahippocampalis 17 - n. oculomotorius; 18 - pons; 19 - nucleus corporis mamillaris; 20 - tractus opticus; 21 - ventriculus tertius; 22 - adhesio interthalamijpa; 23, 24, 25 - ядра thalamus; 26, 28, 31 - gyri frontales inferior, medius, superior; 27 - corpus callosum; 29 - gyrus cinguli; 30 - sulcus cinguli
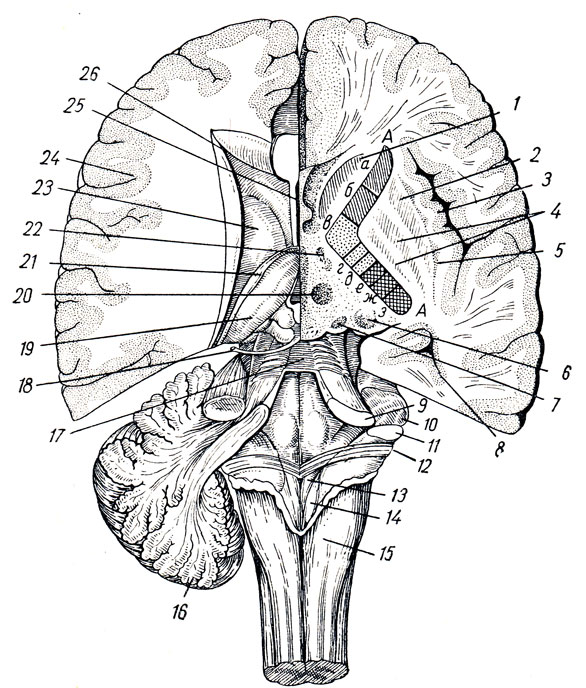
Рис. 297. Полушария мозга на разных уровнях горизонтального разреза. Справа - ниже уровня дна бокового желудочка; слева - над дном бокового желудочка. IV желудочек мозга вскрыт сверху. 1 - nucleus caudatus; 2 - putamen; 3 - кора островка полушария; 4 - globus pallidus; 5 - claustrum; 6 - cauda nuclei caudati; 7 - nucleus corporis geniculati medialis; 8 - cornu inferius ventriculi lateralis; 9 - pedunculus cerebellaris superior; 10 - pedunculus cerebellaris medius; 11 - pedunculus cerebellaris inferior; 12 - striae medullares; 13 - trigonum n. hypoglossi; 14 - trigonum n. vagi; 15 - tuberculum nuclei gracilis; 16 - cerebellum; 17 - velum medullare superius; 18 - n. trochlearis; 19 - thalamus; 20 - nucleus ruber; 21 - stria terminalis; 22 - nucleus hypothalamicus; 23 - nucleus caudatus; 24 - кора полушария; 25 - cavum septi pellucidi; 26 - cornu anterius ventriculi lateralis; AA - capsula interna; a - tractus frontothalamicus; б - tractus frontopontinus; в - tractus corticonuclearis; г - tractus corticospinalis; д - tractus bulbothalamicus и spinothalamicus; e - tractus occipitotemporopontinus; ж - центральный слуховой тракт; з - центральный зрительный тракт
1. Corpus striatum, полосатое тело, состоит из двух не вполне отделенных друг от друга частей - nucleus caudatus и nucleus lentiformis.
A. Nucleus caudatus, хвостатое ядро, лежит выше и медиальнее nucleus lentiformis, отделяясь от последнего прослойкой белого вещества, называемой внутренней сумкой, capsula interna. Утолщенная передняя часть хвостатого ядра, его головка, caput nuclei caudati, образует латеральную стенку переднего рога бокового желудочка, задний же утонченный отдел хвостатого ядра, corpus et cauda nuclei caudati, тянется, как мы уже видели, назад по дну центральной части бокового желудочка; cauda заворачивается на верхнюю стенку нижнего рога. С медиальной стороны nucleus caudatus прилегает к зрительному бугру, отделяясь от него полоской белого вещества, stria terminalis. Спереди и снизу головка хвостатого ядра доходит до substantia perforata anterior, где она соединяется с nucleus lentiformis (с частью последнего, называемой put amen). Кроме этого широкого соединения обоих ядер с вентральной стороны, имеются еще тонкие полоски серого вещества, располагающиеся вперемежку с белыми пучками внутренней капсулы. Они послужили причиной названия - полосатое тело, corpus striatum (см. рис. 296).
Б. Nucleus lentiformis, чечевицеобразное ядро, залегает латерально от nucleus caudatus и зрительного бугра, отделенное от них capsula interna. На горизонтальном разрезе полушария медиальная поверхность чечевицеобразного ядра, обращенная к внутренней сумке, имеет форму угла с верхушкой, направленной к середине; передняя сторона угла параллельна хвостатому телу, а задняя - зрительному бугру. Латеральная поверхность немного выпукла и обращена к латеральной стороне полушария в области островка. Спереди и вентрально, как было уже указано, чечевицеобразное ядро сливается с головкой nucleus caudatus. На фронтальном разрезе чечевицеобразное ядро имеет форму клина, верхушка которого обращена в медиальную сторону, а основание - в латеральную. Чечевицеобразное ядро двумя параллельными белыми прослойками, laminae medullares, разделяется на три членика, из которых латеральный, темно-серого цвета, называется скорлупой, putamen (шелуха), а два медиальных, более светлых носят вместе название бледного шара, globus pallidus (см. рис. 296).
Отличаясь уже по своему макроскопическому виду, globus pallidus имеет также и гистологическую структуру, отличную от других частей полосатого тела. Филогенетически globus pallidus представляет более старое образование (paleostriatum), чем putamen и nucleus caudatus (neostriatum).
Ввиду всех этих особенностей globus pallidus в настоящее время выделяют в особую морфологическую единицу под названием pallidum, тогда как обозначение striatum оставляют только за putamen и nucleus caudatus. Вследствие этого термин "чечевицеобразное ядро" теряет свое прежнее значение и может быть употребляем только в чисто топографическом смысле, а вместо прежнего названия corpus striatum хвостатое и чечевицеобразное ядро именуют стриопаллидарной системой. Стриопаллидарная система представляет собой главную часть экстрапирамидной системы (см. ниже), а кроме того, она является высшим регулирующим центром вегетативных функций в отношении теплорегуляции и углеводного обмена, доминирующим над подобными же вегетативными центрами в hypothalamus.
2. Claustrum, ограда, представляет тонкую пластинку серого вещества, заложенную в области островка, между ним и putamen (см. рис. 297). От последнего она отделяется прослойкой белого вещества, capsula externa, а от коры островка подобной же прослойкой, носящей название capsula extrema.
3. Corpus amygdaloideum, миндалевидное тело, или epistriatum, расположено под putamen в переднем конце височной доли. Corpus amygdaloideum, по-видимому, относится к подкорковым обонятельным центрам. В нем оканчивается идущий из обонятельной доли и substantia perforata anterior пучок волокон, отмеченный при описании зрительного бугра под названием stria terminalis (см. рис. 297).
Белое вещество полушарий
Все пространство между серым веществом мозговой коры и вязальными узлами занято белым веществом. Оно состоит из большого количества нервных волокон, идущих в различных направлениях и образующих проводящие пути конечного мозга. Нервные волокна могут быть разделены на три системы: 1) ассоциационные, 2) комиссуральные и 3) проекционные волокна.
А. Ассоциационные волокна (рис. 298) связывают между собой различные участки коры одного и того же полушария. Они разделяются на короткие и длинные. Короткие волокна, fibrae arcudtae cerebri, связывают между собой соседние извилины в форме дугообразных пучков. Длинные ассоциационные волокна соединяют более отдаленные друг от друга участки коры. Таких пучков волокон существует несколько. Cingulum, пояс, пучок волокон, проходящий в gyrus fornicatus, соединяет различные участки коры gyrus cinguli как между собой, так и с соседними извилинами медиальной поверхности полушария. Лобная доля соединяется с нижней теменной долькой, затылочной долей и задней частью височной доли посредством fasciculus longitudindlis superior. Височная и затылочная доли связываются между собой через fasciculus longitudindlis inferior. Наконец, орбитальную поверхность лобной доли соединяет с височным полюсом так называемый крючковидный пучок, fasciculus uncindtus.
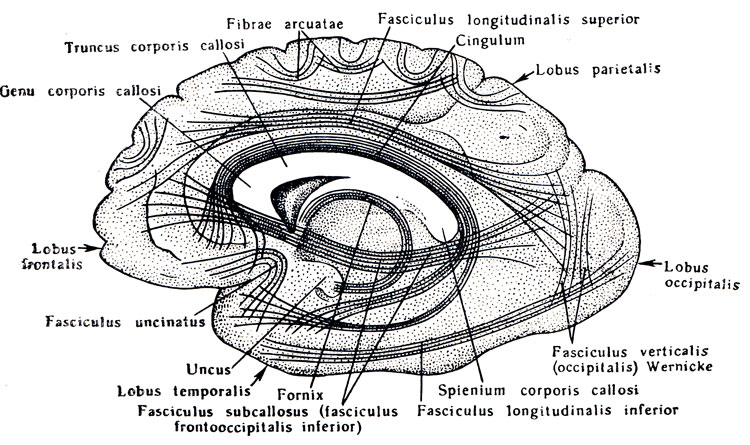
Рис. 298. Схематическое изображение ассоциационных путей мозговых полушарий
Б. Комиссуральные волокна, входящие в состав так называемых мозговых комиссур, или спаек, соединяют симметричные части обоих полушарий. Самая большая мозговая спайка - мозолистое тело, corpus callosum, которое связывает между собой части обоих полушарий, относящиеся к neencephalon.
Две мозговые спайки, commisstlra anterior et commissura fornicis, гораздо меньшие по своим размерам, относятся к rhinencephalon и соединяют: commissura anterior - обонятельные доли и обе парагиппокамповы извилины, commissura fornicis - аммоновы рога.
В. Проекционные волокна связывают мозговую кору частью с thalamus et corpora geniculata, частью с нижележащими отделами центральной нервной системы до спинного мозга включительно. Одни из этих волокон проводят возбуждения центростремительно, по направлению к коре, а другие, наоборот, центробежно.
Проекционные волокна в белом веществе полушария ближе к коре образуют так называемый лучистый венец, corona radidta, и затем главная часть их сходится во внутреннюю сумку, о которой упоминалось выше. Внутренняя сумка, cdpsula interna, как было указано, представляет слой белого вещества между nucleus lentiformis с одной стороны и хвостатым ядром и зрительным бугром - с другой. На фронтальном разрезе мозга внутренняя сумка имеет вид косо идущей белой полосы, продолжающейся в ножку мозга. На горизонтальном разрезе она представляется в форме угла, открытого в латеральную сторону (см. рис. 297), вследствие этого в capsula interna различают переднюю часть, crus anterius cap-sulae internae - между хвостатым ядром и передней половиной внутренней поверхности nucleus lentiformis, заднюю часть, crus posterius, между зрительным бугром и задней половиной чечевицеобразного ядра, и колено, genu capsulae internae, лежащее на месте перегиба между обеими частями внутренней сумки. Проекционные волокна по их длине могут быть разделены на следующие системы, начиная с самых длинных:
1. Trdctus corticospinal (pyramidalis), проводит двигательные волевые импульсы к мышцам туловища и конечностей. Начавшись от пирамидных клеток коры средней и верхней части передней центральной извилины и lobulus paracentralis, волокна пирамидного пути идут в составе лучистого венца, а затем проходят через внутреннюю капсулу, занимая передние две трети ее задней части, причем волокна для верхней конечности идут спереди волокон для нижней конечности. После этого они проходят через ножку мозга, crus cerebri, а оттуда через мост в продолговатый мозг.
2. Trdctus corticonuclear, проводящие пути к двигательным ядрам головных нервов. Начавшись от пирамидных клеток коры нижней части передней центральной извилины, они проходят через колено внутренней сумки и через ножку мозга, затем вступают в мост и, переходя на другую сторону, оканчиваются в двигательных ядрах противоположной стороны, образуя перекрест. Небольшая часть волокон оканчивается без перекреста.
Так как все двигательные волокна собраны на небольшом пространстве во внутренней сумке (колено и передние две трети задней ножки ее), то при повреждении их в этом месте наблюдается односторонний паралич (hemiplegia) противоположной стороны тела.
3. Trdctus corticopontini, пути от мозговой коры к ядрам моста. Они идут от коры лобной доли (tractus frontopontinus), затылочной (tractus occipitopontinus), височной (tractus temporopontine) и теменной (tractus parietopqntinus). В качестве продолжения этих путей из ядер моста идут волокна в составе средних ножек мозжечка в мозжечок. При помощи этих путей кора большого мозга оказывает тормозящее и регулирующее влияние на деятельность мозжечка.
4. Fasciculi thalamocortical et corticothaldmici, волокна от зрительного бугра к коре и обратно от коры к зрительному бугру. Из волокон, идущих от зрительного бугра, необходимо отметить так называемый путь покрышки, который является конечной частью чувствительного пути, направляющегося к центру кожного чувства в заднюю центральную извилину. Выходя из латерального ядра зрительного бугра, волокна этого пути проходят через заднюю часть внутренней капсулы, позади пирамидного пути. Место это было названо "чувствительным перекрестом", так как здесь проходят и другие чувствительные пути, а именно зрительный пучок Грациоле, radiatio optica, идущий от corpus geniculatum laterale и pulvinar зрительного бугра к зрительному центру в коре затылочной доли, затем слуховой пучок, radiatio acustica, направляющийся от corpus geniculatum mediale и нижнего бугорка четверохолмия к извилинам Гешля в височной доле, где заложен центр слуха. Зрительный и слуховой пучки занимают самое заднее положение в задней части внутренней капсулы.
При использовании материалов сайта активная ссылка обязательна:
http://anfiz.ru/ 'Анатомия и физиология человека'