Общие принципы строения нервной системы
Структурной единицей нервной системы является нейроцит - тело нервной клетки с отходящими от него отростками. Специфическая форма деятельности нейроцитов тесно связана с наличием многочисленных клеток нейроглии, которые, окружая нейроциты, выполняют опорную, изолирующую, трофическую, защитную и секреторную функции. Нейроциты крайне разнообразны по своим размерам, форме, количеству и длине отростков, наличие которых является характерной чертой их строения. В зависимости от количества отростков все нейроциты делят на три группы: униполярные - с одним отростком, биполярные - с друмя отростками, мулътиполярные - более двух отростков. В нервной системе человека наиболее многочисленны мультиполярные нейроциты. По своему строению и функциям отростки нейроцитов подразделяют на аксоны, или нейриты, и дендриты. Нейроциты имеют различное количество богато ветвящихся цендритов, функция которых заключается в восприятии раздражений из внешней и внутренней среды организма или от других нейроцитов и проведении нервного импульса к телу нейроцита. В отличие от дендритов нейрит проводит нервный импульс от тела нейроцита к другим нейроцитам или к исполнительным органам (мышцам, железам), причем все нейроциты имеют только один различной длины нейрит. Возбуждение (нервный импульс) в нейроците следует в направлении дендрит - тело нейроцита - аксон - следующий нейроцит или исполнительный орган.
Отростки нервных клеток покрыты оболочками, в связи со строением которых различают мякотные, или миелиновые, и безмякотные, или амиелиновые, нервные волокна. Мякотные нервные волокна состоят из отростка нервной клетки, который занимает в нервном волокне центральное положение и называется осевым цилиндром, или аксоном, тонкой наружной оболочки - аксолеммы и толстой, имеющей перехваты миелиновой оболочки (см. раздел Общие принципы строения анимальных периферических нервов, настоящего издания).
В зависимости от строения, функции и связей все нейроциты подразделяют на чувствительные, или рецепторные, вставочные, или ассоциативные, и двигательные, или эффекторные. Чувствительные нейроциты характеризуются наличием двух отростков: сравнительно длинного - дендрита и аксона. Окончания дендрита образуют специальный воспринимающий аппарат - рецептор, который чувствителен к минимальным количествам энергии раздражения и способен трансформировать ее в нервный импульс. Рецепторы специализированы к определенному виду раздражений (механические, температурные, химические и др.) и отличаются по своему строению.
Все рецепторы подразделяют на две группы: внешние и внутренние.
Внешние рецепторы (экстерорецепторы) воспринимают раздражения внешней среды. Внутренние рецепторы (интерорецепторы) воспринимают раздражения от внутренних органов (висцерорецепторы), а также от мышц, сухожилий, суставов (проприорецепторы).
Наиболее распространенной формой чувствительных нейроцитов являются псевдоуниполярные нейроциты спинномозговых и черепных ганглиев. Псевдоуниполярными эти клетки названы потому, что их нейрит и дендрит возникают из общего выпячивания тела клетки, что создает впечатление наличия одного отростка, который далее Т-образно делится, причем дендрит заканчивается на периферии рецептором, а конечные разветвления аксона вступают в контакт с нейроцитами центральной нервной системы. Нервный импульс в чувствительном нейроците следует в направлении рецептор - дендрит - тело клетки - нейрит - следующий нейрон.
Двигательные нейроциты получают нервный импульс только от других нейронов и передают его через концевые аппараты нейрита (эффекторы) тканям рабочих органов или другому двигательному нейроциту. Двигательные нейроциты разнообразны по форме, часто имеют большие размеры (до 130 мк) и длинный нейрит (до 1 м). Наиболее многочисленную группу нейроцитов составляют вставочные, посредством которых осуществляется контакт между чувствительными и двигательными нервными клетками. Контакт между нейроцитами носит название синапса. Синапсы состоят из концевого аппарата нейрита, синаптической щели, пресинаптической и постсинаптической мембран. Концевой аппарат нейритов имеет форму пуговок, бляшек или колечек, поверхность которых в области синапса покрыта пресинаптической мембраной. Синаптическая щель расположена между пресинаптической и постсинаптической мембранами. Внутри концевого аппарата нейритов находятся синаптические пузырьки, содержимым которых, как предполагают, является возбуждающий (симпатии) или тормозящий (ацетилхолин) медиатор, осуществляющий передачу возбуждения с нейроцита на нейроцит. Концевой аппарат нейрита может вступать в контакт с телом (аксо-соматический синапс), с дендрита-ми: их выпячиваниями - шипиками (аксо-дендритический) и аксонами (аксо-аксональный) второго нейроцита. Возможны также контакты между дендритами различных клеток - дендро-дендритический синапс. Концевой аппарат одного нейрита вступает в контакт с телами и дендритами многих нейроцитов и количество образованных им синапсов может быть очень большим (до 10 000), что свидетельствует о конструктивной и функциональной сложности межнейронных отношений. В области синапсов прерывается путь прохождения нервного импульса с одного нейроцита на другой, и здесь осуществляется регуляция проведения возбуждения. Одним из важнейших свойств синапсов является одностороннее проведение возбуждения с концевых разветвлений нейрита одной клетки на тело или дендриты другой. Это свойство синапсов обеспечивает направленность прохождения нервных импульсов в нервной системе и лежит в основе материального субстракта нервной деятельности - рефлекторных дуг. Синаптический аппарат обеспечивает динамическую связь между отдельными нейронами и определяет единство нервной системы.
В 1862 г. И. М. Сеченов писал: "Все акты сознательной и бессознательной жизни по способу происхождения суть рефлексы". Осуществление рефлекторных актов или рефлексов - основное и специфическое проявление деятельности нервной системы. Рефлекс - реакция организма на изменения внешней и внутренней среды. Цепочка нейроцитов, обеспечивающая рефлекторный акт, носит название рефлекторной дуги. Простая рефлекторная дуга (рис. 190) состоит: из рецептора, воспринимающего энергию внешнего или внутреннего раздражения и трансформирующего его в форму нервного импульса, дендрита чувствительной клетки, ее тела, нейрита чувствительной клетки, синапсов с дендритами или телом двигательной клетки, нейрита двигательной клетки, двигательных окончаний в рабочих органах. В большинстве случаев рефлекторная дуга между чувствительным и двигательным нейроцитами содержит несколько вставочных или ассоциативных клеток. Замыкание рефлекторных дуг происходит в нервных центрах. Нейроцит, приносящий нервный импульс в нервный центр, носит название центростремительного, или афферентного. Нейроцит, осуществляющий ответную реакцию (двигательную или секреторную) благодаря проведению нервного импульса от нервного центра к рабочим органам, называется центробежным, или эфферентным. Переключение возбуждения с центростремительного нейроцита на центробежный осуществляется ассоциативными нейроцитами.
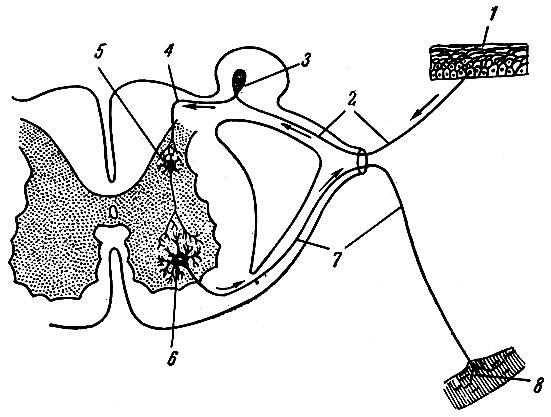
Рис. 190. Схема рефлекторной дуги. 1 - нервное окончание чувствительного нейроцита в коже; 2 - периферический отросток чувствительного нейроцита; 3 - спинномозговой узел с псевдоуниполярной клеткой; 4 - центральный отросток чувствительного нейроцита; 5 - вставочный нейроцит; 6 - двигательная клетка переднего рога; 7 - нейрит двигательной клетки; 8 - нервное окончание в мышце
В настоящее время принято считать, что замыкающим звеном любой рефлекторной дуги является звено обратной связи, или обратной афферентации, благодаря которой в соответствующие нервные центры поступают нервные импульсы, сигнализирующие о результатах совершенного действия. Звено обратной связи, т. е. обратной афферентации, обеспечивает приспособительный характер рефлекторной деятельности.
Тела клеток афферентных нейронов расположены вне центральной нервной системы, в спинномозговых и черепных узлах (тройничный узел, верхний и нижний узлы блуждающего и языко-глоточного нервов и др.). Вне центральной нервной системы находятся эфферентные нейроциты автономной нервной системы. Ассоциативные (вставочные) нейроциты и тела клеток эфферентных нейроцитов соматической нервной, системы входят в состав серого вещества головного и спинного мозга.
На разрезах головного и спинного мозга даже невооруженным глазом видно, что все отделы центральной нервной системы образованы серым и белым веществом. Серое вещество представляет собой скопление нейроцитов, безмякотных и тонких мякотных волокон и нейроглии, которой примерно в 10 раз больше, чем нейроцитов. Белое вещество образовано мякотными нервными волокнами и клетками нейроглии. В сером веществе различают три типа пространственного взаимоотношения нейроцитов: сетевидный, ядерный и корковый.
Сетевидный тип взаимоотношения нейронов является филогенетически наиболее древним и характерен для строения сетчатого образования (formatio reticularis) центральной нервной системы. Сетчатое образование представляет собой центрально расположенный диффузный столб нейронов особого строения, который непрерывно прослеживается на протяжении спинного и стволовой части головного мозга. Нейроциты сетчатого образования характеризуются многочисленными длинными прямыми слабо разветвленными дендритами, а их нейриты не образуют компактных пучков и рассеяны на значительной площади. В сетчатом образовании имеются участки некоторой концентрации нейроцитов.
Ядерный тип организации нейроцитов характеризуется резко выраженной концентрацией этих элементов с образованием пространственно замкнутых клеточных скоплений различной формы и величины. Образующие их нейроциты в основном имеют густо ветвящиеся дендриты, а нейриты объединяются в компактные пучки. К образованиям такого типа относятся двигательные и чувствительные ядра спинного и головного мозга и ядра, преимущественно осуществляющие переключательную функцию (нижние оливы, ядра нежного и клиновидного канатиков и др.).
Для коркового типа организации нейроцитов характерно их правильное послойное распределение с образованием структуры слоистого типа. Такое строение имеют кора полушарий большого мозга и мозжечка и некоторые образования других отделов центральной нервной системы. Наиболее сложные взаимоотношения нейроцитов наблюдаются в коре полушарий большого мозга - высшем отделе центральной нервной системы, под контролем которой протекает деятельность всех нижележащих отделов стволовой части головного и спинного мозга, где также вышележащие отделы оказывают доминирующее влияние на нижележащие.
Функции связи между группами нейроцитов спинномозговых и черепных узлов, сетчатых, ядерных и корковых структур осуществляются нервными волокнами. Одинаковые по происхождению и функции нервные волокна, объединенные в пучки различной плотности, которые соединяют между собой посредством синапсов функционально схожие скопления нервных клеток - нервные центры, называют проводящими путями. Каждый проводящий путь представляет собой комплекс последовательно соединенных нейроцитов в виде цепочки, составляющих рефлекторные дуги различной сложности (двух-, трех- и многонейронные), которые являются функциональной основой деятельности нервной системы. Рефлекторные акты, связанные с деятельностью подкорковых ядер, ствола мозга и спинного мозга, протекают по врожденным наследственным нервным путям, свойственным всем представителям данного вида животных. Такие рефлекторные реакции называют безусловпорефлекторными. Их значение заключается в сохранении особи или вида, регуляции внутренней среды организма. С функцией коры больших полушарий связана условнорефлекторная деятельность, образование нервных связей на основе индивидуального опыта, рефлексов, которые создаются в процессе жизни индивидума, в результате воздействия на организм множества различных раздражений.
В коре больших полушарий осуществляется анализ и синтез различных раздражений (сигналов) внешнего мира, что составляет первую сигнальную систему, наличие которой характерно как для человека, так и для высших животных (И. П. Павлов). У человека в результате его социального развития и трудовой деятельности появилась высшая система сигнализации, связанная с восприятием речи (письменной, устной), что составляет свойственную только деятельности коры большого мозга человека вторую сигнальную систему.
Спинной мозг
Развитие спинного мозга
Все отделы центральной нервной системы человека развиваются из нервной трубки (см. раздел Общая неврология, настоящего издания), которая в результате митотического деления клеток становится многослойной. В этот период в ней можно выделить три слоя: наружный, или краевой, внутренний, или эпендимный, и средний, или мантийный (плащевой). Из клеток внутреннего слоя развиваются эпендимные клетки, которые выстилают полости головного и спинного мозга (центральный канал спинного и желудочки головного мозга). Из клеток мантийного слоя образуются нейробласты и спонгиобласты; первые развиваются в нервные клетки, а вторые - в клетки нейроглии.
В нервной трубке выделяют дорсальную, боковые и вентральную пластинки.
Деление клеток в различных отделах туловищного конца нервной трубки протекает с неодинаковой интенсивностью. Наиболее резко увеличивается количество клеток в ее боковых отделах, где на внутренней поверхности утолщенной боковой стенки появляется пограничная борозда, sulcus limitans, разделяющая боковую стенку на две пластинки: дорсо-латеральную, lamina dorsolateralis, и венгрю-латеральную, lamina ventrolateralis. Из клеток вентро-латеральной пластинки далее образуются передние столбы серого вещества спинного мозга, а из клеток дорсо-латеральной пластинки формируются его задние столбы. Из нейробластов передних столбов дифференцируются корешковые клетки двигательных ядер передних столбов серого вещества, а их отростки образуют брюшные корешки спинного мозга. Нейробласты задних столбов дифференцируются в пучковые и внутренние нервные клетки ядер задних столбов и промежуточной зоны. Нейриты этих клеток проходят кнаружи в краевой слой и образуют большую часть белого вещества спинного мозга, т. е. его проводящие пути.
Неравномерный рост боковых пластинок по сравнению с вентральной и дорсальной приводит к изменению формы нервной трубки. Она прогибается в области донной и покровной пластинок с образованием двух продольных борозд, которые делят спинной мозг на две симметричные половины. Утолщение стенок и изменение формы спинного мозга сопровождается резким сужением его полости - центрального канала.
Строение спинного мозга
Спинной мозг, medulla spinalis, представляет собой неравномерный по толщине сдавленный спереди назад цилиндрической формы тяж длиной в среднем 45 см у мужчин и 41-42 см у женщин (рис. 191). Возле верхнего края атланта спинной мозг без резких границ переходит в продолговатый мозг, а на уровне II поясничного позвонка заканчивается мозговым конусом, conus medullaris, тонкая вершина которого (диаметр до 2 мм) продолжается в конечную нить, filum terminate, которая прикрепляется ко II копчиковому позвонку.
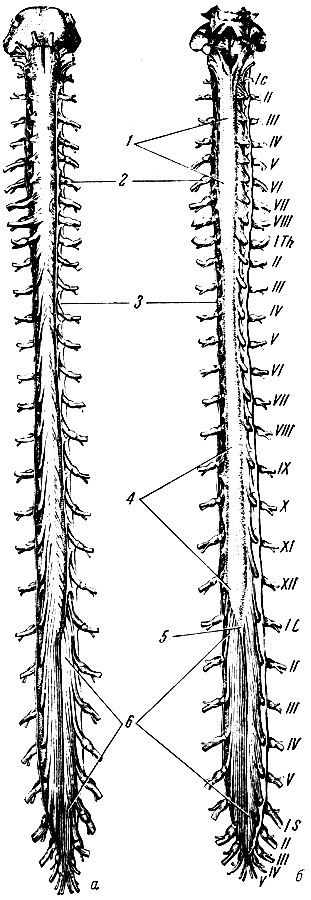
Рис 191. Спинной мозг. Твердая и паутинная оболочки разрезаны. Сосудистая оболочка снята. Римскими цифрами обозначен порядок расположения шейных, грудных, поясничных и крестцовых спинномозговых нервов. а - вид спереди (вентральная поверхность); б - вид сзади дорсальная поверхность); 1 - шейное утолщение; 2 - спинномозговой узел; 3 - твердая оболочка спинного мозга; 4 - поясничное утолщение; 5 - мозговой конус; 6 - конский хвост
Толщина и форма спинного мозга на всем его протяжении не одинаковы. В шейном и поясничном отделах имеются два веретенообразной формы утолщения: шейное, intumescentia cervicalis, и поясничное, intumescentia lumbalis, которые соответствуют местам отхождения корешков нервов к верхней и нижней конечностям. Поперечный диаметр спинного мозга в области шейного утолщения равен 13-14 мм, поясничного - 12 мм, а в грудном отделе - 10 мм.
На передней поверхности спинного мозга имеется глубокая передняя срединная щель, fissura mediana anterior, которая вместе с задней срединной бороздой, sulcus medianus posterior, идущей продольно по его задней поверхности, делит мозг на две симметричные половины. На поверхности каждой половины спинного мозга находятся слабо выраженные передняя и задняя боковые борозды, sulci laterales anteriores et posteriores, представляющие собой место выхода соответственно передних и входа задних корешков. Кроме этих борозд, в шейном и верхнегрудном отделах спинного мозга между sulcus medianus posterior и sulcus lateralis posterior имеется задняя промежуточная борозда, sulcus, intermedius posterior. Отмеченные борозды делят белое вещество спинного мозга на обособленные пучки нервных волокон, вызываемые канатиками, funiculi medullae spinalis, которые хорошо видны на поперечных срезах. Самый узкий передний канатик, funiculus anterior, расположен между fissura mediana anterior и sulcus lateralis anterior. Боковой, канатик, funiculus lateralis, находится между боковыми бороздами, а задний канатик, funiculus posterior, ограничен sulcus lateralis posterior и sulcus medianus posterior. Кроме того, в шейном и верхнегрудном отделах задний канатик подразделяется задней промежуточной бороздой на два пучка: медиально расположенный тонкий пучок, fasciculus gracilis, и латеральный - клиновидный, fasciculus cuneatus.
По бокам спинного мозга видны два вертикальных ряда (передний и задний) пучков нервных волокон, которые называют корешками спинномозговых нервов (рис. 192). Передний брюшной корешок, radix ventralis, преимущественно образован аксонанами двигательных клеток передних рогов спинного мозга. Задний, спинной корешок, radix dorsalis, составляют аксоны чувствительных клеток спинномозговых узлов. Брюшной корешок выходит из передней боковой борозды спинного мозга и является по функции двигательным. Спинной корешок вступает в боковую заднюю борозду и является чувствительным. Соединяясь, брюшной и спинной корешки образуют ствол спинномозгового нерва, который по составу волокон является смешанным. Возле места образования спинномозгового нерва на спинном корешке имеется утолщение - спинномозговой узел, ganglion spinale, представляющий собой скопление псевдоуниполярных чувствительных нервных клеток. Дендриты этих клеток окончиваются на периферии рецепторными аппаратами, а нейриты составляют спинные корешки, проникающие в спинной мозг. Часть образующих их волокон вступает в контакт с клетками спинного мозга, образуя синапсы, а другая часть волокон идет к ядрам продолговатого мозга. Различают 31 пару корешков: 8 шейных, 12 грудных, 5 поясничных, 5 крестцовых и 1 копчиковый.
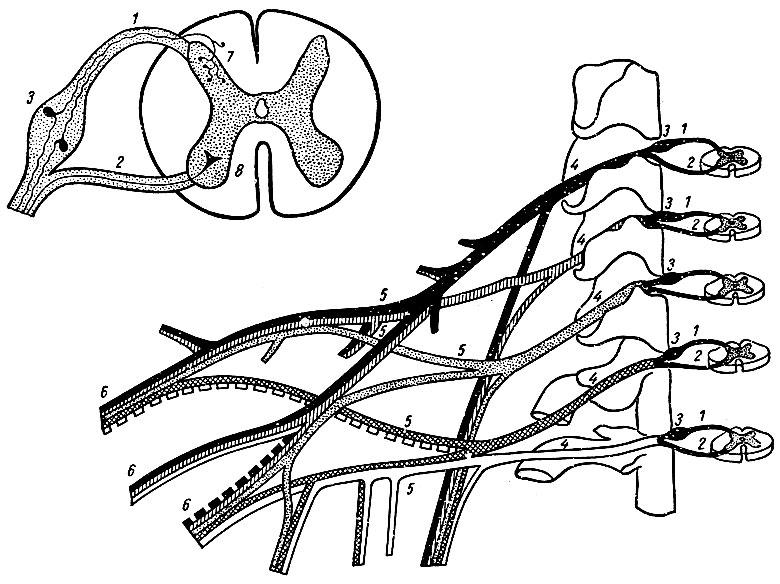
Рис. 192. Элементы периферической нервней системы (схема). 1 - спинной корешок; 2 - брюшной корешок; 3 - спинномозговой узел; 4 - первичные стволы нервного сплетения; 5 - нервное сплетение; 6 - вторичные стволы сплетения; 7 - задний рог; 8 - передний рог
Корешки спинномозговых нервов отличаются по своей толщине. Обычно спинные корешки толще брюшных, так как имеют большее количество нервных волокон. Исключение представляет первая пара шейных нервов, брюшные корешки которых толще спинных. Брюшные и спинные корешки, а также соответствующие им спинномозговые узлы наиболее крупны в области шейного и поясничного утолщений спинного мозга.
Между соседними корешками можно наблюдать межкорешковые связи, т. е. переход волокон из состава одного корешка в другой. Чаще такие связи бывают между спинными корешками, реже - между брюшными. Они особенно многочисленны в корешках, из которых образуются нервы конечностей. Наличие межкорешковых связей является одним из факторов, определяющих индивидуальную изменчивость в строении периферических нервов.
Ввиду того что уже с 4 месяцев у плода наблюдается отставание в росте спинного мозга от роста позвоночного столба, нижней границей спинного мозга взрослого становится уровень II поясничного позвонка. Поэтому наблюдается несоответствие между уровнями расположения сегментов, позвонками и межпозвонковыми отверстиями. Так, например, на уровне тела VII шейного позвонка располагается I грудной сегмент, на уровне X грудного позвонка - I поясничный сегмент, на уровне тела I поясничного позвонка - I крестцовый сегмент. В соответствии с этим корешки по отношению к спинному мозгу располагаются под различными углами и имеют разную длину. В шейном отделе спинного мозга направление корешков близко к горизонтальному, в грудном отделе угол становится все более острым, а у поясничных и крестцовых корешков направление почти отвесное. Поясничные, крестцовые и копчиковые корешки (начиная со II поясничного сегмента) на пути к соответствующим межпозвонковым отверстиям образуют крупный пучок, окружающий снаружи конечную нить, который называется конским хвостом, cauda equina.
Внутреннее строение спинного мозга, как и других отделов центральной нервной системы, изучают на серии поперечных и продольных срезов. На поперечных срезах видно, что спинной мозг образован серым и белым веществом, причем серое вещество занимает центральное положение. Передняя срединная щель и соединительнотканная задняя срединная перегородка, septum medianum posterius, проникают глубоко в мозговое вещество так, что между двумя симметричными половинами спинного мозга остается узкий участок мозговой ткани, в центре которой расположен центральный канал, canalis centralis. Являясь остатком полости первичной нервной трубки, центральный канал прослеживается на всем протяжении спинного мозга. В верхних шейных сегментах он принимает форму сагиттальной щели, которая сообщается с IV желудочком мозга. В области conus medullaris канал смещается ближе к задней срединной борозде и заканчивается небольшим расширением - конечным желудочком, ventriculus terminalis.
Серое вещество спинного мозга, substantia grisea, на поперечных срезах напоминает по форме бабочку или букву "Н" и на всем протяжении имеет вид двух неправильной формы колонн серых столбов, columnae griseae, соединенных между собой узкой перемычкой - серой спайкой, commissura grisea, которая подразделяется центральным каналом на две серые спайки: переднюю, commissura grisea anterior, и заднюю, commissura grisea posterior. Серые столбы подразделяются на передний и задний, columnae griseae anterior et posterior. Кроме того, между VIII шейным и II или III поясничным сегментами имеется боковой выступ серого вещества, который образует боковые столбы, columnae griseae laterales.
На поперечных срезах выступы серого вещества принято называть рогами. Наиболее массивны передние рога, cornua anteriores, передний конец которых расширен в шейном и пояснично-крестцовом отделах спинного мозга. Задние рога, cornua posteriores, значительно уступают в своей величине передним. Различают основание заднего рога, переходящее в сужение - его шейку, cervix cornus posterioris, которая кзади, расширяясь, образует головку заднего рога, caput cornus posterioris, заканчивающуюся верхушкой, apex cornus posterioris. На верхушке заднего рога находится студенистое вещество, substantia gelatinosa, к которому примыкает узкий слой губчатой зоны, zona spongiosa. Верхушку заднего рога окружает зона белого вещества, zona terminalis, образованная центральными отростками клеток спинальных ганглиев.
Между передними и задними рогами находится промежуточное вещество, substantia intermedia, центральная часть которого, substantia intermedia centralis, окружает центральный канал, а ее наружное продолжение - боковое промежуточное вещество, substantia intermedia lateralis, оканчивается боковым рогом на протяжении между VIII шейным и II или III поясничными сегментами.
Между задними и боковыми столбами в белом веществе имеются отдельные мелкие островки или тяжи серого вещества, называемые в совокупности сетчатым образованием спинного мозга, formatio reticularis.
Нервные клетки серого вещества образуют различные по форме и величине скопления - ядра спинного мозга (рис. 193). В задних столбах расположены соматически-чувствительные ядра, в передних - соматически-двигательные, а в боковых столбах - ядра вегетативной нервной системы.
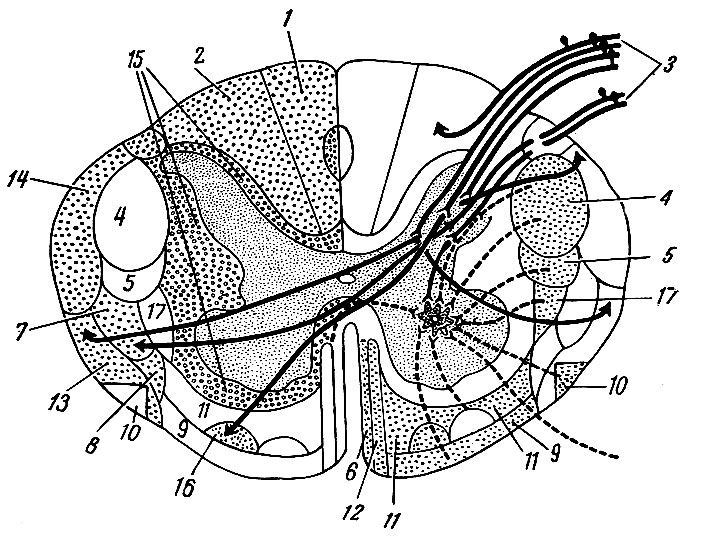
Рис. 193. Схема проводящих путей спинного мозга (поперечный срез). Левая половина среза показывает восходящие системы волокон, правая половина топографию нисходящих систем волокон. 1 - тонкий пучок; 2 - клиновидный пучок; 3 - спинной корешок; 4 - боковой корковоспинномозговой путь (пирамидный); 5 - красноядерно-спинномозговой путь; 6 - покрышечно-спинномозговой путь; 7 - боковой спинно-бугорный путь; 8 - спинно-покрышечный путь; 9 - преддверно-спинномозговой путь; 10 - оливо-спинномозговой путь; 11 - сетчато-спинно-мозговой путь; 12 - передний корково-спинномозговой путь (пирамидный); 13 - передний спинно-мозжечковый путь; 14 - задний спинно-мозжечковый путь; 15 - собственные пучки; 16 - передний спинно-бугорный путь; 17 - бугорно-спинномозговой путь
Соматически-чувствительные ядра задних столбов образованы пучковыми и вставочными клетками. Пучковые клетки различны по величине. Их нейриты образуют пучки волокон, которые заканчиваются как в ядрах спинного, так и головного мозга, являясь вторыми нейроцитами проводящих путей. Вставочные клетки осуществляют связь между клетками серого вещества спинного мозга. Из крупных пучковых клеток образовано грудное ядро, nucleus thoracicus. Оно расположено в области основания заднего рога. Нейриты клеток этого ядра входят в состав бокового канатика белого вещества той же стороны спинного мозга. В центре заднего рога находится собственное ядро, nucleus proprius. Оно также образовано пучковыми клетками, но их нейриты переходят в боковой канатик противоположной стороны. Как грудное, так и собственное ядра спинного мозга, получив нервные импульсы от центральных отростков клеток спинальных ганглиев, переносят их в ядра головного мозга. Нервные клетки губчатой зоны, студенистого вещества и вставочные клетки осуществляют связь между чувствительными клетками спинальных ганглиев и двигательными клетками передних рогов, замыкая местные рефлекторные Дуги.
Соматически-двигательные ядра передних рогов образованы крупными корешковыми клетками, отростки которых формируют основную часть волокон передних корешков и осуществляют двигательную иннервацию поперечнополосатой мускулатуры.
Белое вещество спинного мозга состоит преимущественно из продольно расположенных нервных волокон, составляющих передние, задние и боковые канатики. В образовании канатиков участвуют нейриты чувствительных клеток спинномозговых узлов, отростки клеток серого вещества спинного мозга и отростки клеток различных отделов головного мозга. Кроме канатиков, из белого вещества образована передняя белая спайка, commissura alba anterior, и состоящая из небольшого количества волокон задняя белая спайка, commissura alba posterior. В свою очередь каждый канатик спинного мозга образует несколько различных по функциональному значению пучков нервных волокон - проводящих путей спинного мозга.
Проводящие пути спинного мозга подразделяются на короткие и длинные. Короткие проводящие пути, ассоциационные: это пути собственного аппарата спинного мозга, посредством которых осуществляется одна из основных его функций - рефлекторная. С наличием длинных проводящих путей связана вторая основная функция спинного мозга - проводниковая. Длинные проводящие пути спинного мозга разделяются на восходящие - чувствительные, которые проводят нервные импульсы в головной мозг, и нисходящие - двигательные, следующие от различных центров головного мозга к спинному. Проводящие пути имеют определенную строгую локализацию (см. рис. 193).
К коротким проводящим путям относятся собственные пучки, fasciculi proprii anterior, posterior et lateralis; окружающие в виде узкой полосы серое вещество каждой половины спинного мозга. Они образованы отростками рассеянных в сером веществе спинного мозга пучковых клеток, клеток студенистого вещества и губчатой зоны и осуществляют связь между различными сегментами, а также между чувствительными клетками спинальных ганглиев и двигательными клетками передних рогов, входя в состав местных рефлекторных дуг, посредством которых осуществляются врожденные, безусловные рефлексы.
Длинные проводящие пути являются аппаратом двусторонней связи головного и спинного мозга, посредством которого осуществляются сложные рефлекторные акты.
Задние канатики состоят из волокон, которые являются центральными отростками чувствительных клеток спинномозговых узлов, и собственного заднего пучка. Вступив в состав задних корешков в краевую зону спинного мозга, каждое волокно делится Т-образно на короткую нисходящую и длинную восходящую ветви. Коллатерали этих ветвей образуют большое число синапсов с рассеянными пучковыми клетками, после чего длинные восходящие ветви проходят к ядрам продолговатого мозга. Медиальную часть заднего канатика занимает тонкий пучок, fasciculus gracilis. Он образован отростками клеток спинальных ганглиев 19 нижних сегментов спинного мозга. Латерально расположен клиновидный пучок, fasciculus cuneatus, который образован отростками клеток спинальных ганглиев 12 верхних сегментов спинного мозга. Волокна тонкого и клиновидного пучков образуют синапсы с клетками тонкого и клиновидного ядер продолговатого мозга. Они являются вторыми нейроцитами пути сознательной проприоцептивной чувствительности (мышечно-сустав-ного чувства), а также вибрационной, тактильной и дискриминационной чувствительности (способность воспринимать раздельно два одновременно нанесенных раздражения).
Боковые канатики образуют: 1) нейриты нейроцитов чувствительных ядер спинного мозга, которые проводят нервные импульсы от низших центров к высшим (вторые нейроциты афферентных проводящих путей); 2) нейриты эфферентных нейроцитов ядер стволовой части мозга и коры полушарий головного мозга, по которым нервные импульсы следуют от высших центров к низшим (первые нейроциты эфферентных путей).
Афферентные пути бокового канатика образуют следующие тракты.
1. Задний спинно-мозжечковый путь, tractus spinocerebellars posterior, составляют нейриты грудного ядра заднего рога той же стороны, которые образуют пучок волокон, расположенных по периферии задней части боковых столбов.
2. Передний спинно-мозжечковый путь, tractus spinocerebellars anterior, формируют нейриты медиального промежуточного ядра преимущественно противоположной стороны. Пучок расположен кпереди от предыдущего. Первыми нейроцитами спинно-мозжечковых путей являются чувствительные клетки спинномозговых узлов, которые проводят нервные импульсы от проприорецепторов мышц, сухожилий, суставов, в связи с чем эти пути выполняют роль бессознательной координации движений.
2. Спинно-покрышковый путь, tractus spinotectalis, является частью волокон переднего спинно-мозжечкового пути, которые заканчиваются не в мозжечке, а в буграх четверохолмия.
4. Боковой спинно-бугорный путь, tractus spinothalamics lateralis, представляет собой крупный пучок волокон, расположенный кнутри от переднего спинно-мозжечкового пути. Его образуют нейриты клеток собственного ядра заднего рога противоположной стороны. Первыми нейроцитами этого пути являются чувствительные клетки спинномозговых узлов, которые проводят в собственное ядро экстероцептивные импульсы (болевые, температурные, тактильные), а нейриты клеток собственного ядра (вторые нейроциты) несут эти импульсы к зрительным буграм промежуточного мозга. Часть волокон бокового спинно-бугорного пути после перекреста в передней белой спайке спинного мозга переходит в передний канатик.
Эфферентными путями бокового канатика считают следующие.
1. Боковой пирамидный или корково-спинномозговой путь, tractus руrаmidalis seu corticospinalis lateralis, состоит из нейритов пирамидных клеток коры полушарий головного мозга, которые после перекреста в продолговатом мозге образуют крупный пучок волокон бокового канатика. Книзу он уменьшается в размерах за счет посегментных окончаний его волокон на двигательных клетках передних рогов. Боковой пирамидный пучок расположен кнутри от заднего спинно-мозжечкового пути и является проводником импульсов сознательных целенаправленных движений.
2. Красноядерно-спинномозговой путь, tractus rubrоspinalis, находится кпереди от бокового пирамидного пути и кнутри от переднего спинно-мозжечкового пути. Этот путь образуют нейриты красного ядра среднего мозга, которое является важнейшим звеном так называемой экстрапирамидной системы. Волокна tractus rubrospinalis заканчиваются на клетках передних рогов и осуществляют бессознательную регуляцию движений и мышечного тонуса.
3. Сетчато-спинномозговой путь, tractus reticulospinalis, расположен между передним и боковым пирамидными путями. Он образован клетками formatio reticularis мозгового ствола противоположной стороны и заканчивается посегментно на двигательных клетках передних рогов. Выполняет функцию бессознательной регуляции движений.
4. Оливо-спинномозговой путь, tractus olivоspinalis, расположен кпереди от переднего спинно-мозжечкового пути. Его образуют нейриты клеток сливы продолговатого мозга. По этому тракту проходят импульсы от промежуточного центра равновесия (оливы) к двигательным клеткам передних рогов. Непосредственно к серому веществу в боковом канатике прилегает собственный пучок спинного мозга.
Передние канатики образованы главным образом нейритами эфферентных нейроцитов мозгового ствола и коры полушарий головного мозга, которые передают нервные импульсы от высших центров к низшим.
1. Покрышечно-спинномозговой путь, tractus tectospinalis, занимает медиальную часть переднего канатика. Он образован нейритами клеток ядер четверохолмия, которые оканчиваются в ядрах передних рогов, и является путем проведения импульсов защитных двигательных рефлекторных реакций на внезапные световые и звуковые-раздражения.
2. Передний пирамидный, или корково-спинномозговой, путь, tractus pyramidalis seu cortico spinalis anterior, лежит латеральнее tractus tectospinalis. Он представляет собой узкий пучок волокон, которые являются нейритами пирамидных клеток коры полушарий головного мозга, оканчивающихся в двигательных ядрах передних рогов. Волокна переднего пирамидного пути не совершают перекреста в продолговатом мозге, но перед вступлением в передний рог через переднюю белую спайку переходят на противоположную сторону. Передний пирамидный путь является путем проведения импульсов сознательных, произвольных движений.
3. Передний сшшно-бугорный путь, tractus spinothalamics anterior, расположен кнаружи от переднего пирамидного пути (см. боковой спино-бугорный путь).
4. Преддверно-спинномозговой путь, tractus vestibulospinalis, занимает самое латеральное положение в переднем канатике. Он образован нейритами клеток вестибулярных ядер стволовой части мозга, которые оканчиваются посегментно в ядрах передних рогов и проводят импульсы к спинному мозгу от вестибулярного аппарата. К серому веществу в переднем канатике непосредственно прилегает собственный пучок спинного мозга.
Оболочки спинного мозга
Спинной и головной мозг покрыты тремя соединительнотканными оболочками, которые развиваются из окружающей мозговую трубку мезодермы (рис. 194). Снаружи расположена твердая мозговая оболочка, dura mater, образованная плотной волокнистой соединительной тканью. Глубже находится паутинная оболочка, arachnoidea, которая представляет собой тонкий, бессосудистый листок рыхлой волокнистой соединительной ткани. Непосредственно к веществу мозга прилегает сосудистая оболочка, pia mater, которая образована также волокнистой соединительной тканью, но в отличие от паутинной оболочки содержит сети кровеносных сосудов мозга. Все три оболочки в виде единого, непрерывного футляра покрывают спинной и головной мозг.
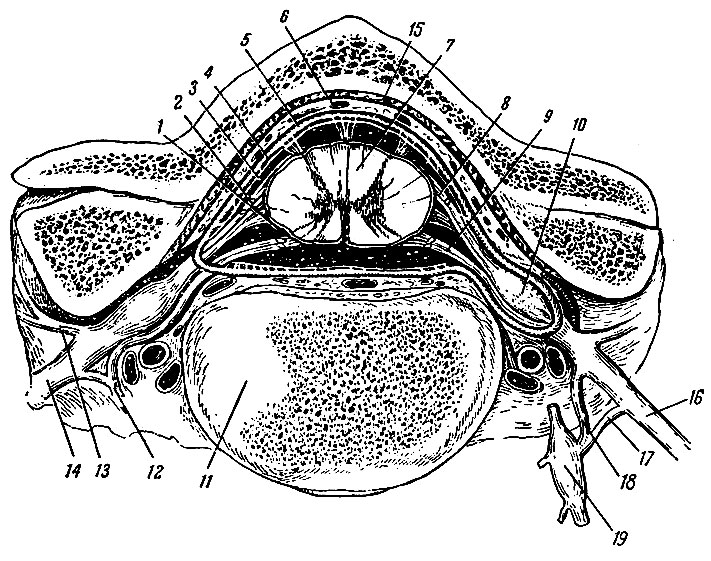
Рис. 194. Оболочки спинного мозга. 1 - мягкая оболочка; 2 - паутинная оболочка; 3 - подпаутинное пространство; 4 - субдуральное пространство; 5 - твердая оболочка; 6 - эпидуральное пространство; 7 - спинной мозг; 8 - спинной корешок; 9 - брюшной корешок; 10 - спинномозговой узел; 11 - позвонок; 12, 18 - белые соединительные ветви; 13 - задние ветви спинномозговых нервов; 14 - брюшные ветви спинномозговых нервов; 15 - надкостница; 16 - спинномозговой нерв; 17 - серые соединительные ветви; 19 - узел симпатического ствола
Твердая оболочка спинного мозга, dura mater spinalis, представляет собой мешок цилиндрической формы, который свободно покрывает спинной мозг. В области большого затылочного отверстия она плотно сращена с его краем, а на уровне II поясничного позвонка заостряется и переходит в нить твердой оболочки спинного мозга, filum durae matris spinalis. Она доходит до II поясничного позвонка, где и прикрепляется. Между dura mater и надкостницей позвоночного канала, которую называют наружным листом твердой оболочки, имеется значительное по объему эпидуральное пространство, cavum epjdurale, заполненное жировой клетчаткой и венозным сплетением. В эпидуральном пространстве также проходят покрытые отрогами твердой оболочки корешки спинномозговых нервов. Эти отроги имеют вид рукава и обычно содержат оба корешка. Отроги твердой оболочки, ее нить и фиброзные пучки волокон, которые соединяют ее переднюю поверхность с задней продольной связкой позвоночника, фиксируют dura mater в позвоночном канале. Между внутренней поверхностью твердой оболочки, которая покрыта эндотелием, и глубже расположенной паутинной оболочкой имеется узкое субдуральное пространство, cavum subdurale.
Паутинная оболочка спинного мозга, arachnoidea spinalis, повторяет форму dura mater и местами прочно с ней связана соединительнотканными волокнами. Образующий ее тонкий, прозрачный листок с обеих сторон покрыт эндотелием. Между паутинной и сосудистой оболочками имеется, широкое подпаутинное пространство, cavum subarachnoidale, заполненное спинномозговой жидкостью, liquor cerebrospinal. Это пространство особенно широко в области cauda equina спинного мозга. Краниально подпаутинное пространство спинного мозга непосредственно продолжается в одноименное пространство головного мозга.
Паутинная и сосудистая оболочки соединены между собой тонкими соединительнотканными нитями, которые пронизывают подпаутинное пространство. Спинной мозг связан с твердой оболочкой симметрично расположенными по бокам зубчатыми связками, lig. denticulatum.
Сосудистая оболочка спинного мозга, pia mater spinalis, непосредственно прилежит к мозговому веществу и образует расположенную в передней срединной щели переднюю продольную перегородку, septum longitudinale anterior. Сосудистая оболочка вместе с мозговыми сосудами проникает в мозговую ткань.
Головной мозг
Общий обзор строения головного мозга
Головной мозг, encephalon, состоит из трех основных частей: полушарий большого мозга, hemispherium cerebri, мозгового ствола, truncus cerebri, мозжечка, cerebellum. Выпуклую верхне-боковую поверхность большого мозга, fades superlateralis cerebri, образуют только полушария головного мозга. Его несколько уплощенная, более сложная по рельефу нижняя поверхность, fades inferior cerebri, представлена как нижними поверхностями полушарий, так и некоторыми отделами мозгового ствола и мозжечка (рис. 195). Оба полушария разделены продольной щелью большого мозга, fissura longitudinalis cerebri, в глубине которой можно видеть медиальные поверхности каждого полушария, fades medialis cerebri, и соединяющую их спайку - мозолистое тело, corpus callosum.
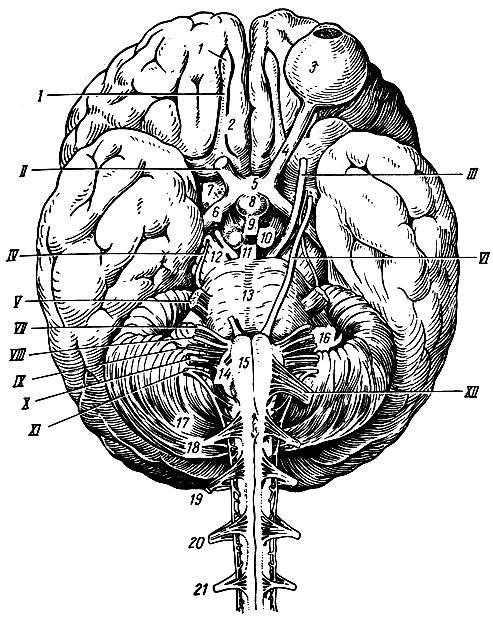
Рис. 195. Нижняя поверхность головного мозга. I - обонятельный путь; II - зрительный нерв; III - глазодвигательный нерв; IV - блоковый нерв; V - тройничный нерв; VI - отводящий нерв; VII - лицевой нерв; VIII - преддверно-слуховой нерв; IX - языко-глоточный; X - блуждающий нерв; XI - добавочный нерв; XII - подъязычный нерв; 1 - обонятельная луковица; 2 - обонятельный путь; 3 - левое глазное яблоко; 4 - зрительный нерв; 5 - зрительный перекрест; 6 - зрительный путь; 7 - переднее продырявленное вещество; 8 - гипофиз; 9 - серый бугор; 10 - сосковидные тела; 11 - заднее продырявленное вещество; 12 - ножка большого мозга; 13 - мост; 14 - олива; 15 - пирамида; 16 - клочок; 17 - мозжечок; 18-21 - корешки четырех верхних спинномозговых нервов
Большая часть нижней поверхности головного мозга образована полушариями большого мозга, которые спереди разделены сквозным участком продольной щели, задней стенкой которой на нижней поверхности является конечная пластинка, lamina terminalis, - одно из образований мозгового ствола. Позади конечной пластинки по средней линии расположен ряд образований, также принадлежащих мозговому стволу: зрительный перекрест, chiasma opticum, серый бугор, tuber cinereum, с воронкой, infundibulum, и гипофизом, hypophysis, сосковидные тела, corpora mamillaria, ножки большого мозга, pedunculi cerebri, мост, pons, и продолговатый мозг, medulla oblongata, который является непосредственным продолжением спинного мозга. Кнаружи от моста и продолговатого мозга видна нижняя поверхность полушарий мозжечка. Остальные образования мозгового ствола (зрительные бугры, коленчатые тела, крыша среднего мозга и др.) скрыты в толще головного мозга и видны на его срединном разрезе.
Развитие головного мозга
Головной мозг образуется из переднего отдела нервной трубки, который уже в самых ранних стадиях развития отличается от туловищного отдела своей шириной. Неравномерный рост различных участков стенки этого отдела приводит к образованию трех расположенных друг за другом выпячиваний - первичных мозговых пузырей: переднего, prosencephalon, среднего, mesencephalon, и заднего, rhombencephalon. Далее передний и задний мозговые пузыри подразделяются на два вторичных мозговых пузыря, в результате чего возникает пять сообщающихся между собой мозговых пузырей, из которых развиваются все отделы головного мозга: конечный, telencephalon, промежуточный, diencephalon, средний, mesencephalon, задний, metencephalon, и добавочный, myelencephoton (рис. 196). Процесс образования пяти мозговых пузырей происходит одновременно с появлением изгибов головного отдела мозговой трубки в сагиттальном направлении. Вначале появляется дорсальный теменной изгиб в области mesencephalon, затем в том же направлении - затылочный изгиб между myelencephalon и спинным мозгом и, наконец, третий вентральный мостовой изгиб - в области metencephalon. Этот процесс сопровождается усиленным ростом боковых отделов головного конца нервной трубки и отставанием в росте дорсальной и вентральной стенок (покровной и донной пластинок). Утолщенные боковые отделы разделяются пограничной бороздой на основную и крылъную пластинки, из которых нейробласты основной пластинки образуют двигательные, а нейробласты крыльной - чувствительные центры. Между обеими пластинками в промежуточной зоне располагаются важные автономные центры. Пограничная борозда прослеживается на всем протяжении туловищного и головного отделов нервной трубки до промежуточного мозга. Здесь заканчивается основная пластинка, в связи с чем нервные клетки конечного мозга являются производными только крыльной пластинки. Наиболее значительная дифференцировка и изменения в форме наблюдаются при развитии производных переднего мозгового пузыря telencephalon и diencephalon.
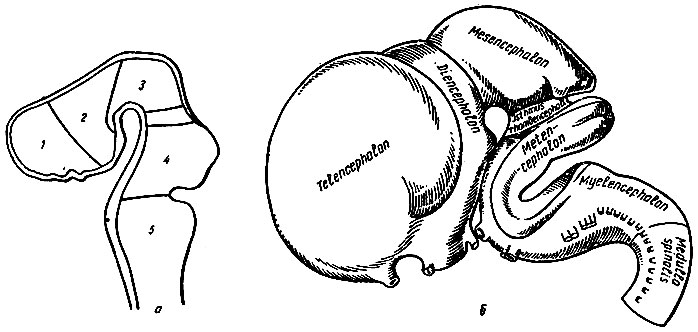
Рис. 196. Развитие головного мозга (по Р. Д. Синельникову). а - пять мозговых пузырей; 1 - первый пузырь - конечный мозг; 2 - второй пузырь - промежуточный мозг; 3 - третий пузырь - средний мозг; 4 - четвертый пузырь - собственно задний мозг; 5 - пятый пузырь - продолговатый мозг; между третьим и четвертым пузырями - перешеек; б - модель развивающегося мозга на стадии пяти пузырей
Конечный мозг, telencephalon, образуется из парного выпячивания вперед и кнаружи стенки первичного переднего мозгового пузыря, из которых развиваются правое и левое полушария мозга. Стенки этих выпячиваний быстро увеличиваются в объеме, значительно опережая в росте другие отделы мозга, и прикрывают производные других мозговых пузырей сначала с боков, а затем спереди и сверху. Неравномерный рост мозгового вещества определяет появление на поверхности образовавшихся полушарий борозд и извилин, среди которых большим постоянством отличаются те, которые появляются наиболее рано (sulcus cerebri lateralis, sulcus centralis и др.). Вместе с ростом полушарий углубляется продольная щель между ними и резко изменяется конфигурация их полостей - боковых желудочков. Межжелудочковое отверстие, сообщающее боковые желудочки с третьим, суживается. В основании полушарий развиваются скопления серого вещества - базальные или подкорковые ядра. Зачаток обонятельного мозга также относится к производным telencephalon.
Промежуточный мозг, diencephalon, формируется из задней части переднего мозгового пузыря. В процессе развития происходит резкое утолщение боковых стенок этого отдела, где образуются крупные скопления серого вещества - зрительные бугры. Кроме того, в очень ранней стадии развития, когда еще только начинается разделение переднего мозгового пузыря, боковые стенки отдают наружные выпячивания - два глазных пузыря, из которых в дальнейшем развивается сетчатая оболочка глаза и зрительные нервы. Сильное развитие зрительных бугров резко суживает полость промежуточного мозга и превращает ее в узкую продольную щель - III желудочек. Из дорсальной стенки diencephalon развивается шишковидное тело, а из выпячивания вентральной стенки образуются серый бугор, воронка и задняя доля гипофиза. Кзади от серого бугра определяются зачатки сосковидных тел.
Средний мозговой пузырь, mesencephalon, характеризуется довольно равномерным утолщением стенок, что превращает его полость в узкий канал - мозговой водопровод, соединяющий III и IV желудочки мозга. Из дорсальной стенки пузыря развивается пластинка четверохолмия, вначале нижние, а затем верхние бугры. Вентральная стенка пузыря в связи с развитием клеток и волокон других отделов головного мозга превращается в массивные волокнистые пучки - ножки мозга.
Задний мозговой пузырь, rhombencephalon, подразделяется на задний мозг, metencephalon, и продолговатый мозг, myelencephalon, а также на узкую перетяжку - перешеек ромбовидного мозга, isthmus rhombencephali, который отделяет задний мозг от среднего. Из перешейка развиваются верхние ножки мозжечка и передний мозговой парус. С вентральной стороны образуется мост, а с дорсальной - сначала червь, а затем полушария мозжечка. Развитие myelencephalon приводит к образованию продолговатого мозга.
Полости metencephalon и myelencephalon сливаются и образуют IV желудочек мозга, который сообщается с центральным каналом спинного мозга и мозговым водопроводом. Вентральные и боковые стенки желудочка в процессе развития резко утолщаются, а дорсальная стенка остается тонкой и в области продолговатого мозга состоит только из эпителиального слоя, который срастается с сосудистой оболочкой мозга, образуя tela chorioidea inferior.
Стволовая часть головного мозга
К стволовой части головного мозга относят продолговатый мозг, мозговой мост, мозжечок и образования среднего и промежуточного мозга (рис. 197).
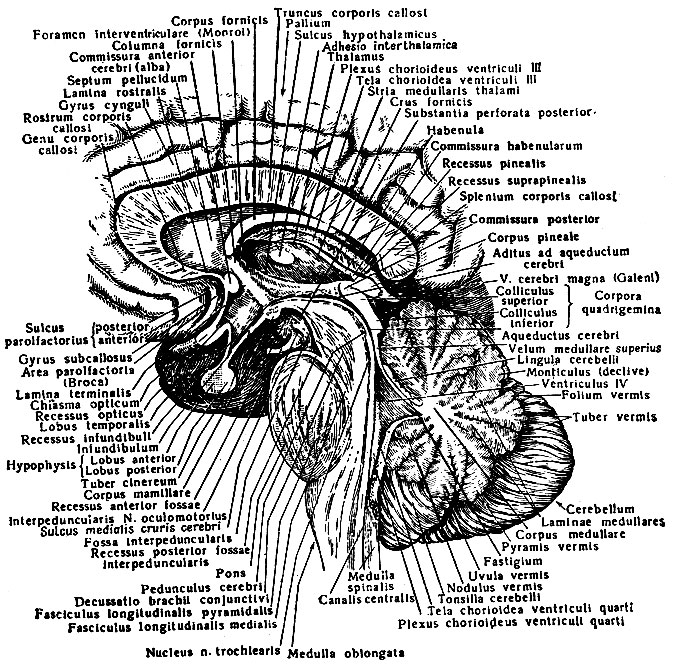
Рис. 197. Ствол головного мозга (сагиттальный разрез)
Мозговой ствол является филогетически самым древним отделом головного мозга, который анатомически и функционально связан со спинным мозгом и полушариями головного мозга, оказывающими на функции всех его отделов и центров мозгового ствола регулирующее и контролирующее влияние. В ядрах стволовой части мозга замыкаются как сравнительно простые, так и крайне сложно построенные рефлекторные дуги. Здесь находятся жизненно важные центры регуляции дыхания, сердечной деятельности, сосудистого тонуса, функций автономной нервной системы, деятельности эндокринных желез и др. Сетчатое образование ствола (см. раздел Продолговатый мозг, настоящего издания), являясь сложным рефлекторным центром, осуществляет под контролем коры полушарий большого мозга регуляцию уровня возбудимости и тонуса различных отделов центральной нервной системы.
По своему строению стволовая часть головного мозга значительно отличается от спинного мозга. Прежде всего ствол мозга утрачивает характерную для спинного мозга метамерность в строении и его нельзя разделить на подобные по своей структуре сегменты. Серое вещество ствола разделяется пучками белого вещества на различной величины и формы скопления - ядра, которые располагаются как в центре ствола, так и на периферии.
Продолговатый мозг
Продолговатый мозг, medulla oblongata, представляет собой отдел мозгового ствола длиной в среднем 25 мм, который является непосредственным продолжением спинного мозга и по своей форме напоминает усеченный конус. Утолщенный верхний отдел продолговатого мозга переходит в мозговой мост (рис. 198). Нижней границей продолговатого мозга считают место выхода первой пары корешков шейных нервов, что соответствует уровню большого затылочного отверстия, верхней границей с передней поверхности является нижний край моста, а с задней - мозговые полосы, ромбовидной ямки.
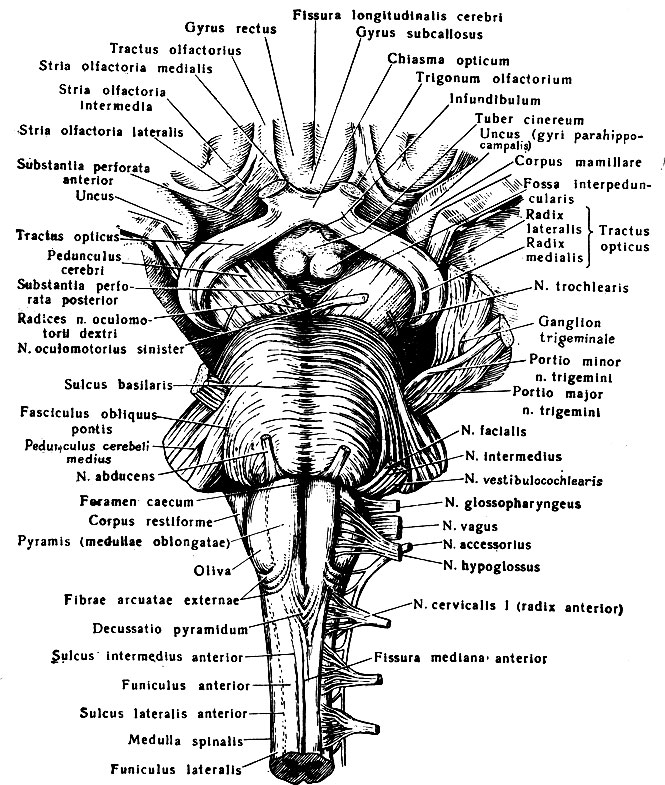
Рис. 198. Вентральная поверхность продолговатого мозга, моста и среднего мозга
В продолговатом мозге различают переднюю, вентральную, заднюю, дорсальную и боковые поверхности, вдоль которых на всем его протяжении проходят продольные борозды, продолжающиеся в соответствующие борозды спинного мозга: fissura mediana anterior, sulcus medianus posterior, sulci laterales anterior et posterior. На передней поверхности продолговатого мозга между передней срединной щелью и передней латеральной бороздой находится парное возвышение, которое называют пирамидой, pyramis. Пирамиды состоят из волокон двигательных произвольных или пирамидных путей. Большая часть волокон, образующих пирамиды, в 6-7 мм от нижней границы продолговатого мозга совершает перекрест и переходит на противоположную сторону, образуя tractus pyramidalis lateralis бокового канатика спинного мозга, а оставшиеся неперекрещенными волокна составляют tractus pyramidalis anterior его переднего канатика. Перекрест пирамидных волокон, decussatio pyramidum, прерывает переднюю срединную щель. Кнаружи от каждой пирамиды между передней и задней латеральными бороздами расположено овальной формы возвышение - олива, oliva. Из передней латеральной борозды между пирамидой и оливой выходят корешки подъязычного нерва, n. hypoglossus. Позади оливы из задней латеральной борозды выходят корешки языко-глоточного, блуждающего и добавочного черепных нервов.
Нижний отдел задней поверхности продолговатого мозга напоминает по своему рельефу спинной мозг (рис. 199). Здесь кнаружи от задней срединной борозды находятся задние канатики, разделенные промежуточной бороздой, sulcus intermedins, на медиальный - тонкий и латеральный - клиновидный пучки. В верхнем отделе задней поверхности задние канатики заканчиваются утолщениями - бугорками тонкого и клиновидного ядер, tuberculi nuclei gracilis et nuclei cuneati. Клетки этих ядер являются вторыми нейроцитами проводящих путей задних канатиков. Кнаружи от клиновидного канатика имеется незначительное возвышение, которое образует ядро спинномозгового пути тройничного нерва, nucleus tractus spinalis n. trigenimi. Это ядро отделено от поверхности продолговатого мозга тонким слоем волокон, представляющих собой нейриты клеток чувствительного узла тройничного нерва. Эти волокна образуют спинномозговой путь (болевой и тактильной чувствительности) тройничного нерва, tractus spinalis n. trigemini.
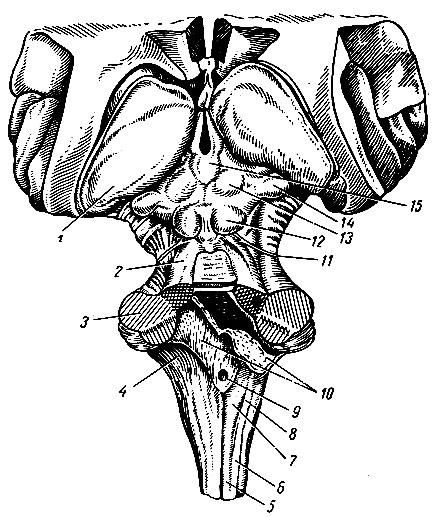
Рис. 199. Ствол головного мозга. 1 - подушка зрительного бугра; 2 - верхняя мозжечковая ножка; 3 - средняя мозжечковая ножка; 4 - нижняя мозжечковая ножка; 5 - тонкий пучок, 6 - клиновидный пучок; 7 - бугорок тонкого пучка; 8 - бугорок клиновидного пучка; 9 - срединное отверстие IV желудочка; 10 - сосудистая основа IV желудочка; 11 - блоковый нерв; 12 - нижний холмик четверохолмия; 13 - верхний холмик четверохолмия; 14 - медиальное коленчатое тело; 15 - шишковидное тело
Бугорки тонких ядер и два расходящихся в стороны довольно массивных пучка волокон, которые называются нижними мозжечковыми ножками, pedunculi cerebellares inferiores, ограничивают с боков относящуюся к продолговатому мозгу часть ромбовидной ямки - дна IV желудочка мозга. Нижние мозжечковые ножки образованы волокнами, которые соединяют мозжечок со спинным и продолговатым мозгом. Наружный отдел нижних мозжечковых ножек состоит из волокон заднего спинно-мозжечкового пути, небольшой части волокон переднего спинно-мозжечкового пути, волокон от олив продолговатого мозга преимущественно противоположной стороны - оливо-мозжечкового пути, tractus olivocerebellaris, и небольшой части волокон от ядер задних канатиков. Внутренний отдел нижних ножек мозжечка образован в основном эфферентными волокнами мозжечка к вестибулярным ядрам. Кроме того, в их состав входят афферентные волокна вестибулярных ядер к мозжечку.
Продолговатый мозг состоит из серого и белого вещества, взаимоотношения которых в нижних отделах напоминают спинной мозг, а в верхних - значительно от него отличаются. Основное отличие заключается в том, что серое вещество продолговатого мозга располагается в виде отдельных, различных по форме и величине скоплений - ядер продолговатого мозга. Ядра продолговатого мозга подразделяются на ядра черепных нервов и переключательные ядра.
В дорсальной части продолговатого мозга расположены ядра следующих черепных нервов: подъязычного, добавочного, блуждающего, языко-глоточного, преддверно-улиткового и ядро спинномозгового тракта тройничного нерва. Эти ядра преимущественно находятся в заднем отделе продолговатого мозга, который относится к ромбовидной ямке. Однако некоторые из них (например, ядро подъязычного нерва) начинаются в нижних отделах продолговатого мозга, за пределами ромбовидной ямки, а ядро добавочного нерва прослеживается только в его нижней части.
В топографии ядер черепных нервов отмечается определенная, связанная с развитием ромбовидного мозга, закономерность. Медиально возле средней линии находятся двигательные ядра, латерально - чувствительные, а автономные ядра составляют продольный ряд, которой занимает между ними промежуточное положение.
Ядро подъязычного нерва имеет значительную длину (до 10-12 мм). Оно расположено в нижнем отделе продолговатого мозга впереди от центрального канала, а в верхнем отделе - в нижнем углу ромбовидной ямки возле средней линии. Ядро образовано крупными двигательными клетками, отростки которых образуют несколько пучков волокон, проходящих в толще продолговатого мозга к передней боковой борозде. Здесь между пирамидой и оливой они выходят на поверхность в виде 10-15 корешков, из которых образуется ствол подъязычного нерва. Подъязычный нерв распределяется среди мышц языка, осуществляя их двигательную иннервацию. В ядре подъязычного нерва оканчиваются волокна от коры полушарий головного мозга, образующие пучок волокон двигательного произвольного пирамидного пути. Кроме того, в нем оканчиваются волокна от чувствительного ядра тройничного нерва и ядра одиночного пучка.
Ядра добавочного нерва расположены в продолговатом п спинном мозге. Спинномозговое ядро лежит в передних рогах спинного мозга и прослеживается с V шейного сегмента. Отростки клеток этого ядра образуют корешки n. accessorius. Краниальная часть нерва образуется отростками клеток двух ядер, расположенных в нижнем отделе продолговатого мозга. Из них переднее ядро находится в сетчатом образовании продолговатого мозга, а заднее - позади центрального канала. Волокна краниальной части нерва в виде 4-5 корешков выходят из задней борозды между оливой и веревчатым телом. Добавочный нерв иннервирует грудино-ключично-сосцевидные и трапециевидные мышцы.
Из трех ядер блуждающего нерва вегетативное дорсальное ядро, nucleus dorsalis, расположено кнаружи и кзади от ядра подъязычного нерва. Оно представляет собой продольный столб клеток длиною до 20 мм и прослеживается от нижней границы продолговатого мозга до уровня верхнего полюса оливы. Двигательное соматическое ядро блуждающего нерва носит название двоякого, nucleus ambiguus, так как его клетки дают начало волокнам как блуждающего, так и языко-глоточного нервов. Это ядро имеет длину около 10 мм и располагается в нижнем отделе сетчатого образования продолговатого мозга.
Двигательные волокна от дорсального и двоякого ядер в составе 12-15 корешков выходят из продолговатого мозга в области задней боковой борозды, выше корешков добавочного нерва. Эти волокна осуществляют двигательную иннервацию как поперечнополосатой, так и гладкой мускулатуры пищеварительного тракта, органов дыхания и сердца. В составе указанных корешков блуждающего нерва к его чувствительному ядру - ядру одиночного пути, nucleus tractus solitarii, подходят центральные отростки клеток его афферентных узлов (gangl. superius et inferius), расположенных в области яремного отверстия, а также центральные отростки чувствительных клеток ствола блуждающего нерва. В толще продолговатого мозга в сетчатом образовании центральные отростки делятся на восходящие и нисходящие ветви, из которых образуется одиночный путь, tractus solitarius. Одиночный путь находится кнаружи от дорсального ядра блуждающего нерва. Он окружен серым веществом - ядром одиночного пути, где заканчиваются первые нейроциты чувствительных путей блуждающего нерва, осуществляющих чувствительную иннервацию тех же органов, в двигательной иннервации которых он участвует.
Языко-глоточный нерв, как и блуждающий, имеет три ядра, расположенных в продолговатом мозге. Из них двигательное соматическое двоякое ядро, nucleus ambiguus, является общим с блуждающим нервом; Волокна языко-глоточного нерва являются отростками клеток верхнего отдела этого ядра. Двигательное вегетативное ядро, которое называют нижним слюноотделительным ядром, nucleus salivatorius inferior, образовано клетками, лежащими в ретикулярной формации между двояким ядром и оливой. Это ядро иннервирует околоушную слюнную железу. Волокна двигательного дорсального ядра языко-глоточного нерва выходят из продолговатого мозга в составе 5 или 6 корешков из задней боковой борозды рядом с корешками блуждающего нерва. В составе этих корешков в продолговатый мозг вступают центральные отростки клеток афферентных узлов языко-глоточного нерва, gangl. superius et inferius, которые расположены в области яремного отверстия. Эти волокна входят в состав одиночного пути и заканчиваются в окружающем его сером веществе - ядро одиночного пучка. Они являются первыми нейроцитами, осуществляющими чувствительную иннервацию миндалин, небных дужек, слизистой оболочки задней трети языка, глотки, а также передают специальную вкусовую чувствительность.
Комплекс ядер преддверно-улиткового нерва расположен в области латеральных углов ромбовидной ямки. Имеются 4 преддверных ядра: латеральное, nucleus lateralis, медиальное, nucleus medialis, верхнее, nucleus superior, и нижнее, nucleus inferior. Эти ядра отличаются друг от друга по своему строению и связям. В них заканчиваются центральные отростки рецепторных нейроцитов вестибулярного узла, gangl. vestibulare, который находится в meatus acusticus internus и проводит к ним импульсы от рецепторного аппарата преддверия и полукружных каналов. Несколько выше преддверных ядер находятся дорсальное и вентральное слуховые ядра, nuclei cochleares ventralis et dorsalis, имеющие длину до 3 мм. В них заканчиваются центральные отростки рецепторных нейронов расположенного в улитке спирального узла, gangl. spirale, которые проводят импульсы от слухового аппарата. Отростки клеток вентрального ядра образуют трапециевидное тело моста, а отростки дорсального - striae medullares ромбовидной ямки.
Из моста через продолговатый мозг до средних шейных сегментов проходит нисходящий пучок волокон - спинномозговой путь тройничного нерва, который образован центральными отростками клеток его чувствительного узла. Этот путь на всем протяжении прилегает к ядру спинномозгового пути тройничного нерва, где волокна этого пути заканчиваются. Если в области моста и на уровне олив продолговатого мозга это ядро расположено в толще мозгового вещества, то возле нижнего полюса олив оно занимает поверхностное положение, образуя серый бугорок и канатик.
Переключательные ядра продолговатого мозга представляют собой массивные скопления клеток. Наиболее крупными из этих ядер являются оливы, составляющие три клеточных скопления: оливное ядро, nucleus olivaris, медиальное добавочное ядро, nucleus olivaris accessorius medialis, дорсальное добавочное оливное ядро, nucleus olivaris accessorius dorsalis. Оливное ядро представляет собой элипсовидной формы образование длиной до 10 мм, которое состоит из серого и белого вещества. Серое вещество в виде тонкой складчатой пластинки ограничивает центрально расположенное белое вещество, оставляя свободным медиальный участок оливы. Это место носит название ворот оливного ядра, hilus nuclei olivaris. Кнутри от оливного ядра находится медиальное добавочное оливное ядро, а кзади расположено дорсальное добавочное оливное ядро. Весь этот комплекс ядер является промежуточным центром равновесия.
Ядра задних канатиков - тонкое и клиновидное - представляют собой скопления клеток длиной до 13 мм. В одних ядрах заканчиваются центральные отростки первого чувствительного нейроцита, которые образуют тонкий и клиновидный пучки спинного мозга и проводят импульсы глубокой и отчасти тактильной чувствительности. Отростки клеток этих ядер - вторые нейроциты этого пути - выше места перекреста пирамид в виде внутренних дугообразных волокон, fibrae arcuatae internae, обходят спереди центральный канал, образуя верхний чувствительный перекрест петли, decussatio lemniscorum. После перекреста эти волокна образуют внутреннюю, или медиальную, петлю, lemniscus medialis, которая проходит, увеличиваясь в объеме, через все отделы ствола к зрительному бугру промеячуточного мозга. Увеличение объема медиальной петли связано с присоединением к ней пучков нервных волокон болевой и температурной чувствительности (спинно-бугорный путь), а также от ядра спинно-мозгового пути тройничного нерва. Кроме ядер черепных нервов и переключательных ядер, к серому веществу продолговатого мозга относится центрально расположенное сетчатое образование, formatio reticularis. Участки концентрации нервных клеток называют ядрами сетчатого образования. В отличие от черепных и переключательных ядер они представляют собой пространственно не замкнутые клеточные группы без четких границ между отдельными ядрами. Наиболее крупные нервные клетки, сходные по строению с двигательными, образуют медиально расположенную группу ядер сетчатого образования; латеральнее от них находится группа ядер, клетки которых напоминают по своей структуре чувствительные ядра ствола. В ядрах сетчатого образования заканчиваются коллатерали как чувствительных, так и двигательных путей, проходящих через мозговой ствол, вследствие чего наблюдается постоянный уровень возбуждения составляющих его нейронов, который регулируется корой полушарий головного мозга. Сетчатое образование стволовой части мозга имеет большое значение в регуляции возбудимости и тонуса различных отделов центральной нервной системы, обеспечивает готовность различных центров к деятельности, усиливает или тормозит рефлекторную деятельность спинного мозга (tractus reticulospinalis), поддерживает бодрствующее состояние коры полушарий головного мозга.
Белое вещество продолговатого мозга состоит из пучков собственных и проходящих через него нервных волокон. Собственные, или эндогенные волокна подразделяются на короткие и длинные. Короткие волокна соединяют между собой ядра, расположенные в пределах продолговатого мозга. Длинные эндогенные волокна являются отростками клеток ядер продолговатого мозга, которые заканчиваются в других отделах нервной системы. К этой группе относятся волокна медиальной петли, оливо-спинномозговые, оливо-мозжечковые, сетчато-спинномозговые пути.
Проходящие через продолговатый мозг тракты, которые не возникают и не оканчиваются в его ядрах, носят название экзогенных. Среди них различают эфферентные (корково-спинномозговой, красноядерно-спинномозговой, покрышечно-спинномозговой) и афферентные (передний и боковой спинно-бугорные пути, передний и задний спинно-мозжечковые пути, спинно-покрышечный путь). Топографоанатомические взаимоотношения проводящих путей и ядер продолговатого мозга видны на рис. 200.
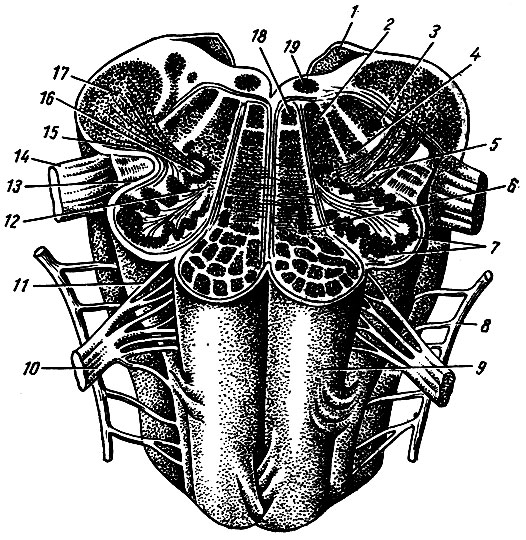
Рис. 200. Продолговатый мозг (горизонтальный разрез на уровне оливы). 1 - нижний мозговой парус; 2 - сетчатое образование; 3 - ядро спинномозгового пути тройничного нерва; 4 - двоякое ядро; 5 - оливо-спинномозговой путь; 6 - медиальное добавочное оливное ядро; 7, 16 - оливное ядро; 8 - добавочный нерв; 9 - пирамида; 10 - подъязычный нерв; 11 - олива; 12 - ворота оливного ядра; 13 - покрышечно-спинномозговой путь; 14 - блуждающий нерв; 15 - красно-ядерно - спинномозговой путь; 17 - нижняя ножка мозжечка; 18 - медиальный продольный пучок; 19 - ядро подъязычного нерва
Мост
Мост, pons, является частью мозгового ствола длиной 25 мм, которая расположена между продолговатым и средним мозгом. Его вентральная поверхность образована выпуклостью белого цвета, снаружи состоящей из поперечно расположенных волокон. Дорсальная поверхность моста составляет верхнюю часть дна IV желудочка - ромбовидной ямки, образуя ее верхний треугольник. Эта часть ромбовидной ямки ограничена верхними мозжечковыми ножками. Поперечные волокна передней поверхности образуют средние мозжечковые ножки, которые погружаются в толщу полушарий мозжечка. Границей между мостом и средней мозжечковой ножкой служит linea trigeminofacialis, проходящая между корешками тройничного и лицевого нервов. Посередине вентральной поверхности моста расположена основная борозда, sulcus basilaris. В боковом отделе вентральной поверхности моста, ближе к его переднему краю, находятся корешки тройничного нерва. В области мосто-мозжечкового угла (образован продолговатым мозгом, мостом и мозжечком) расположены корешки лицевого, промежуточного и преддверно-улиткового нервов, а ближе к средней линии, между задним краем моста и пирамидами, - корешки отводящих нервов.
На поперечных срезах моста различают крупную вентральную часть, pars ventralis pontis, и меньшую - дорсальную, pars dorsalis pontis, границей между которыми является пучок поперечных волокон - трапециевидное тело, corpus trapezoideum. Вентральная и дорсальная части моста образованы серым и белым веществом. Однако вентральная часть состоит преимущественно из белого вещества.
Серое вещество вентральной части слагается из многочисленных собственных ядер моста, nuclei pontis. В этих ядрах заканчиваются корково-мостовые пути, tractus corticopontinus, и коллатерали от пирамидных путей. Волокна клеток ядер моста образуют поперечные волокна моста, fibrae pontis transversae, которые, в основном, переходя на противоположную сторону, составляют средние ножки мозжечка и заканчиваются в клетках коры его полушарий. Поперечные волокна моста в вентральной части его нижнего отдела образуют поверхностный и глубокий слои, между которыми проходят пучки пирамидного пути. В верхних отделах моста глубокий слой его поперечных волокон увеличивается в объеме и появляется третий слой, разделяющий пирамидные пути на более мелкие пучки.
Серое вещество дорсальной части моста состоит из центрально расположенного сетчатого образования, ядер черепных нервов и переключательных ядер. Сетчатое образование моста является непосредственным продолжением одноименного образования продолговатого мозга.
В мосту расположены ядра следующих черепных нервов: отводящего, лицевого, тройничного, преддверно-улиткового.
Ядро отводящего нерва, nucleus n. abducentis, образовано крупными двигательными клетками. Оно расположено возле средней линии дна IV желудочка и имеет длину около 3 мм. Отростки клеток ядра выходят через толщу моста между его задним краем и пирамидой в виде корешка отводящего нерва.
Ядро лицевого нерва, nucleus n. facialis, длиной от 2 до 5,6 мм, образовано двигательными клетками. Оно расположено в сетчатом образовании дорсальной части моста. Отростки клеток этого ядра образуют внутримозговой отдел корешка лицевого нерва, который в толще моста имеет сложный ход. Корешок из сетчатого образования проходит по дну IV желудочка, образует колено, genu n. facialis, окружающее ядро отводящего нерва и далее идет вперед через толщу моста в область мостомозжечкового угла. Волокна лицевого нерва распределяются в мимической мускулатуре и осуществляют ее двигательную иннервацию. Несколько кзади от ядра лицевого нерва в сетчатом образовании моста находится верхнее слюноотделительное ядро, nucleus salivatorius superior, которое является секреторным вегетативным центром иннервации подъязычной и подчелюстной слюнных желез и слезной железы. Чувствительным ядром этого нерва является ядро одиночного пути, nucleus tractus solitarii, где оканчиваются центральные отростки узла промежуточного нерва - узла колена, gangl. geniculi, расположенного в канале лицевого нерва пирамиды височной кости.
В среднем и верхнем отделах дорсальной части моста расположены чувствительное и двигательное ядро тройничного нерва. Двигательное ядро тройничного нерва, nucleus motorius n. trigemini, образовано крупными двигательными клетками и имеет длину около 4 мм. Отростки клеток двигательного ядра образуют двигательный корешок тройничного нерва и распределяются в жевательной мускулатуре, осуществляя ее двигательную иннервацию. В составе двигательного корешка тройничного нерва проходят также волокна от ядра тройничного нерва, расположенного в среднем мозге, латеральнее водопровода большого мозга. Верхнее чувствительное ядро тройничного нерва, nucleus sensorius n. trigemini superior, лежит кнаружи от двигательного ядра. Оно меньше, чем ядро спинномозгового тракта тройничного нерва. В клетках этого ядра оканчиваются центральные отростки чувствительных клеток тройничного узла. Эти отростки образуют чувствительный корешок тройничного нерва, который через толщу базальной части моста подходит к верхнему чувствительному ядру. Здесь часть волокон корешка заканчивается, а остальные волокна проходят к ядрам спинномозгового и среднемозгового путей Тройничного нерва. Отростки клеток верхнего чувствительного ядра спинномозгового тракта (вторые нейроциты) переходят на противоположную сторону и в составе медиальной петли вступают в зрительный бугор.
К переключательным ядрам дорсальной части моста относятся верхние оливы, ядра трапециевидного тела и боковой петли. Во всех этих ядрах происходит переключение импульсов слухового пути.
Верхняя олива, oliva superior, расположена в латеральных отделах трапециевидного тела, которое образуют преимущественно отростки клеток вентрального ядра n. vestibulocochlearis. Между волокнами трапециевидного тела находятся скопления серого вещества - вентральное и дорсальное ядра трапециевидного тела, nuclei ventralis et dorsalis corporis trapezoidei. Большая часть волокон, которые возникают в вентральном ядре n. vestibulocochlearis, переходят на противоположную сторону и заканчиваются в верхней оливе и ядрах трапециевидного тела. Меньшая часть этих волокон заканчивается в соответствующих ядрах своей стороны. Отростки клеток верхней оливы образуют так называемую боковую петлю, lemniscus lateralis, среди волокон которой находится ядро боковой петли, nucleus lemnisci lateralis. Боковая петля представляет собой значительный по толщине пучок волокон; в ее состав входят отростки клеток дорсального ядра n. vestibulocochlearis, а также отростки клеток ядра трапециевидного тела и ядра боковой петли. Боковая петля заканчивается в первичных слуховых центрах - нижних буграх четверохолмия и медиальном коленчатом теле.
Белое вещество моста состоит из эндогенных и экзогенных волокон. Короткие эндогенные волокна соединяют отдельные ядра моста и не выходят за его пределы. Так, установлены нервные связи между ядрами лицевого и тройничного нервов, посредством которых замыкаются рефлекторные дуги при различных раздражениях кожи лица с ответной реакцией мимических мышц. Длинные эндогенные волокна возникают в ядрах моста и заканчиваются в других отделах центральной нервной системы.
К этой группе относятся волокна, идущие от собственных ядер моста к мозжечку - поперечные волокна моста, fibrae pontis transversae, волокна латеральной петли, пучки волокон от некоторых черепных нервов. Поперечные волокна моста образуют среднюю мозжечковую ножку, посредством которой осуществляется влияние коры полушарий головного мозга на деятельность мозжечка.
Экзогенные волокна моста представлены эфферентными и афферентными путями, которые участвуют в образовании белого вещества продолговатого мозга. Кроме них, в состав белого вещества моста входят корково-ядерные волокна, fibrae corticonucleares, и корково-мостовой путь, tractus certicopontinus. Пирамидный путь поперечными волокнами моста разделен на несколько отдельных пучков.
Мозжечок
Мозжечок, cerebellum, тесно связан с продолговатым мозгом, мостом и средним мозгом и, располагаясь дорсальнее от этих образований, заполняет большую часть задней черепной ямки (см. рис. 197). Вес мозжечка около 150 г. Его наибольший поперечный размер составляет 10-12 см, продольный в области червя - до 4 см, а в области полушарий - до 6 см. В мозжечке различают центральную узкую часть - червь, vermis, и две крупные выпуклые боковые части - полушария, hemispherii cerebelli (рис. 201). От полушарий червь отделяют правая и левая продольные щели, которые лучше выражены на нижней поверхности мозжечка. На верхней поверхности щели неглубокие и прерывистые, в связи с чем граница между червем и полушариями здесь слабо выражена. Верхняя поверхность мозжечка более ровная, чем нижняя, так как полушария выступают книзу больше, чем червь. Поэтому между их внутренними поверхностями и червем образуется широкое и довольно глубокое пространство - долинка мозжечка, vallecula cerebelli. Посередине долинки имеется глубокая, направленная кверху и кзади выемка, соответствующая шатру IV желудочка. В области переднего, наиболее широкого отдела долинки расположен дорсальный отдел среднего мозга, а далее книзу - мост и продолговатый мозг.
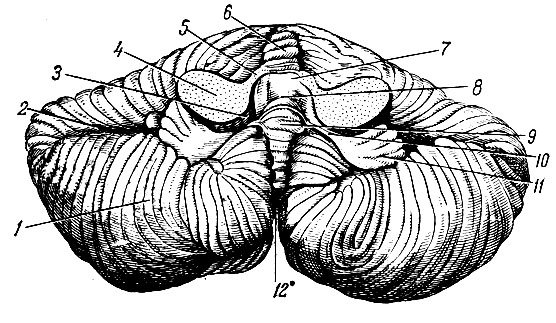
Рис. 201. Мозжечок (вид снизу). 1 - полушарие мозжечка; 2 - горизонтальная щель мозжечка; 3 - нижний мозговой парус; 4 - средняя ножка мозжечка; 5 - верхняя ножка мозжечка; 6 - червь; 7 - верхний мозговой парус; 8 - IV желудочек; 9 - узелок; 10 - ножка клочка; 11 - клочок; 12 - задняя вырезка мозжечка
Различной глубины многочисленные щели, fissurae cerebelli, разделяют поверхность полушарий и червя мозжечка на доли, lobi cerebelli, дольки, lobuli cerebelli, листки, folia cerebelli. Наличие большого количества щелей значительно увеличивает поверхность мозжечка. Наиболее глубокой щелью является горизонтальная, fissura horizontalis, которая соответствует границе между верхней и нижней поверхностями. Щели мозжечка, разделяющие его на доли, не прерываясь проходят через червь на полушария. Поэтому каждой доле червя соответствуют две доли полушарий. Верхняя поверхность червя состоит из язычка, lingula, центральной дольки, lobulus centralis, вершины, culmen, ската, declive, и листа червя, folium vermis (см. рис. 197). Этим долям червя соответствуют следующие доли полушария: крыло центральной дольки, ala lobuli centralis, четырехугольная долька, lobulus quadrangularis, простая долька, lobulus simplex, и верхняя полу лунная долька, lobulus semilunaris superior.
Нижняя поверхность червя слагается из бугра червя, tuber vermis, пирамиды, pyramis, язычка, uvula, и узелка, nodulus. Дольки полушария, соответствующие этим долям, следующие: нижняя полулунная долька, lobulus semilunaris inferior, двубрюшная долька, lobulus biventer, миндалина, tonsilla, и клочок, flocculus. Соединяющие клочок и узелок полулунные пластинки белого вещества носят название ножки клочка, pedunculi flocculi. К ножкам клочка прикрепляется задний мозговой парус.
В отличие от других отделов мозгового ствола серое вещество мозжечка преимущественно сосредоточено на его поверхности в виде двухслойной коры, cortex cerebelli. Под корой находится белое вещество, масса которого носит название мозгового тела, corpus medullare. Внутри мозгового тела расположены парные подкорковые ядра серого вещества.
Кора покрывает как свободную поверхность извилин мозжечка, так и поверхность, расположенную в глубине борозд. Она состоит из двух слоев: светлого наружного - молекулярного, stratum molecularе, и темного внутреннего - зернистого, stratum granulosum. В глубине молекулярного слоя находится один ряд крупных ганглиозных клеток, которые являются эфферентными клетками коры, в то время как клетки молекулярного и зернистого слоев - вставочные и ассоциативные нейроны.
Подкорковые ядра мозжечка представляют собой-различной формы и величины скопления серого вещества. Из них наиболее крупное - зубчатое ядро, nucleus dentatus, расположенное во внутренне-нижнем отделе мозгового тела. Зубчатое ядро представляет собой складчатую пластинку, которая незамкнута с внутренней стороны и окаймляет центрально расположенное белое вещество. Участок зубчатого ядра, где пластинка серого вещества отсутствует, носит название ворот, hilus nuclet dentati. К зубчатому ядру подходят нейриты ганглиозных клеток полушарий мозжечка и сравнительно небольшое количество аксонов ганглиозных клеток червя. По этим нейритам нервные импульсы из коры передаются клеткам зубчатого ядра. Нейриты клеток зубчатого ядра образуют его центрально расположенное белое вещество, выходят через ворота и далее составляют основную часть волокон верхней ножки мозжечка. Пробковидное ядро, nucleus embolijormis, расположено в белом веществе полушария, медиальнее верхнего края зубчатого ядра. Самое медиальное положение среди подкорковых ядер мозжечка над шатром IV желудочка занимает ядро шатра, nucleus fastigii. Между пробковидным ядром и ядром шатра расположено шаровидное ядро, nucleus globosus.
Белое вещество мозжечка состоит из вне- и внутримозжечковых волокон, которые составляют мозговое тело мозжечка и центральную часть его листовидных извилин.
Группу внутримозжечковых волокон образуют отростки клеток коры мозжечка. Среди них различают: 1) ассоциационные волокна, которые соединяют различные участки коры мозжечка; 2) комиссуральные волокна, соединяющие участки коры противоположных полушарий; 3) короткие проекционные волокна - отростки ганглиозных клеток к подкорковым ядрам мозжечка. Короткие проекционные волокна являются первыми нейронами эфферентных путей мозжечка.
К внемозжечковым волокнам относятся длинные проекционные эфферентные и афферентные волокна, посредством которых мозжечок соединен с другими отделами головного мозга. Эти волокна образуют три пары мозжечковых ножек, из которых нижние и средние образованы преимущественно афферентными волокнами, а верхние - эфферентными волокнами, возникающими в подкорковых ядрах мозжечка.
В составе нижних мозжечковых ножек в мозжечок вступают: задний спинномозговой путь, от ядер задних канатиков - наружные дугообразные волокна, fibrae arcuatae externae, волокна от преддверных ядер к ядру шатра, волокна от оливы, tractus olivocerebellaris. Кроме того, в состав нижних мозжечковых ножек входит путь от ядра шатра к боковому преддверному ядру. Средние мозжечковые ножки образуют поперечные волокна моста, которые соединяют ядра моста с корой мозжечка.
В составе верхних мозжечковых ножек в мозжечок вступает передний спинно-мозжечковый путь, а в основном это образование составляют отростки клеток зубчатого ядра, которые после перекреста заканчиваются в красном ядре среднего мозга и зрительном бугре. Мозжечок участвует в координации всех сложных двигательных актов организма.
Перешеек ромбовидного мозга
К перешейку ромбовидного мозга, isthmus rhombencephali, относят верхние мозжечковые ножки, верхний мозговой парус и треугольник петли.
Верхний мозговой парус, velum medullare superius, представляет собой тонкую треугольной формы пластинку белого вещества, которая находится между обеими верхними мозжечковыми ножками и образует верхнюю часть крыши IV желудочка. От верхнего участка верхнего мозгового паруса к борозде, расположенной между буграми четверохолмия, тянется непарный тяж белого вещества - уздечка, frenulum. По бокам уздечки находятся корешки блоковых нервов.
Треугольник петли, trigonum lemnisci, является местом, где волокна латеральной петли расположены наиболее близко к поверхности. Этот участок ограничивают ручка нижнего холмика, верхняя мозжечковая и латеральная борозды среднего мозга, sulcus lateralis mesencephali, которая служит границей между перешейком и средним мозгом.
Четвертый желудочек
Четвертый желудочек, ventriculus quartus, является полостью добавочного и заднего мозга, в связи с чем в образовании его стенок участвуют продолговатый мозг, мост, мозжечок и перешеек.
По форме IV желудочек сравнивают с палаткой, в которой различают дно, боковые стенки и крышу. Дно этого желудочка составляет ромбовидная ямка, fossa rhomboidea, которая представляет собой ограниченный верхними и нижними мозжечковыми ножками ромбовидной формы участок дорсальной поверхности моста и продолговатого мозга (рис. 202). Верхний угол ромбовидной ямки переходит в водопровод большого мозга - полость среднего мозга. Нижний угол прикрыт небольшой пластинкой - задвижкой, obex, и продолжается в центральный канал спинного мозга. Боковые углы ромбовидной ямки переходят в боковые карманы IV желудочка, recessus lateralis. Срединная борозда, sulcus medianus, делит ромбовидную ямку на две симметричные половины. По бокам от этой борозды находится парный продольный валик - медиальное возвышение, eminentia medialis, наружной границей которого является пограничная борозда, sulcus limitans. Сверху и снизу пограничная борозда заканчивается углублениями - верхней и нижней ямками, foveae superior et inferior. Верхняя ямка соответствует месту расположения двигательного ядра тройничного нерва. Медиальное возвышение книзу принимает форму треугольника, который называют треугольником подъязычного нерва, trigonum n. hypoglossi. Здесь находится ядро подъязычного нерва. Латеральнее этого треугольника расположено серого цвета треугольной формы возвышение - треугольник блуждающего нерва, trigonum n. vagi, где расположено дорсальное ядро блуждающего нерва. В области моста eminentia medialis имеет вид кругловатого возвышения и называется лицевым бугром, colliculus facialis. Здесь находится окружающая ядро отводящего нерва внутримозговая часть корешка лицевого нерва. Несколько выше верхней ямки видно аспидно-серого цвета вдавление - голубоватое место, locus ceruleus.
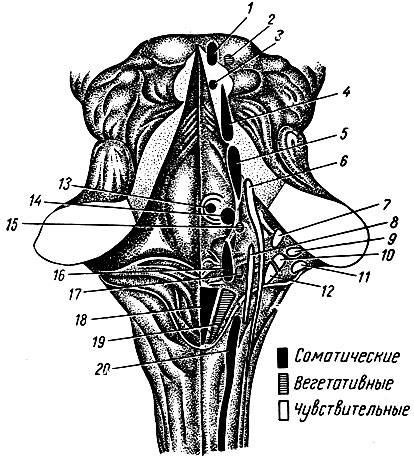
Рис. 202. Ромбовидная ямка и ядра черепных нервов. 1 - ядро глазодвигательного нерва; 2 - добавочное ядро глазодвигательного нерва; 3 - ядро блокового нерва; 4 - ядро среднемозгового пути тройничного нерва; 5 - двигательное ядро трой-пичного нерва; 6 - верхнее чувствительное ядро тройничного нерва; 7 - верхнее преддверное ядро; 8 - ядро одиночного пути; 9 - вентральное улитковое ядро; 10 - боковое преддверное ядро; 11 - дорсальное улитковое ядро; 12 - медиальное преддверное ядро; 13 - ядро отводящего нерва; 14 - ядро лицевого нерва; 15 - верхнее слюноотделительное ядро; 16 - двоякое ядро; 17 - нижнее слюноотделительное ядро; 18 - ядро подъязычного нерва; 19 - дорсальное ядро блуждающего нерва; 20 - ядро добавочного нерва
Наружные отделы ромбовидной ямки в области боковых углов называются преддверным полем, area vestibularis. Здесь расположены ядра преддверно-улиткового нерва.
Боковые стенки IV желудочка образованы тремя парами мозжечковых ножек. Крыша IV желудочка, tegmen ventriculi quarti, образуется верхним мозговым парусом, веществом мозжечка и нижним мозговым парусом (см. рис. 197).
Нижний мозговой парус, velum medullare inferius, формируется тонкой эпителиальной пластинкой - остатком задней стенки первичного мозгового пузыря. Эта пластинка дополняется сосудистой основой IV желудочка, tela chorioidea ventriculi quarti. Нижний мозговой парус становится хорошо виден после удаления миндалины мозжечка.
По средней линии нижнего мозгового паруса имеется срединное отверстие IV желудочка, apertura mediana ventriculi quarti. Боковыми отверстиями, aperturae laterales ventriculi quarti, заканчиваются латеральные карманы IV желудочка. Указанные отверстия соединяют между собой полость IV желудочка и субарахноидальное пространство головного мозга. Кроме tela chorioidea ventriculi quarti, в полости IV желудочка имеется сосудистое сплетение, plexus chorioideus, которое в виде двух тяжей, переплетенных между собой и сильно извитых сосудов, располагается вдоль средней линии, проходит в боковые карманы и выходит наружу через боковые отверстия IV желудочка в области мосто-мозжечкового угла позади клочка.
Средний мозг
Средним мозгом называют отдел мозгового ствола, который расположен между мостом и промежуточным мозгом. В его состав входят ножки большого мозга и крыша среднего мозга.
Ножки большого мозга, pedunculi cerebri, представляют собой два расходящихся под острым углом массивных валика, которые образованы продольно ориентированными нервными волокнами. Между ножками мозга находится межножковая ямка, fossa inter peduncularis, которая закрыта тонкой пластинкой, пронизанной множеством отверстий для кровеносных сосудов, - задним продырявленным веществом, substantia perforata posterior. На медиальной поверхности ножки мозга имеется медиальная борозда, sulcus medialis cruris cerebri, в которой расположен корешок глазодвигательного нерва. Латеральную поверхность ножки мозга огибает корешок блокового нерва. В области вступления ножек мозга в толщу полушарий на их поверхности расположены зрительные тракты.
Крыша среднего мозга, tectum mesencephali, составляет его дорсальный отдел, который скрыт под полушариями большого мозга. Пластинка крыши, lamina tecti, продольной и поперечной бороздами делится на два верхних и два нижних холмика, colliculi superiores et inferiores. В переднем отделе продольной борозды находится шишковидная железа, а от ее заднего конца возникают волокна, образующие уздечку верхнего мозгового паруса. Наружная поверхность каждого холмика переходит в пучок волокон, который называют ручкой холмика, brachium colliculi. Ручка верхнего холмика проходит в область промежуточного мозга к латеральному коленчатому телу, corpus geniculatum laterale, а часть ее волокон - в зрительный тракт. Ручка нижнего холмика вступает в медиальное коленчатое тело, corpus geniculatum mediate.
Полостью среднего мозга является узкий канал длиной около 2 см, который называют водопроводом большого мозга, aqueductus cerebri. Этот канал выстлан эпендимой и соединяет IV и III желудочки мозга.
На поперечных срезах среднего мозга выделяют три отдела: крышу среднего мозга, tectum mesencephali, дорсальную часть pedunculus cerebri - покрышку, tegmentum, и вентральную часть pedunculus cerebri - ногу большого мозга, crus cerebri. Границей между покрышкой и ногой большого мозга является черное вещество ножек большого мозга, substantia nigra. Нога большого мозга образована белым веществом, которое состоит из продольных эфферентных путей (пирамидных и корково-мостовых).
Покрышку и крышу наряду с белым веществом образуют ядра серого вещества, причем белое вещество покрышки состоит как из эфферентных (красноядерно-спинномозговой путь), так и из афферентных (медиальная и латеральная петли) проводящих путей.
Серое вещество крыши среднего мозга составляет ядра верхних и нижних холмиков.
Ядра нижних холмиков, nuclei colliculi inferiores, являются первичными слуховыми рефлекторными центрами. В них заканчивается часть волокон латеральной петли. Отростки клеток этих ядер образуют ручки нижних холмиков, которые вступают в медиальное коленчатое тело, а часть волокон входит в состав tractus tectospinalis et tractus tectobulbaris, заканчивающихся в двигательных ядрах мозгового ствола и спинного мозга. При участии ядер нижних холмиков осуществляются ориентировочные звуковые рефлексы - поворот головы и туловища по направлению к новому звуку.
Ядра верхних холмиков - серые слои, stratum griseum colliculi superioris, являются первичными зрительными рефлекторными центрами. В ядрах верхних холмиков заканчивается часть волокон зрительного тракта, а также волокна от спинного мозга, идущие в составе tractus spinotectalis, и ответвления латеральной и медиальной петли. Клетки этих ядер образуют основную массу волокон tractus tectospinalis et tractus tectobulbaris, которые, как уже известно, заканчиваются в двигательных ядрах мозгового ствола и спинного мозга. Они осуществляют зрительные ориентировочные рефлексы - движение туловища и глаз на световое раздражение.
Серое вещество покрышки среднего мозга представлено несколькими ядрами и ретикулярной формацией, которая является продолжением в краниальном направлении сетчатого образования моста. В окружающем водопровод большого мозга центральном сером веществе выделяют значительные по длине (5-6 мм) ядра глазодвигательных нервов. Эти парные ядра расположены вентрально от водопровода большого мозга на уровне верхних холмиков крыши среднего мозга. Верхний конец этих ядер заходит в область промежуточного мозга. Ядро глазодвигательного нерва состоит из двух отделов: двигательного соматического и автономного парасимпатического. Это ядро посредством волокон медиального продольного пучка, fasciculus longitudinalis medialis, связано с ядрами блокового и отводящего нервов, а также с системой преддверных ядер. Часть волокон, возникающих в ядре глазодвигательного нерва, оканчивается в ядре лицевого. Благодаря нервным связям между ядрами нервов глазодвигательного аппарата осуществляется регуляция согласованной деятельности мышц глазного яблока, а их связи с преддверными ядрами объясняют реакцию глазных яблок на раздражения вестибулярного аппарата (нистагм). Через медиальный продольный пучок правые и левые ядра глазодвигательного и отводящего нервов связаны между собой, благодаря чему осуществляется сочетание движений обоих глазных яблок. В ядре глазодвигательного нерва оканчиваются волокна от верхних холмиков крыши среднего мозга, посредством которых осуществляется связь этого ядра со зрительным анализатором.
На задней поверхности медиального продольного пучка на уровне верхней части нижних холмиков крыши среднего мозга расположены парные ядра блокового нерва. Самым длинным из ядер черепных нервов среднего мозга является ядро среднемозгового пути тройничного нерва. Группа клеток, образующих это ядро, расположена непосредственно около водопровода большого мозга с латеральной стороны на протяжении 22 мм. Аксоны клеток этого ядра образуют tractus mesencephalicus n. trigemini, который по наружной стенке IV желудочка доходит до среднего отдела моста, где присоединяется к двигательному корешку тройничного нерва.
Самым крупным ядром покрышки среднего мозга является парное красное ядро, nucleus ruber (рис. 203). В этих ядрах заканчиваются большая часть волокон верхних мозжечковых ножек, волокна от бледного шара, globus pallidus, волокна от зрительного бугра, а также ответвления медиальной и латеральной петель и ядер крыши среднего мозга. От клеток красного ядра начинается красноядерно-спинномозговой путь, tractus rubrоspinalis, который после перекреста в среднем мозге заканчивается в двигательных ядрах спинного мозга. Совместно с сетчатым образованием мозгового ствола красные ядра регулируют мышечный тонус.
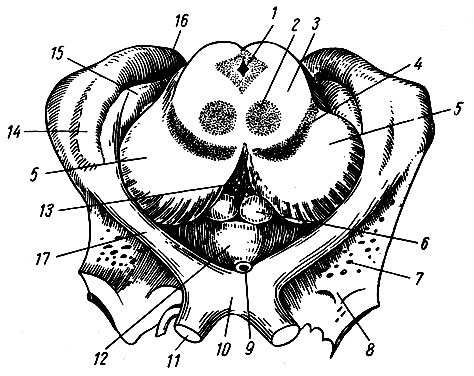
Рис. 203. Забугорная и подбугорная области. 1 - водопровод большого мозга; 2 - красное ядро; 3 - покрышка; 4 - черное вещество; 5 - ножка мозга; 6 - сосковидное тело; 7 - переднее продырявленное вещество; 8 - обонятельный треугольник; 9 - воронка; 10 - зрительный перекрест; 11 - зрительный нерв; 12 - серый бугор; 13 - заднее продырявленное вещество; 14 - боковое коленчатое тело; 15 - медиальное коленчатое тело; 16 - подушка; 17 - зрительный тракт
Регуляцию мышечного тонуса осуществляет также черное вещество, substantia nigra, ножек мозга. Черное вещество образовано клетками, которые содержат черный пигмент - меланин. Оно является границей crus cerebri и покрышки среднего мозга и прослеживается на всем его протяжении. Наряду с регуляцией мышечного тонуса черное вещество участвует также в координации сложных двигательных актов - жевания и глотания.
Белое вещество среднего мозга образовано короткими и длинными эндогенными волокнами и длинными экзогенными.
Crus cerebri образуют длинные экзогенные эфферентные волокна, среди которых 3/5 принадлежит пирамидным путям, занимающим центральную часть этого образования. Кнаружи и кнутри от пирамидного пути расположены корково-мостовые тракты.
В покрышке среднего мозга проходят различные восходящие и нисходящие пучки волокон, среди которых длинными афферентными экзогенными являются волокна, образующие латеральную и медиальную петли. Длинные эндогенные возникают в ядрах среднего мозга: красном ядре (красноядерно-спинномозговой путь), ядрах крыши (покрышечно-спинно-мозговой путь), и др., а короткие эндогенные волокна соединяют ядра среднего мозга.
Промежуточный мозг
Промежуточный мозг, diencephalon, - образования конечного отдела мозгового ствола, которые, как и полушария головного мозга, развиваются из переднего мозгового пузыря. В связи с развитием и функцией образований, входящих в состав промежуточного мозга, его подразделяют на филогенетически старую подбугорную область, hypothalamus, - высший подкорковый центр автономной нервной системы, и филогенетически новую область - зрительный мозг, thalamencephalon, - центр афферентных проводящих путей. Полостью промежуточного мозга является III желудочек.
К подбугорной области, hypothalamus, относят образования вентрального отдела промежуточного мозга, которые входят в состав нижней стенки или дна III желудочка. В подбугорной области различают следующие образования головного мозга: серый бугор с воронкой, гипофиз, конечную пластинку, перекрест зрительных нервов, зрительные пути и сосковидные тела (см. рис. 203).
Серый бугор, tuber cinereum, представляет собой серого цвета возвышение, расположенное между перекрестом зрительных нервов спереди и сосковидными телами сзади. Вершина возвышения, суживаясь, образует воронку, infundibulum, которая переходит в придаток мозга - гипофиз, hypophysis cerebri. Воронке со стороны дна III желудочка соответствует углубление, recessus infundibuli (см. рис. 197). Серый бугор и воронка образованы тонкой пластинкой серого вещества, которая кпереди постепенно истончается и переходит в конечную пластинку, lamina terminalis.
Зрительный перекрест, chiasma opticum, состоит главным образом из волокон зрительных нервов, при неполном перекресте которых образуется примерно четырехугольной формы пластинка. Задние углы этой пластинки продолжаются в зрительные тракты, tractus opticus. Каждый зрительный тракт огибает ножку мозга и заканчивается в подкорковых зрительных центрах: латеральном коленчатом теле, подушке зрительного бугра и верхнем холмике крыши среднего мозга. Кроме волокон зрительных нервов, в зрительном перекресте и зрительных трактах расположены комиссуральные волокна, которые соединяют медиальные коленчатые тела, верхний холмик крыши среднего мозга с подбугорной областью противоположной стороны, а также hypothalamus правого и левого полушарий. Области зрительного перекреста со стороны дна III желудочка соответствует зрительное углубление, recessus opticus.
Сосковидные тела, corpora mamillaria, расположены между серым бугром и задним продырявленным веществом в виде двух белого цвета округлых образований величиной с горошину. Внутри каждого из сосковидных тел под слоем белого вещества находятся два ядра - латеральное и медиальное. В сосковидных телах заканчиваются волокна столбиков свода, columnae fornicis, а также волокна из покрышки моста. От медиальных ядер сосковидных тел возникают пучки волокон к передним ядрам зрительных бугров - сосково-бугорный пучок, fasciculus mamillothalamicus, и к ядрам покрышки моста - сосково-покрышечный пучок, fasciculus mamillotegmentalis.
В сером веществе подбугорной области выделяют 32 пары ядер, которые являются высшими подкорковыми центрами регуляции деятельности симпатического и парасимпатического отделов автономной нервной системы, секреторных функций желез внутренней секреции, терморегуляции, обмена веществ и др. В то же время функция ядер подбугорной области находится под контролем коры полушарий большого мозга.
Зрительный мозг
В зрительном мозге, thalamencephalon, различают три отдела: средний отдел - зрительный бугор, thalamus, верхний отдел - надбугорную область, epithalamus, задний отдел - забугорную область, metathalamus (рис. 193, 204).
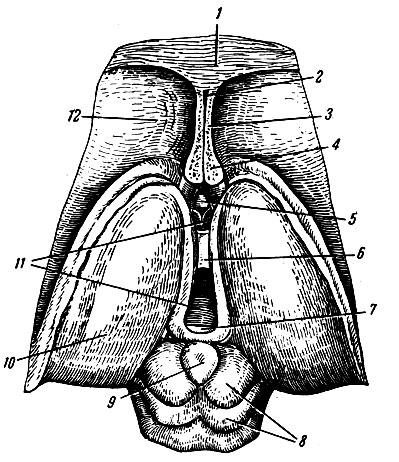
Рис. 204. Промежуточный и средний мозг (вид сверху). 1 - мозолистое тело; 2 - полость прозрачной перегородки; 3 - прозрачная перегородка; 4 - свод (поперечный разрез столбиков); 5 - передняя спайка; 6 - межбугорное сращение; 7 - задняя спайка; 8 - крыша среднего мозга; 9 - шишковидное тело? 10 - зрительный бугор; 11 - III желудочек; 12 - хвостатое ядро (головка)
Зрительный бугор, thalamus, представляет собой парное массивное образование яйцевидной формы, на переднем конце которого имеется передний бугорок, tuberculum anterius; задний утолщенный конец носит название подушки, puluinar. Из четырех поверхностей зрительного бугра латеральная прилегает к внутренней капсуле, capsula interna, вентральиая - соединяется с подбугорной областью, а дорсальная и медиальная - свободны. Дорсальная поверхность зрительного бугра выпукла и покрыта тонким слоем белого вещества, который называют' поясным слоем, stratum zonale. Эта поверхность зрительного бугра снаружи отграничена пограничной полоской, stria terminalis, которая отделяет ее от хвостатого ядра, nucleus caudatus, а снутри - мозговой полоской бугра, stria medullaris thalami.
Медиальная поверхность зрительного бугра серого цвета. Снизу от подбугорной области она отделяется подбугорной бороздой, sulcus hypothalamics, которая проходит от межжелудочкового отверстия, foramen interventricular, ко входу в водопровод большого мозга. Медиальные поверхности зрительных бугров соединены между собой межбугорным сращением, adhesio interthalamica.
Зрительный бугор представляет собой наиболее крупное скопление серого вещества стволовой части мозга. На его разрезах, проведенных в разных плоскостях, видно, что вся масса серого вещества разделена полосками белого вещества - мозговыми пластинками бугра, lamina medullaris thalami, на несколько ядер различной величины. Выделяют группу передних и вентральных ядер, латеральное, медиальное и другие ядра зрительного бугра.
Передние ядра расположены в области переднего конца и переднего бугорка зрительного бугра. Здесь заканчивается сосково-бугорный пучок, fasciculus mamillothalamicus, связывающий зрительный бугор с обонятельной системой. Крупное медиальное ядро зрительного бугра связано с вегетативными ядрами подбугорной области и корой больших полушарий. Значительные размеры имеет боковое ядро зрительного бугра, которое кзади без резких границ переходит в подушку. Это ядро считают ассоциативным ядром зрительного бугра, по волокнам которого передаются импульсы от других ядер зрительного бугра в кору полушарий головного мозга. Вентральные ядра зрительного бугра расположены ниже латерального ядра и отделены от него мозговыми пластинками. В них заканчиваются часть волокон верхней мозжечковой ножки, волокна медиальной петли, петля тройничного нерва, lemniscus trigeminus. Волокна вентральных ядер связаны с корой задней центральной извилины как непосредственно, так и через латеральное ядро зрительного бугра.
Главное функциональное значение зрительных бугров заключается в том, что они являются коллектором чувствительных путей, по которым к коре больших полушарий следуют импульсы от экстеро- и интерорецепторов.
Забугорная область, metathalamus, состоит из парных образований - коленчатых тел. Латеральное коленчатое тело, corpus geniculatum laterale, в виде небольшого удлиненного бугорка расположено под подушкой зрительного бугра. Оно состоит из чередующихся слоев серого и белого вещества и является переключательным ядром зрительного пути. Здесь оканчиваются волокна латерального корешка зрительного тракта. Отростки клеток этого ядра вместе с волокнами от подушки зрительного бугра образуют зрительную лучистость, radiatio optica. Этот пучок волокон заканчивается в коре шпорной борозды, sulcus calcarinus, затылочной доли мозга.
Медиальное коленчатое тело, corpus geniculatum mediate, больше латерального. Оно имеет форму вытянутого валика, который расположен между верхними холмиками крыши среднего мозга и подушкой зрительного бугра. В клетках, образующих ядро медиального коленчатого тела, заканчиваются волокна латеральной (слуховой) петли. Волокна от клеток этого ядра, которое является подкорковым переключательным центром слухового пути, заканчиваются в коре верхней височной извилины.
Надбугорная область, epithalamus, состоит из нескольких образований, расположенных над зрительными буграми. К ним относятся мозговые полоски, striae medullares, которые разделяют дорсальную и медиальную поверхности зрительных бугров. Треугольной формы расширение этих полосок образует треугольник поводка, trigonum habenulae, - белый пучок волокон, отходящий от поводка, habenula. Правый и левый поводки соединены между собой спайкой, commissura habenularum. Кзади каждый поводок переходит в мозговую пластинку, которая соединяет их с шишковидным телом, corpus pineale, - железой внутренней секреции. От основания железы, вентрально к верхним бугоркам крыши среднего мозга, проходит вторая мозговая пластинка, в месте перегиба которой находится задняя мозговая спайка, commissura posterior.
Третий желудочек
Третий желудочек, ventriculus tertius, является полостью промежуточного мозга и представляет собой узкую вертикальную щель, которая находится между медиальными поверхностями зрительных бугров. Боковыми стенками III желудочка являются медиальные поверхности зрительных бугров и образования подбугорной области. Нижней стенкой, или дном III желудочка являются передние участки ножек мозга, заднее продырявленное вещество, сосковидные тела, серый бугор с воронкой и зрительный перекрест. В области дна III желудочка имеются два кармана: углубление воронки, recessus infundibuli, и зрительное углубление, recessus opticus.
Переднюю стенку желудочка создают конечная пластинка, столбики свода и передняя мозговая спайка. Между изгибом столбиков свода и зрительным бугром находится овальной формы межжелудочковое отверстие, foramen interventriculare, которое соединяет полость бокового и III желудочков. Пространство между столбиками свода и передней мозговой спайкой носит название recessus triangularis. Задняя стенка желудочка образована спайкой поводков, основанием шишковидной железы и задней мозговой спайкой. В области основания эпифиза находится углубление шишковидного тела, recessus pinealis.
Верхняя стенка, или крыша, III желудочка расположена между зрительными буграми под сводом и мозолистым телом. Она имеет вид тонкой пластинки, называемой сосудистой основой III желудочка, которая прикрепляется к внутреннему краю мозговой полоски зрительного бугра, а сзади - к верхней поверхности поводков и шишковидной железе, образуя здесь надшишковидное углубление.
Конечный мозг
Конечный мозг, telencephalon, является самым крупным отделом центральной нервной системы, значительно превышающим по своему объему стволовую часть головного мозга, которую он покрывает (рис. 205). В образованиях конечного мозга сосредоточены центры, которые управляют деятельностью различных отделов мозгового ствола и спинного мозга. Кора полушарий головного мозга осуществляет высшую нервную деятельность и определяет поведение организма в зависимости от беспрерывно изменяющихся условий внешней среды. Если в низших отделах нервной системы рефлекторные реакции протекают по наследственным нервным путям, то в коре больших полушарий эти нервные связи создаются в процессе индивидуальной жизни организма под воздействием множества поступающих в нее раздражений от рецепторных приборов.
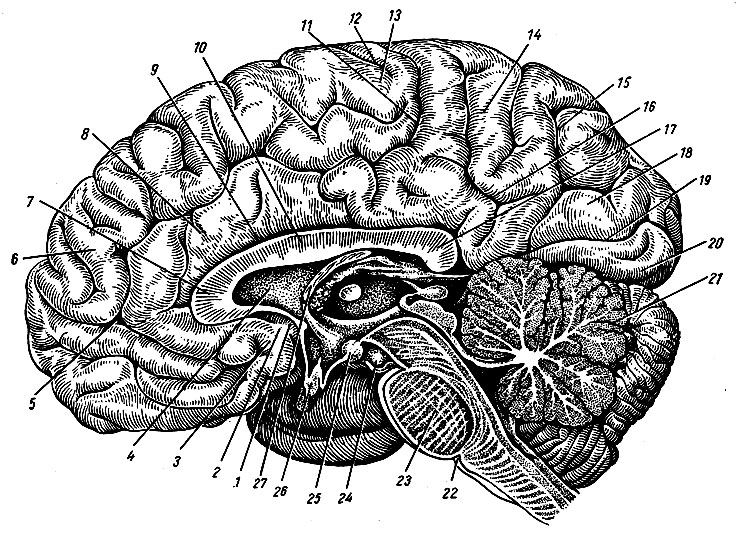
Рис. 205. Внутренняя поверхность правого полушария мозга, стволовой части и мозжечка. Водопровод мозга, IV желудочек. 1 - подмозолистая извилина; 2 - задняя обонятельная борозда; 3 - обонятельная площадка; 4 - прозрачная перегородка; 5, 11 - борозда пояса; 6 - верхняя лобная извилина; 7 - колено мозолистого тела; 8 - извилина пояса; 9 - борозда мозолистого тела; 10 - ствол мозолистого тела; 12 - центральная борозда; 13 - околоцентральная долька; 14 - предклинье; 15 - теменно-затылочная борозда; 16 - подтеменная борозда; 17 - валик мозолистого тела; 18 - клин; 19 - шпорная борозда; 20 - медиальная затылочно-височная извилина; 21 - долька нижнего червя; 22 - задняя граница моста; 23 - мост; 24 - глазодвигательный нерв; 25 - сосцевидное тело; 26 - зрительный перекрест; 27 - свод
Конечный мозг составляют правое и левое полушария, hemispherii dextrum et sinistrum, которые соединены между собой мозолистым телом, corpus callosum, и другими спайками или комиссурами (передняя спайка, спайка свода). Все перечисленные образования - мозолистое тело, а также расположенный под ним свод, fornix, передняя спайка и спайка свода - относятся к конечному мозгу.
В каждом полушарии различают плащ, или мантию, pallium, обонятельный мозг, rhinencephalon, и подкорковые ядра.
Плащ
Форма полушарий головного мозга соответствует форме черепа. Каждое из них имеет три поверхности: сферическую - верхне-боковую, fades superolateral hemispherii, сравнительно плоскую - медиальную, fades medialis hemispherii, и довольно сложной формы нижнюю поверхность, fades inferior hemispherii. Перечисленные поверхности полушария разделяют края: верхний (верхне-медиальный), нижний (нижне-боковой) и средний (нижне-медиальный). Передний выступающий участок полушария называют лобным полюсом, polus frontalis, задний - затылочным полюсом, polus occipitalis, а нижнюю заостренную часть бокового отдела - височным полюсом, polus temporalis.
Поверхность полушарий разделяется различной глубины и длины бороздами большого мозга, sulci cerebri, на большое число извилин. Извилиной большого мозга, gyrus cerebri, называют участок мозгового вещества в форме валика, который ограничен двумя бороздами. Все борозды полушарий подразделяют на три группы: первичные, вторичные и третичные, из которых первичные борозды глубоки, постоянно встречаются и рано появляются в онтогенезе. Вторичные борозды также довольно постоянны, но появляются позже первичных, а третичные - непостоянны и крайне изменчивы. Глубокие первичные борозды делят каждое полушарие на большие участки, называемые долями большого мозга. В каждом полушарии имеется пять долей: лобная, lobus frontalis, теменная, lobus parietalis, височная, lobus temporalis, затылочная, lobus occipitalis, и островок, insula, который скрыт в глубине боковой борозды. Поверхность полушарий как в глубине борозд, так и на вершине извилин представляет собой значительный по толщине слой серого вещества, который называют корой большого мозга, cortex cerebri. Толщина коры в различных участках полушарий составляет около 3 мм. Вследствие наличия многочисленных борозд кора полушарий головного мозга имеет складчатый вид.
Борозды и извилины верхне-боковой поверхности полушария. Самой глубокой бороздой верхне-боковой поверхности полушария является боковая борозда, sulcus lateralis. Она прослеживается от основания мозга в виде глубокой щели, которая разделяется на три ветви: две короткие - переднюю, ramus anterior, восходящую, ramus ascendens и длинную, направленную назад и вверх, заднюю, ramus posterior. Задняя ветвь заканчивается примерно на границе средней и задней третей верхне-боковой поверхности полушария (рис. 206). Несколько кзади от середины полушария с медиальной поверхности, сверху вниз и сзади наперед проходит центральная борозда, sulcus centralis, которая, как правило, не достигает боковой борозды. Наиболее глубоким участком центральной борозды является ее средняя часть.
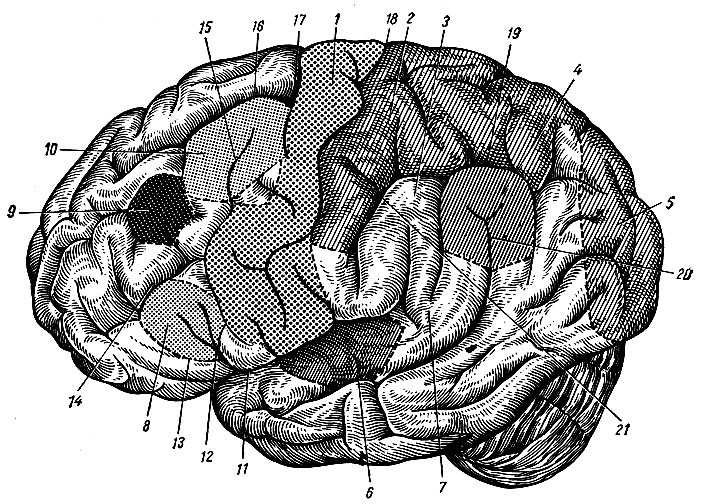
Рис. 206. Верхне-боковая поверхность левого полушария. 1 - ядро двигательного анализатора; 2 - ядро кожного анализатора; 3 - ядро двигательного анализатора, посредством которого синтезируются привычные целенаправленные движения; 4 - ядро зрительного анализатора письменной речи; 5 - ядро зрительного анализатора (зрительная память); 6 - ядро слухового анализатора; 7- ядро слухового анализатора речи; 8 - ядро двигательного анализатора артикуляции речи; 9 - ядро двигательного анализатора, имеющего отношение к сочетанному повороту головы и глаз; 10 - ядро двигательного анализатора письменной речи; 11 - боковая борозда мозга; 12 - ее восходящая ветвь; 13 - ее передняя ветвь; 14 - нижняя лобная борозда; 15 - нижняя предцентральная борозда; 16 - верхняя лобная борозда; 17 - верхняя предцентральная борозда; 18 - центральная борозда; 19 - внутритеменная борозда; 20 - верхняя височная борозда; 21 - зацентральная борозда
Участок полушария, расположенный кпереди от центральной и кверху от боковой борозды, относится к лобной доле. Позади центральной борозды находится теменная доля. Ее ограничивают спереди центральная борозда, снизу - задняя ветвь боковой борозды; сзади границей являются две борозды: сверху - теменно-затылочная, sulcus pariet о occipitalis, снизу - поперечная затылочная, sulcus occipitalis transversus. Книзу от боковой борозды расположена височная доля, кзади переходящая в затылочную. Границей этих долей считают линию, которую проводят от теменно-затылочной борозды к нижнему краю полушария. Затылочная доля расположена позади теменно-затылочной и поперечной затылочной борозд. Ее границей с височной долей считают линию, соответствующую продолжению книзу поперечной затылочной борозды.
Лобная доля. Параллельно центральной борозде в задней части лобной доли проходит прецентральная борозда, sulcus precentralis. От этой борозды почти под прямым углом в продольном направлении идут верхняя и нижняя лобные борозды, sulci frontales superior et inferior. Указанные борозды делят лобную долю на 4 извилины: прецентральную, gyrus precentralis, верхнюю лобную, gyrus frontalis superior, среднюю лобную, gyrus frontalis medius, и нижнюю лобную, gyrus frontalis inferior.
Теменная доля. Позади центральной борозды и параллельно ей проходит зацентральная борозда, sulcus post centralis. Верхний конец этой борозды находится у верхнего края полушария, а нижний - часто переходит во внутритеменную борозду, sulcus intraparietalis, которая расположена параллельно верхнему краю полушария. Центральная и зацентральная борозды ограничивают вертикальную зацентралъную извилину, gyrus postcentralis. Остальная часть теменной доли делится внутритеменной бороздой на верхнюю и нижнюю теменные дольки, lobuli parietales superior et inferior. Нижняя теменная долька огибает концы задней ветви сильвиевой и верхней височной борозд. Участок этой дольки, огибающий конец задней ветви боковой борозды, называют надкраевой извилиной, gyrus supramarginalis, а участок, огибающий конец верхней височной борозды, угловой извилиной, gyrus angularis.
Височная доля. На латеральной поверхности височной доли имеются две продольные - верхняя и нижняя височные борозды, sulci temporales superior et inferior. Направление этих борозд примерно соответствует боковой борозде. Между боковой и верхней височной бороздой расположена верхняя височная извилина, gyrus temporalis superior, на верхней поверхности которой имеются две или три поперечные височные извилины, gyri temporales transversi. Верхняя и нижняя височные борозды ограничивают среднюю височную извилину, gyrus temporalis medius, которая соединяется с нижерасположенной нижней височной извилиной несколькими переходными извилинами. Нижняя височная извилина, gyrus temporalis inferior, расположена на границе верхне-боковой поверхности и основанием мозга.
Затылочная доля. Борозды латеральной поверхности затылочной доли непостоянны по количеству и направлению. Поперечные затылочные борозды, sulci occipitales transversi, разделяют латеральную поверхность этой доли на одноименные извилины. На нижне-боковой поверхности имеется вдавление - пред затылочная вырезка, incisura preoccipitalis. Это вдавление служит ориентиром для проведения условной границы между затылочной долей сзади и теменной и височной долями спереди.
Островок. Островок, insula, расположен в глубине боковой борозды и образует ее дно. Он представляет собой треугольной формы выступ, вершина которого направлена кнаружи и вниз. Островок окружен круговой бороздой островка, sulcus circularis insulae. Поверхность островка делится центральной бороздой островка, sulcus centralis insulae, на переднюю и заднюю доли. Неглубокие борозды делят поверхность островка на 5-7 извилин, из которых короткие, gyri breves insulae, находятся в передней доле, а длинная, gyrus longus insulae, в задней доле. Покрывающие островок края боковой борозды называют покрышкой, operculum, которая разделяется на лобную, operculum frontale, лобно-теменную, operculum frontoparietal, и височную, operculum temporale.
Борозды и извилины нижней поверхности полушария. Участок нижней поверхности полушария, fades inferior hemispherii, расположенный кпереди от боковой борозды, относится к лобной доле. Здесь в сагиттальном направлении параллельно продольной щели мозга проходит обонятельная борозда, sulcus olfactorius. Обонятельная борозда прикрыта обонятельной луковицей, bulbus olfactorius, и обонятельным путем, tractus olfactorius. Между обонятельной бороздой и продольной щелью мозга расположена прямая извилина, gyrus rectus. Кнаружи от обонятельной борозды несколько глазничных бороздок, sulci orbitales, ограничивают gyri orbitales, которые занимают остальную часть базальной поверхности лобной доли.
Участок нижней поверхности, расположенный позади боковой борозды, относится к височной и -затылочной долям. Наружной бороздой нижней поверхности височной доли является затылочно-височная, sulcus occipitotemporalis, которая вместе с нижней височной бороздой ограничивает нижнюю височную извилину, gyrus temporalis inferior. Медиальнее затылочно-височной борозды и почти параллельно ей находится глубокая окольная борозда, sulcus collateralis. Между этими бороздами расположена боковая затылочно-височная извилина, gyrus occipitotemporalis lateralis. Кнутри от окольной борозды находится медиальная затылочно-височная извилина, gyrus occipitotemporalis medialis. Эту извилину ограничивают окольная и шпорная, sulcus calcarinus, борозды. Латеральная и медиальная затылочно-височные извилины соединены переходными извилинами с парагиппокампальной извилиной, gyrus parahippocampalis.
Борозды и извилины медиальной поверхности полушария. На медиальной поверхности полушария, fades medialis hemispherii, непосредственно над мозолистым телом имеется борозда мозолистого тела, sulcus corporis callosi, которая огибает все отделы мозолистого тела и переходит в глубокую борозду гиппокампа, sulcus hippocampi (см. рис. 205). Выше борозды мозолистого тела проходит вторая дугообразная борозда, которую называют бороздой пояса, sulcus cinguli. Эта борозда начинается под клювом мозолистого тела, идет назад и заканчивается на верхнем крае полушария несколько кзади от верхнего конца центральной борозды. Непосредственным продолжением sulcus cinguli кзади является подтеменная борозда, sulcus subparietalis. Над бороздой пояса в передней половине полушария расположен медиальный отдел верхней лобной извилины, задней границей которой на этой поверхности считают околоцентральную борозду, sulcus paracentralls. Между околоцентральной бороздой и задним концом борозды пояса расположена околоцентральная долька, lobulus paracentralis. Кзади от этой дольки находится четырехугольной формы образование - предклинье, precuneus, которое ограничено спереди концом борозды пояса, снизу - подтеменной бороздой, sulcus subparietalis, сзади - теменно-затылочной бороздой. Позади предклинья расположен клин, cuneus, который относится к затылочной доле. Клин ограничен спереди теменно-затылочной, а сзади - шпорной бороздой. Борозда пояса и борозда мозолистого тела ограничивают поясную извилину, gyrus cinguli, суженное место которой - перешеек, isthmus, продолжается в gyrus hippocampi. Эта извилина образует направленный кзади изгиб, который называют крючком, uncus. Извилину гиппокампа ограничивает о внутренней стороны борозда гиппокампа, sulcus hippocampi, а с наружной - sulcus collateralis. Суженное место перехода извилины пояса в извилину гиппокампа расположено позади валика мозолистого тела, splenium corporis callosi. В борозде гиппокампа находится зубчатая извилина, gyrus dentatus, которая представляет собой серого цвета зазубренную полоску.
Строение коры полушарий головного мозга. Кору полушарий головного мозга образуют 10-14 млрд. разнообразных по форме, величине и функции нервных клеток, которые расположены слоями. Имеется и вертикальная организация нейроцитов коры в виде колонн. Принято различать 6 слоев коры: 1) молекулярный, 2) наружный зернистый, 3) слой пирамидных клеток, 4) внутренний зернистый, 5) ганглионарный, 6) слой полиморфных клеток.
Кора большого мозга в целом является рецепторной, воспринимающей структурой, куда, как на экран, проецируются возникающие в рецептор-ном аппарате нервные импульсы. Во все области коры вступает множество афферентных волокон. Эфферентные волокна идут к различным подкорковым ядрам, двигательным ядрам мозгового ствола и спинного мозга и к другим нейронам коры, не выходя за ее пределы.
С точки зрения истории развития, строения и функции кора полушарий большого мозга делится на зону новой коры, которая занимает 95,6% общей площади, зону старой и древней коры с их межуточными зонами. Древняя и старая кора в функциональном отношении связана главным образом с обонятельной функцией - самой древней функцией конечного мозга.
Особенности топографии клеточных структур коры, их величина, форма, плотность расположения, ширина различных слоев и др. послужили основой для выделения в коре ряда цитоархитектонических областей, а в пределах областей - цитоархитектонических полей. Различают следующие цитоархитектонические области: затылочную, нижнюю теменную, верхнюю теменную, постцентральную, прецентральную, лобную, височную, островковую и лимбическую (лимбическая область коры: извилина пояса, перешеек и извилина гиппокампа). Каждая из этих областей подразделяется на различное число полей, которых во всей коре насчитывают более 50.
Учение о цитоархитектонике коры полушарий головного мозга соответствует учению И. П. Павлова о коре как о системе корковых концов анализаторов. Анализатор, по И. П. Павлову, "есть сложный нервный механизм, начинающийся наружным воспринимающим аппаратом и кончающийся в мозгу". И. П. Павлов показал, что все отделы анализаторов, начиная с рецепторного прибора, нервных центров и кончая корковым концом, анализируют внешние и внутренние раздражения, причем в корковом конце анализатора осуществляются наиболее тонкие формы анализа и особо сложный синтез этих раздражений. Корковый конец анализатора, по И. П. Павлову, состоит из "ядра" и "рассеянных элементов". Ядро анализатора по структурным и функциональным особенностям подразделяют на центральное поле ядерной зоны и периферическое. Считают, что в первом формируются тонко дифференцированные ощущения, а во втором - более сложные формы отражения внешнего мира. Рассеянные элементы коркового конца анализатора представляют собой те нейроны, которые расположены за пределами ядра и осуществляют более простые функции. Локализация ядер основных анализаторов следующая (рис. 207).
Рис. 207. Карта цитоархитектонических полей мозга человека (по данным Московского института мозга). Вверху - наружная поверхность (пояснение в тексте). Внизу внутренняя поверхность (пояснение в тексте)
1. Предцентральная извилина и передний отдел околоцентральной дольки входят в состав прецентральной области - двигательной или моторной зоны коры (цитоархитектонические поля 4, 6). Афферентные клетки этих полей воспринимают проприоцептивные импульсы от мышц, сухожилий, суставов, а двигательные клетки 5-го и 6-го слоев образуют основную массу волокон двигательного произвольного пути. Эти волокна связывают кору полей 4 и 6 с двигательными ядрами черепных нервов и ядрами передних рогов спинного мозга. Группа нейронов, которые осуществляют иннервацию определенных мышц или мышечных групп, расположены в такой последовательности: в верхних отделах находятся центры для мышц нижней конечности, затем туловища, далее мышц верхней конечности и в нижнем отделе - мышц головы. Наибольшую площадь всей зоны занимают центры иннервации кисти руки, лица, губ, языка, а меньшую площадь - центры иннервации мышц туловища и нижних конечностей. Центры двигательной зоны осуществляют иннервацию противоположной части тела.
2. В задней части средней лобной извилины (поле 8) и на границе теменной и затылочной долей (поле 19) расположены центральные отделы двигательного анализатора, которые осуществляют координацию поворота головы и глаз в противоположную сторону. Участки коры (поле 6 и 8), расположенные кпереди от моторной зоны, называют премоторной зоной. Отростки клеток этой зоны связаны как с ядрами передних рогов спинного мозга, так и с подкорковыми ядрами, красным ядром, черной субстанцией и др.
3. В зацентральной извилине (поля 1, 2, 3, 5) находится ядро кожного анализатора (центры температурной, болевой, осязательной, мышечно-суставной чувствительности). Последовательность расположения центров и их территория соответствуют моторной зоне коры.
4. Корковые концы интероцептивного анализатора, расположены в премоторной зоне (поля 6 и 8). и лимбической области коры. Эфферентные пути из этих участков коры идут к ядрам гипоталамуса и другим центрам автономной системы.
5. В области верхней височной извилины расположено ядро слухового анализатора (поле 41). Здесь проецируются волокна, которые проводят импульсы от рецепторного аппарата, расположенного в улитке внутреннего уха.
6. Корковый конец зрительного анализатора расположен в затылочной доле мозга (поля 17, 18, 19). Ядро зрительного анализатора находится в области шпорной борозды (поле 17). Здесь спроецирована сетчатка латеральной1 половины одного глаза и медиальной половины другого. Поэтому полная слепота возникает при двустороннем поражении ядер зрительного анализатора. В случаях поражения полей 17 и 18 наступает потеря зрительной памяти. При поражении поля 19 человек утрачивает способность к ориентировке в новой для него обстановке.
7. В области крючка и гиппокампа расположено ядро обонятельного анализатора.
8. В нижнем отделе зацентральной извилины находится ядро вкусового анализатора.
9. Поля лимбической области коры благодаря многочисленным связям с другими ее областями выполняют важную роль в синтезе афферентных раздражений.
10. В левой нижней теменной дольке, gyrus supramargmalis (поле 40), у правшей находится центр целенаправленных комбинированных движений. Такие сложные движения вырабатываются путем обучения и практики в течение индивидуальной жизни, путем образования и закреплениявременных связей. При поражении поля 40 больной, несмотря на отсутствие явлений паралича, теряет способность пользоваться предметами обихода, утрачивает производственные навыки, что называют апраксией.
11. В правой и левой верхних теменных дольках (поле 7) находятся центры особого сложного вида кожной чувствительности - стереогностического чувства, т. е. способности узнавания предметов на ощупь. При поражении теменной дольки больной не может узнать предмет, ощупывая его рукой, противоположной очагу поражения. Кроме стереогнозии, различают слуховую гнозию - узнавание предметов по звуку (птицу - по голосу, автомобиль - по шуму мотора), зрительную гнозию - узнавание предметов по виду и др. Праксия и гнозия являются функциями высшего порядка, осуществление которых связано как с первой, так и со второй сигнальной системой. Эти функции являются специфическими функциями человека.
Одной из филогенетически новых функций коры является развитие высокоспециализированных анализаторов речи, которая появилась в ходе социального развития человека, в результате трудовой деятельности. "Сначала труд, а затем и вместе с ним членораздельная речь явились двумя самыми главными стимулами, под влиянием которых мозг обезьяны постепенно превратился в человеческий мозг, который при всем своем сходстве с обезьяньим далеко превосходит его по величине и совершенству*.
*(К. Маркс и Ф. Энгельс. Сочинения. Т. 20. Изд. 2-е. М., 1961, с. 490.)
Функция речи крайне сложна. Она не может быть узко локализована в каком-либо участке коры. В осуществлении этой сложнейшей функции участвуют обширные территории коры. У большинства людей-правшей речевые функции, функции узнавания (гнозия), целенаправленного действия (праксия) связаны с определенными цитоархитектоническими полями левого полушария, а у левшей - наоборот.
12. В задней части нижней лобной извилины (поле 44), у правшей в левом полушарии расположен двигательный анализатор артикуляции речи (речедвигательный анализатор). Здесь осуществляется анализ раздражений от двигательного аппарата, посредством которого образуются слоги, слова и осуществляется сложная функция сочетания движений этого аппарата. Этот центр образовался рядом с проекционной областью двигательного анализатора для мышц губ, языка, гортани. При поражении этого центра человек способен произносить отдельные речевые звуки, но способность образовать из этих звуков слова он утрачивает (двигательная или моторная афазия). При поражении поля 45 наблюдается аграмматизм - больной утрачивает способность составлять из слов предложения, согласовать слова в предложении.
13. Тесно связан с речедвигательным анализатором слуховой анализатор устной речи, который расположен в задней части верхней височной извилины, (поле 42). При поражении этого центра утрачивается связь между словом как сигналом определенного представления, предмета иди понятия. Больной не понимает значения слов, хотя слышит их (сенсорная афазия).
14. В заднем отделе средней лобной извилины (поле 8) рядом с ядром двигательного анализатора поворота глаз и головы в противоположную сторону расположено ядро двигательного анализатора письменной речи - графии. Оба анализатора связаны между собой, но центр графии тесно связан и с полем 40, расположенным в gyrus supramarginalis. При повреждении этой области больной не может производить двоения, которые необходимы для начертания букв.
15. В нижней теменной дольке в области gyrus angularis расположен зрительный анализатор письменной речи (поле 39). При повреждении этого поля больной утрачивает способность анализа написанных букв, т. е. теряет способность читать (алексия).
Обонятельный мозг
К обонятельному мозгу, rhinencephalon, относятся: обонятельная луковица, bulbus olfactorius, обонятельный тракт, tractus olfactorius, обонятельный треугольник, trigonum olfactorium, переднее продырявленное вещество, substantia perforata anterior, извилина пояса, gyrus cinguli, перешеек, isthmus, парагиппокампалъная извилина, gyrus parahippocampalis, зубчатая извилина, gyrus dentatus, и гиппокамп, hippocampus.
Извилина пояса, перешеек, парагиппокампальная извилина, гиппокамп, зубчатая извилина, свод и миндалевидное тело входят в состав лимбической системы. Считают, что эта система выполняет не только обонятельную функцию, но имеет также важное значение в процессах синтеза афферентных раздражений, эмоциональных реакций и реакций так называемого врожденного поведения (половые, поисковые и оборонительные). О связях в системе образований обонятельного мозга см. Проводящие пути обонятельного анализатора.
Подкорковые ядра
Подкорковые ядра расположены в белом веществе полушарий головного мозга ближе к их основанию, в связи с чем эти ядра иногда называют базальными. Они образуют три парных скопления серого вещества: полосатое тело, corpus striatum, миндалевидное тело, corpus amygdaloideum, и ограду, claustrum (рис. 208).
Рис. 208. Фронтальный разрез полушарий, проходящий черз полосатое тело и зрительный бугор. 1 - свод; 2 - сосудистое сплетение III желудочка; 3 - сосудистое сплетение бокового желудочка; 4 - боковой желудочек; 5 - хвостатое ядро; 6 - задняя ножка внутренней капсулы; 7 - скорлупа; 8 - бледный шар; 9 - наружная капсула; 10 - ограда; 11 - боковая борозда мозга; 12, 14, 15 - верхняя, средняя и нижняя височные извилины; 13 - островок; 16 - парагиппокампальная извилина; 17 - глазодвигательный нерв; 18 - мост; 19 - ядро сосковидного тела; 20 - зрительный тракт; 21 - III желудочек; 22 - межбугорное сращение; 23-25 - ядра зрительного нерва; 26, 28, 31 - нижняя, средняя и верхняя лобные извилины; 27 - мозолистое тело; 29 - извилина пояса; 30 - борозда пояса
Полосатое тело состоит из двух ядер: хвостатого, nucleus caudatus, и чечевицеобразного, nucleus lentiformis.
Хвостатое ядро представляет собой округлой формы образование, которое в виде дуги окружает зрительный бугор и чечевицеобразное ядро. Резко утолщенный, булавовидный передний отдел хвостатого ядра называют головкой, caput nuclei caudati. Кзади головка истончается и образует тело хвостатого ядра, corpus nuclei caudati, которое переходит в хвост, cauda nuclei caudati. Головка, тело и хвост участвуют в образовании стенок бокового желудочка мозга.
Чечевицеобразное ядро представляет собой крупное пирамидальной формы скопление серого вещества, которое расположено кнаружи от хвостатого ядра и зрительного бугра и отделено от них внутренней капсулой. Выпуклая наружная поверхность чечевицеобразного ядра обращена к островку, а суженная вершина - внутрь и вниз. Чечевицеобразное ядро неоднородно по своему строению и разделяется пластинками белого вещества на три части: наружную, темного цвета - скорлупу, puta-mеп, и две светлые медиальные - бледный шар, globus pallidus. Хвостатое и чечевицеобразное ядра представляют собой главное звено экстрапирамидной системы, основной функцией которой является осуществление сложных безусловнорефлекторных двигательных актов.
Ограда, claustrum, - сравнительно тонкая пластинка серого вещества, которая расположена в белом веществе, разделяющим наружную поверхность, putamen и кору островка. Белое вещество, которое разделяет ограду и скорлупу, называют наружной капсулой, capsula externa, а участок белого вещества между оградой и корой островка - внешней капсулой, capsula extrema. Ограда является сложным образованием, связи которого до настоящего времени мало изучены, а функциональное значение не ясно.
Миндалевидное тело, corpus amygdaloideum, - крупное ядро, которое расположено под скорлупой в переднем отделе височной доли. Это ядро имеет сложное строение и состоит из нескольких ядер, различающихся по клеточному составу. Миндалевидное тело, являющееся подкорковым обонятельным центром, входит в состав лимбической системы (см. раздел Подкорковые ядра, настоящего издания).
Боковые желудочки
Полостями конечного мозга являются боковые желудочки, ventriculi laterales, которые расположены в толще его полушарий (рис. 209). В каждом боковом желудочке различают центральную часть, pars centralis, и три рога: передний, cornu anterius, задний, cornu posterius, и нижний, cornu inferius.
Рис. 209. Боковые желудочки, вскрытые сверху путем удаления части полушарий вместе с мозолистым телом. 1 - передний рог; 2 - хвостатое ядро (головка); 3- межжелудочковое отверстие; 4 - чечевицеобразное ядро (в разрезе); 5 - конечная полоска; 6 - верхняя поверхность зрительного бугра; 7 - гиппокамп; 8 - боковое возвышение; 9 - бахромка гиппокампа; 10 - ножка свода; 11 - задний рог бокового желудочка; 12 - медиальная стенка заднего рога; 13 - птичья шпора; 14, 15 - задние рога; 16 - валик мозолистого тела; 17, 19 - сосудистые сплетения в центральной части бокового желудочка и продолжение его в нижний рог; 18 - спайка свода; 20 - столб свода; 21 - прозрачная перегородка; 22 - полость прозрачной перегородки; 23 - мозолистое тело
Центральная часть в виде узкой горизонтальной щели расположена в теменной доле. Сверху ее ограничивают волокна мозолистого тела, снизу - тело хвостатого ядра, наружный отдел верхней поверхности зрительного бугра и кнутри от него сосудистое сплетение бокового желудочка. Кпереди центральная часть продолжается в передний рог, который находится в лобной доле.
Передний рог ограничен сверху и спереди мозолистым телом, с латеральной стороны и снизу - головкой хвостатого ядра, а с медиальной стороны - прозрачной перегородкой, septum pellucidum, которая представляет собой тонкую пластинку, состоящую из двух листков, между которыми имеется полость, cavum septi pellucidi. Эта пластинка расположена между мозолистым телом и сводом (см. рис. 205). Кзади центральная часть расширяется. Ее продолжением в затылочной доле является задний рог. В височную долю центральная часть продолжается в виде нижнего pord.
Задний рог ограничен слоем нервных волокон мозолистого тела. На его медиальной стенке имеются два валика. Верхний валик называют луковицей заднего рога, bulbus cornus posterioris, которая образуется задним пучком волокон мозолистого тела. Нижний валик носит название птичьей шпоры, calcar avis. Птичья шпора образуется за счет вдавления в полость желудочка вещества затылочной доли.
Нижний рог самый длинный. Он начинается от расширения центральной части и заканчивается в области височного полюса. В образовании его стенок участвуют tapetum и хвост nucleus caudatus. На внутренней поверхности нижнего рога имеется возвышение - гиппокамп, hippocampus, который образуется вследствие вдавления в стенки мозга борозды гиппокампа. Передний, немного утолщенный отдел гиппокампа разделен бороздами на несколько бугорков пальцевидной формы. Вдоль медиального края гиппокампа расположена бахромка, fimbria hippocampi. Гиппокамп представляет собой сместившуюся в полость желудочка часть старой коры. На дне нижнего рога имеется боковое возвышение, eminentia collateralis, которое образуется одноименной бороздой. Межжелудочковые отверстия, foramina interventricularia, соединяют полость боковых желудоков с III желудочком мозга. Это отверстие образуют столбик свода и передний конец зрительного бугра.
Белое вещество полушарий головного мозга
Белое вещество полушарий образовано волокнами проводящих путей конечного мозга, которые группируются в три системы: 1) ассоциационные, или сочетательные, 2) комиссуральные, или спаечные, 3) проекционные.
Ассоциационные волокна соединяют различные участки коры в пределах одного полушария (рис. 210). Эти волокна делятся на короткие и длинные. К коротким волокнам относятся дугообразные волокна, которые соединяют кору двух соседних извилин. Они образуют самый поверхностный слой белого вещества, расположенный непосредственно под корой. Длинные волокна лежат глубже коротких, но по отношению ко всей массе белого вещества полушария они являются поверхностными. Пучки длинных ассоциационных волокон соединяют отдаленные друг от друга участки коры. Известны следующие пучки длинных ассоциационных волокон.
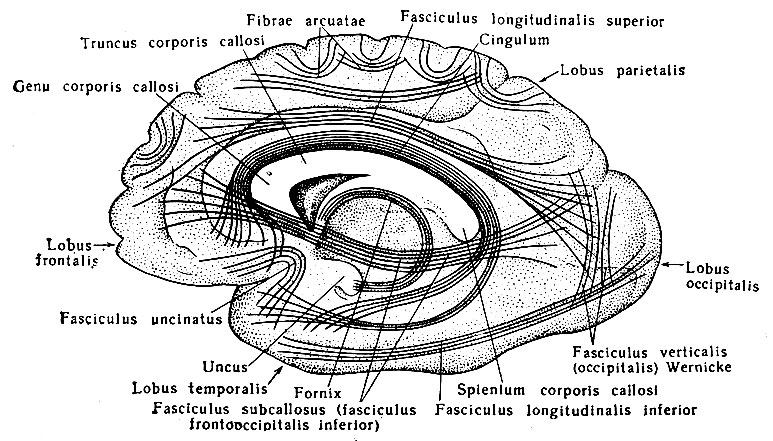
Рис. 210. Схематическое изображение ассоциационных путей мозговых полушарий
1. Пояс, cingulum, прослеживается от переднего продырявленного вещества до извилины гиппокампа; он соединяет кору извилин медиальной поверхности полушария и относится к обонятельному мозгу.
2. Нижний продольный пучок, fasciculus longitudinalis inferior, соединяет затылочную долю с височной. Крупные, различной длины волокна, образуют пучок, расположенный вдоль наружной стенки заднего и нижнего рогов бокового желудочка.
3. Верхний продольный пучок, fasciculus longitudinalis superior, соединяет лобную долю с теменной и височной долями.
4. Крючковатый пучок, fasciculus uncinatus, соединяет прямую и глазничные извилины лобной доли с височной долей.
Комиссуральные нервные пути соединяют области коры обоих полушарий. Они образуют следующие комиссуры, или спайки.
1. Мозолистое тело, corpus callosum, образовано волокнами, соединяющими различные участки новой коры, в связи с чем его величина у человека значительно больше, чем у животных.
Сравнительно тонкая средняя часть мозолистого тела называется стволом, truncus corporis callosi. Кпереди ствол утолщается и изгибается книзу, образуя колено мозолистого тела, genu corporis callosi, нижний конец которого, истончаясь, переходит в клюв, rostrum (см. рис. 197). Задний утолщенный конец мозолистого тела - валик, spienium corporis callosi, несколько изогнут книзу. Вся верхняя поверхность мозолистого тела покрыта тонким слоем серого вещества - серым облачением, indusium griseum.
Вдоль середины верхней поверхности мозолистого тела расположена медиальная продольная полоска, stria longitudinalis medialis, кнаружи от которой лежит боковая продольная полоска, stria longitudinalis lateralis. Серое облачение и продольные полоски представляют собой видоизмененную кору.
Ствол мозолистого тела составляют волокна, которые соединяют кору передних центральных извилин, теменных и височных долей. Эти волокна имеют преимущественно поперечное направление. Колено и клюв мозолистого тела образуют волокна коры передних отделов лобных долей, а валик состоит из волокон затылочных долей. Пучки волокон колена, клюва и валика дугообразно изогнуты и образуют его большие и малые щипцы, forceps major et minor. В состав мозолистого тела, кроме поперечных волокон, соединяющих симметричные участки коры обоих полушарий, входит небольшое количество пучков продольных волокон, которые связывают участки коры разных долей противоположных полушарий.
2. Передняя мозговая спайка, commissura cerebri anterior, и спайка свода, commissura fornicis, являются комиссурами обонятельного мозга. Передняя мозговая спайка расположена позади клюва мозолистого тела. Она состоит из двух пучков; один соединяет переднее продырявленное вещество, а другой - извилины височной доли, преимущественно гиппокампову извилину.
Спайка свода соединяет центральные части двух дугообразных пучков нервных волокон, которые образуют расположенный под мозолистым телом свод, fornix (рис. 211). В своде различают центральную часть - столбы свода, columnae fornices, и ножки свода, crura fornicis. Центральные части соединяет треугольной формы пластинка - спайка свода, commissura fornicis, задний отдел которой сращен с нижней поверхностью мозолистого тела. Столбы свода представляют собой цилиндрической формы образования, которые, изгибаясь кзади, вступают в подбугорную область и заканчиваются в сосковидных телах.
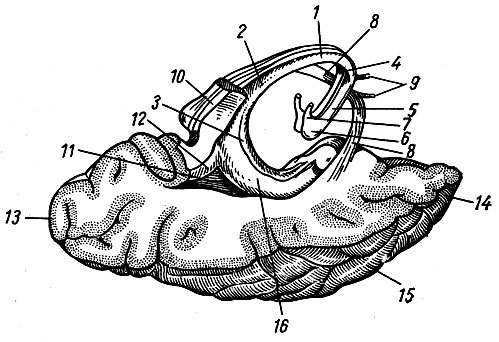
Рис. 211. Свод, гиппокамп и средняя комиссура. 1 - тело свода; 2 - ножка свода; 3 - лента гиппокампа; 4, 5 - ствол свода; 6 - сосковидное тело; 7 - бугорно-сосковидный пучок; 8,9 - передняя спайка; 10 - валик мозолистого тела; 11 - боковой треугольник; 12 - птичья шпора; 13 - затылочный полюс; 14 - височный полюс; 15 - средняя височная извилина; 16 - гиппокамп
Проекционные пути соединяют кору полушарий головного мозга с ядрами мозгового ствола и спинного мозга. Различают эфферентные - двигательные пути и афферентные - чувствительные пути. Двигательные пути проводят нервные импульсы центробежно от клеток двигательных областей коры к двигательным ядрам спинного мозга, ядрам мозгового ствола и подкорковым ядрам. Благодаря этим путям двигательные центры коры головного мозга проецируются на периферию. Посредством чувствительных путей осуществляется проекция поверхности тела в чувствительные и двигательные центры коры. Чувствительные волокна являются отростками клеток спинномозговых узлов и чувствительных узлов черепных нервов. Они образуют первые нейроны чувствительных путей. Отростки клеток спинномозговых узлов и узлов черепных нервов заканчиваются на клетках или дендритах переключательных ядер спинного или продолговатого мозга. От клеток переключательных ядер начинаются вторые нейроны чувствительного пути, которые в составе медиальной петли идут к вентральным ядрам зрительного бугра. В этих ядрах заканчиваются отростки клеток вторых нейронов и начинаются третьи, отростки которых идут в соответствующие ядерные центры коры. Как чувствительные, так и двигательные пути образуют в веществе полушарий систему лучеобразно расходящихся пучков, получившую название лучистого венца, corona radiata. Система волокон, которая образована проводящими путями каждого полушария, собирающаяся в компактный, мощный пучок, заполняющий пространство между зрительным бугром, с одной стороны, хвостатым и чечевицеобразным ядрами - с другой, называют внутренней капсулой, capsula interna (рис. 212). На срезах полушарий головного мозга внутренняя капсула имеет вид довольно широкой полосы белого вещества (см. рис. 208), в которой различают следующие отделы: переднюю ножку, cms anterior, заднюю ножку, crus posterior, и колено, genu.
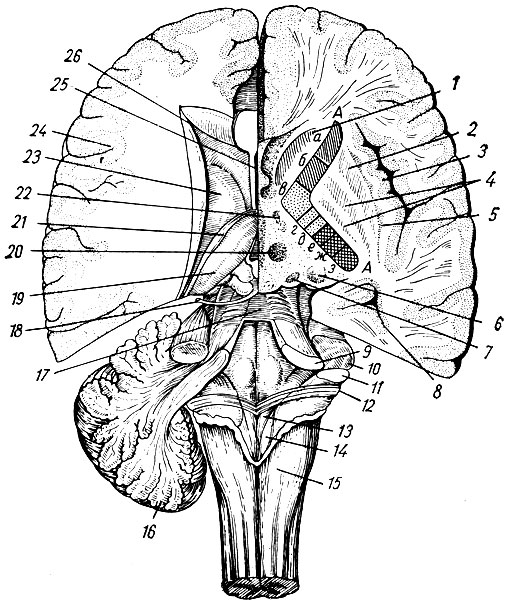
Рис. 212. Полушария мозга на разных уровнях горизонтального разреза. Справа - ниже уровня дна бокового желудочка, слева - над дном бокового желудочка. IV желудочек мозга вскрыт сверху. 1 - головка хвостатого ядра; 2 - скорлупа; 3 - кора островка полушария; 4 - бледный шар; 5 - ограда; 6 - хвост хвостатого ядра; 7 - ядро медиального коленчатого тела; 8 - нижний рог бокового желудочка; 9 - верхняя ножка мозжечка; 10 - средняя ножка мозжечка; 11 - нижняя ножка мозжечка; 12 - мозговые полоски; 13 - треугольник подъязычного нерва; 14 - треугольник блуждающего нерва; 15 - бугорок тонкого ядра; 16 - мозжечок; 17 - верхний мозговой парус; 18 - блоковый нерв; 19 - зрительный бугор; 20 - красное ядро; 21 - конечная полоска; 22 - ядра подбугорной области; 23 - хвостатое ядро; 24 - кора полушария; 25 - полость прозрачной перегородки; 26 - передний рог бокового желудочка; АА - внутренняя капсула; а - лобно-бугорный путь; б - лобно-мостовой путь; в - корково-ядерный путь; г - корково - спинномозговой путь; д - луковично-бугорный путь и спинно-бугор-ный путь; е - затылочно-височно-мостовой путь; ж - центрально-слуховой путь; з - центральный зрительный тракт
Передняя ножка внутренней капсулы находится между хвостатым и чечевицеобразным ядрами. Она образована волокнами, идущими от коры лобной доли к зрительному бугру, tractus frontothalamicus, к красному ядру, tractus frontorubralis, к ядрам моста, tractus frontopontine. Кроме этих волокон, которые имеют продольное направление, здесь проходят поперечные волокна от хвостатого к чечевицеобразному ядру.
Задняя ножка внутренней капсулы длиннее передней. Она расположена между зрительным бугром и чечевицеобразным ядром. Передние две трети задней ножки образуют волокна от клеток коры прецентральной извилины к двигательным ядрам спинного мозга, которые составляют корково-спинномозговой путь, tractus cortico spinalis. Задняя треть этой ножки образована чувствительными волокнами - отростками клеток ядер зрительного бугра к коре зацентральной извилины - бугорно-корковые пучки, fasciculi thalamocorticates. Здесь же проходят волокна от коры к зрительному бугру, которые составляют корково-бугорные пучки, fasciculi соrticothalamici.
Колено внутренней капсулы занимает область угла, образованного передней и задней ножками. Его образуют волокна от нижних отделов коры прецентральной извилины к ядрам черепных нервов среднего мозга, моста и продолговатого мозга, fibrae corticonucleares. Кзади задняя ножка внутренней сумки продолжается в ее зачечевицеобразную часть, pars retrolentiformis capsulae internae, которую образуют пучки волокон от первичных зрительных центров к коре затылочной доли; книзу от чечевицеобразного ядра в подчечевицеобразной части внутренней сумки, pars sublenti-formis capsulae internae, расположены пучки волокон от внутреннего коленчатого тела к верхней височной извилине.
Проводящие пути головного и спинного мозга
Проводящие пути головного и спинного мозга делят на: 1) ассоциационные, 2) комиссуральные, 3) проекционные. Ассоциационные и комиссуральные пути были рассмотрены при описании различных отделов центральной нервной системы. Проекционные пути подразделяются на чувствительные - афферентные и двигательные - эфферентные. Наиболее важные из них следующие.
Афферентные проводящие пути
1. Проводящие пути тактильной, болевой и температурной чувствительности (так называемая общая чувствительность) образованы цепью из нейроцитов. Первыми нейроцитами этих путей являются псевдоуниполярные клетки спинномозговых узлов. Дендриты этих клеток в составе периферических нервов идут в кожу, где заканчиваются экстерорецепторами, воспринимающими раздражения общей чувствительности. Нейриты псевдоуниполярных клеток в составе задних корешков вступают в корешковую зону спинного мозга и заканчиваются в клетках заднего рога. Вторыми нейроцитами этого пути являются клетки задних рогов, нейриты которых через переднюю белую спайку переходят на противоположную сторону и образуют передний и боковой спино-бугорные тракты. Эти пути проходят продолговатый мозг и вступают в покрышку моста, в верхнем отделе которого соединяются с наружным отделом медиальной петли. Вместе с медиальной петлей они проходят кнаружи от красных ядер в покрышке среднего мозга и вступают в зрительный бугор, клетки которого являются третьими нейроцитами этого пути. Их нейриты образуют бугорно-корковые пучки, fasciculi thalamocorticales, которые заканчиваются в ядрах кожного анализатора (зацентральная извилина, верхняя теменная долька).
2. Проводящие пути тактильной, болевой и температурной чувствительности от кожи головы, твердой мозговой оболочки, склеры, роговицы и конъюнктивы глаза, слизистой оболочки рта, носа и его придаточных полостей, костей черепа, нижней челюсти и зубов образуют цепи из 3 нейроцитов. Тела первых нейроцитов этих путей расположены в тройничном узле. Дендриты псевдоуниполярных клеток этого узла заканчиваются соответствующими рецепторами в перечисленных образованиях. Нейриты клеток образуют чувствительный корешок тройничного нерва, который вступает в толщу моста и делится там на восходящую и нисходящую ветви.
Восходящую ветвь образуют довольно толстые волокна тактильной и проприоцептивной чувствительности, а тонкие волокна нисходящей ветви проводят к ядру спинномозгового пути тройничного нерва импульсы болевой и температурной чувствительности. Восходящие волокна тактильной чувствительности заканчиваются в верхнем чувствительном ядре тройничного нерва. От клеток этого ядра возникают волокна вторых нейроцитов данного пути. Эти волокна образуют перекрест, в области моста и затем входят в состав медиальной петли, вместе с которой проходят в покрышке ножки мозга и заканчиваются в зрительном бугре. Нейриты клеток зрительного бугра в составе волокон задней трети задней ножки внутренней капсулы направляют к коре нижнего участка зацентральной извилины, где и заканчиваются. Нисходящие волокна чувствительного корешка внутри моста образуют крупный по объему спинномозговой путь тройничного нерва, в составе которого имеется большое количество тонких безмиелиновых волокон. Этот путь проходит до IV шейного сегмента, постепенно заканчиваясь в клетках одноименного ядра, причем волокна от самых наружных отделов кожи лица, какой бы из трех ветвей этого нерва они ни принадлежали, заканчиваются в самых нижних отделах ядра, а от более медиальных - в верхнем его отделе. Вторыми нейроцитами пути болевой и температурной чувствительности являются клетки ядра спинномозгового пути тройничного нерва. Их нейриты совершают перекрест, проходят в ретикулярной формации продолговатого мозга и моста и вступают в состав медиальной петли, вместе с которой заканчиваются в зрительном бугре. Ход и место окончания третьего нейроцита этого пути такой же, как пути тактильной чувствительности.
3. Проводящие пути проприоцептивной (глубокой) чувствительности передают в область коркового ядра двигательного анализатора импульсы глубокой чувствительности от проприоцепторов мышц, сухожилий, суставов, которые служат сигналами изменений напряжения мышц, суставных сумок и других элементов двигательного аппарата, т. е. сигналами кинестических раздражений. Первыми чувствительными нейроцитами этих путей являются клетки спинномозговых узлов. Дендриты псевдоуниполярных клеток спинномозговых узлов заканчиваются проприоцепторами в мышцах, сухожилиях, суставах, а нейриты в составе задних корешков вступают в корешковую зону задних рогов спинного мозга и, не прерываясь в его ядрах, переходят в задние канатики. В составе задних канатиков проходит также часть волокон тактильной чувствительности. Волокна проприоцептивной чувствительности из нижних сегментов спинного мозга составляют медиально расположенный в заднем канатике тонкий пучок, а соответствующие волокна 12 верхних сегментов образуют клиновидный пучок, лежащий латерально от тонкого пучка. Тонкий и клиновидный пучки заканчиваются в одноименных переключательных ядрах продолговатого мозга. Вторыми нейроцитами являются клетки переключательных ядер тонкого и клиновидного канатиков, волокна которых изгибаются, обходят спереди центральный канал и в межоливарном слое образуют перекрест с волокнами противоположной стороны. Эти волокна называют внутренними дугообразными, а их перекрест - верхним чувствительным перекрестом, или перекрестом петли, так как они составляют медиальную петлю. Медиальная петля в области моста расположена позади от пирамидного пути. Уже в верхних отделах продолговатого мозга к ней начинают присоединяться пути тактильной, болевой и температурной чувствительности - спинно-бугорные тракты. Соединение медиальной петли и спинно-бугорных трактов заканчивается в верхних отделах моста, в результате чего здесь имеется крупный пучок нервных волокон, которые являются проводниками всех видов чувствительности. В этом пучке волокна проприоцептивной чувствительности (мышечно-суставное чувство) занимают наиболее медиальное положение. Кнаружи от них находится путь тактильной чувствительности, латеральнее от него - температурной, снаружи от которого располагается путь болевой чувствительности.
Из моста медиальная петля переходит в покрышку среднего мозга, располагаясь кнаружи от красных ядер и затем вступая в зрительный бугор, где заканчивается в одном из ядер вентральной группы. Отростки клеток этого ядра являются третьими нейроцитами проводящих путей проприоцептивной чувствительности, которые образуют бугорно-корковые пучки. Эти пучки заканчиваются в ядрах кожного и двигательного анализаторов (см. рис. 207).
Считают, что проприоцептивные пути от двигательного аппарата головы проходят в составе проводящих путей тройничного, лицевого, языко-глоточного, блуждающего, подъязычного и добавочного черепных нервов. Эти пути также составляют цепи из 3 нейроцитов и заканчиваются в ядрах кожного и двигательного анализаторов. В связи с тем, что рассмотренный путь проприоцептивной чувствительности заканчивается в коре, где осуществляется высшая форма анализа и синтеза поступивших кинестических раздражений. Эти раздражения становятся ощущением и осознаются, т. е. человек получает представление о положении частей своего тела в пространстве и изменениях этого положения. Кроме рассмотренного пути проприоцептивной чувствительности, существуют проприоцептивные пути, нервные импульсы по которым не доходят до коры полушарий головного мозга и, следовательно, не становятся ощущениями. К ним относятся проприоцептивные пути к мозжечку.
4. Проприоцептивные пути к мозжечку участвуют в безусловнорефлекторной регуляции мышечного тонуса, равновесия, координации и синергии движений. Различают передний и задний спинно-мозжечковые пути, первыми нейроцитами которых являются псевдоуниполярные клетки спинно-мозговых узлов. Дендриты этих клеток заканчиваются проприоцепторами в мышцах, сухожилиях, суставах и постоянно воспринимают раздражения, сигнализирующие о состоянии двигательного аппарата. Нейриты этих клеток в составе спинных корешков вступают в спинной мозг и заканчиваются в грудном и промежуточно-медиальном ядрах заднего рога.
Нейриты клеток грудного ядра выходят в боковой канатик спинного мозга той же стороны и образуют задний спинно-мозжечковый путь, который в составе нижних мозжечковых ножек вступает в мозжечок и заканчивается в коре червя. Нейриты клеток промежуточно-медиального ядра как своей, так и противоположной сторон в боковом канатике спинного мозга образуют передний спинно-мозжечковый путь. Волокна этого пути проходят продолговатый мозг и мост; возле крыши среднего мозга неперекрещенные в спинном мозге волокна образуют перекрест с волокнами противоположной стороны, после чего передний спинно-мозжечковый в составе верхних мозжечковых ножек проходит в червь мозжечка и заканчивается на клетках его коры. Оба проприоцептивных пути к мозжечку несут к нему нервные импульсы со своей стороны.
5. Обонятельные проводящие пути подразделяют на периферический и центральный отделы. К первому относят систему центральных отростков обонятельных клеток, которые через отверстия решетчатой пластинки решетчатой кости проникают из полости носа в полость черепа и заканчиваются на митральных клетках обонятельной луковицы. Обонятельные луковицы, а также ряд подкорковых и корковых образований составляют центральный отдел обонятельного проводящего пути. Нейриты митральных клеток, соединившись, образуют обонятельный тракт. Часть нейритов митральных клеток заканчивается на зернистых клетках обонятельной луковицы, нейриты которых образуют синапсы с митральными клетками. Считают, что такие связи между клетками в пределах обонятельной луковицы способствуют усилению обонятельных импульсов. В области обонятельного треугольника некоторые волокна обонятельного тракта заканчиваются. Большая же их часть разделяется на три пучка, из которых наружный заканчивается в коре крючка парагиппокампальной извилины, средний - в переднем продырявленном веществе, а медиальный - проходит в околообонятельное поле коры лобной доли мозга. Обонятельные центры коры соединены проекционными волокнами с подкорковыми обонятельными центрами, из которых сосковидные тела в свою очередь соединены с многими ядрами мозгового ствола (сосково-бугорный и сосково-покрышечный пучки).
6. Проводящие пути от органов растительной жизни - внутренних органов, кровеносных и лимфатических сосудов и др. образуют цепи из 3 нейроцитов. Первыми нейроцитами этих путей являются клетки узлов некоторых черепных нервов (тройничного, лицевого, языко-глоточного и блуждающего) и спинномозговых узлов, вторые нейроциты - отростки клеток ядер указанных нервов, а третьи - отростки клеток зрительного бугра, которые заканчиваются в премоторной зоне и лимбической области коры.
Значительное место в чувствительной иннервации органов растительной жизни принадлежит блуждающему нерву. Клетки первого нейроцита чувствительного пути расположены в верхнем и нижнем его узлах. Дендриты этих клеток заканчиваются интерорецепторами во внутренних органах, а нейриты в составе корешков нерва вступают в толщу продолговатого мозга, где заканчиваются в ядре одиночного пучка. Отростки клеток этого ядра являются вторыми нейроцитами. Они образуют перекрестное, после чего в области моста присоединяются к медиальной петле, вместе с которой проходят в зрительный бугор. Клетки последнего - третьи нейроциты этого пути - заканчиваются в нижнем отделе зацентральной извилины.
Эфферентные проводящие пути
1. Пирамидный путь, tractus pyramidalis, осуществляет сознательную, произвольную регуляцию движений. Его составляет цепочка из 2 нейроцитов, среди которых различают центральный и периферический. Центральными нейроцитами называют клетки коркового ядра двигательного анализатора (см. рис. 207), которые заканчиваются в двигательных ядрах черепных нервов (fibrae corticonucleares) и двигательных ядрах передних рогов спинного мозга (tractus corticospinalis). Периферическими нейроцитами являются клетки двигательных ядер черепных нервов и двигательных ядер спинного мозга, нейриты которых в составе периферических нервов заканчиваются в соответствующих мышцах двигательными окончаниями. Пирамидный путь проводит импульсы от коркового ядра к активной части двигательного аппарата - мышцам. Эти импульсы возникают в коре как результат анализа и синтеза раздражений, поступивших в нее по проприоцептивному пути, а также по другим путям (зрительным, вестибулярным, тактильным).
Основную массу волокон, которые являются нейритами нейроцитов пирамидного пути, составляют отростки пирамидных клеток прецентральной и зацентральной извилин. Кроме них, в состав этого пути входят волокна от задней трети верхней и средней лобных извилин, передней трети верхней теменной дольки и переднего отдела надкраевой извилины. От клеток моторной зоны коры возникает основная масса толстых волокон, тогда как из остальных отделов идут более тонкие волокна этого пути. Все эти волокна образуют компактный пучок, который заполняет собой пространство между зрительным бугром и чечевицеобразным ядром, образуя колено и две трети заднего бедра внутренней капсулы. Колено внутренней капсулы составляют волокна от нижней части прецентральной и зацентральной извилин, которые заканчиваются в двигательных ядрах черепных нервов,- корково-ядерные волокна. Остальные волокна образуют переднюю и среднюю трети задней ножки внутренней капсулы и переходят в средний мозг, где от них отделяются пучки волокон к ядрам глазодвигательного и блокового нервов, а также к ядрам отводящего и добавочного нервов. Из среднего мозга двигательный путь следует в вентральную часть моста, где его волокнами делится на несколько пучков. В средних отделах моста от двигательного пути отделяется пучок волокон к двигательному ядру тройничного нерва своей и противоположной сторон. От клеток этих ядер возникают двигательные волокна тройничного нерва, которые в составе его нижнечелюстного нерва идут к жевательной мускулатуре и заканчиваются в ней двигательными окончаниями.
В нижнем отделе моста от двигательного пути отделяются волокна к двигательным ядрам лицевого нерва своей и противоположной сторон, после чего двигательный путь переходит в продолговатый мозг, где снова принимает вид компактного пучка. В верхнем отделе продолговатого мозга от двигательного пути отходят волокна к двигательным ядрам блуждающего, языко-глоточного и подъязычного нервов. После отхождения этих пучков двигательный путь состоит только из волокон, соединяющих кору с ядрами передних рогов спинного мозга. Большая часть этих волокон совершает перекрест в нижнем отделе продолговатого мозга, decussatio pyramidum, и переходит в боковой канатик спинного мозга, где образует, tractus corticospinali lateralis, волокна которого заканчиваются в двигательных ядрах передних рогов спинного мозга. Меньшая часть волокон в области продолговатого мозга перекреста не образует и в спинном мозге составляет tractus corticospinalis anterior; часть его волокон заканчивается в двигательных ядрах спинного мозга своей стороны, а другая часть через переднюю белую спайку, совершает перекрест и заканчивается в таких же ядрах противоположной стороны. Передний пирамидный пучок прослеживается только в шейных и верхних грудных сегментах спинного мозга.
Таким образом, пирамидный путь осуществляет преимущественно перекрестную иннервацию мускулатуры тела, и при одностороннем поражении центральных-нейроцитов наступает паралич мускулатуры на стороне, противоположной очагу поражения. Такой односторонней иннервацией обладают не все мышечные группы. Большая часть мышц, а именно: мышцы глазного яблока, жевательные, мимические, мышцы верхней части лица, глотки, гортани, шеи, туловища и промежности, получают так называемую двустороннюю иннервацию, т. е. от центральных нейронов как своей, так и противоположной стороны, что обеспечивает в известной степени сохранение иннервации этих мышц при одностороннем поражении центральных нейронов. Односторонней иннервацией обладают мышцы конечностей, языка и мимические мышцы нижней половины лица, в связи с чем при одностороннем поражении центральных нейронов этих мышц наступает полная утрата способности к движению, т. е. паралич.
2. Проводящие пути экстрапирамидной системы проходят к передним рогам спинного мозга вне пирамидных путей, через покрышку и сетчатое образование мозгового ствола. К экстрапирамидной системе относят следующие образования: хвостатое и чечевицеобразное ядра, зрительный бугор, черное вещество, красное ядро, образования подбугорной области, мозжечок, оливу продолговатого мозга и ядра ретикулярной формации мозгового ствола. В связи с большим количеством образований в составе экстрапирамидной системы ее проводящая система сложна. По сравнению с пирамидной системой экстрапирамидная система является филогенетически более старым образованием. Функция ее заключается в осуществлении сложных безусловнорефлекторных двигательных актов, основанных на опыте вида, в то время как двигательная корковая деятельность основана на условных рефлексах, закрепляющих индивидуальный опыт. Экстра-пирамидная система осуществляет автоматическую, без участия сознания, регуляцию и координацию степени напряжения (тонуса) мышечной системы, сокращение или расслабление определенных мышечных групп (сложные синергии), обеспечивает готовность двигательного аппарата к выполнению сложных, точных, целенаправленных действий. Поражение экстрапирамидной системы проявляется главным образом изменениями мышечного тонуса и двигательной активности.
Нервные импульсы поступают в образования экстрапирамидной системы от коры полушарий головного мозга, от зрительных бугров, ядер подбугорной области и мозжечка - важнейшего центра координации как произвольных, так и непроизвольных сложных двигательных актов. По переднему и заднему спинно-мозжечковым путям в мозжечок поступают: импульсы, сигнализирующие о состоянии органов движения, часть волокон от ядер задних канатиков, от оливы продолговатого мозга, которая выполняет функцию промежуточного центра равновесия, и от преддверных ядер. Волокна от вестибулярных ядер в основном заканчиваются в шаровидном ядре, ядре шатра и в коре клочка. Лишь небольшая часть их заканчивается в зубчатом ядре.
От коры различных отделов полушарий головного мозга, главным образом от лобной и височной долей, возникают двухнейронные пути к мозжечку, по которым осуществляются контроль и регуляция его деятельности. Первыми нейроцитами этих путей являются клетки коры этих отделов, нейриты которых составляют лобно-мостовой и височно-мостовой пучки; из них первый проходит в составе волокон передней ножки внутренней капсулы, а второй - задней ножки. Оба пучка заканчиваются в собственных ядрах моста своей стороны. Клетки собственных ядер моста являются вторыми нейроцитами путей к мозжечку от коры полушария мозга. Их нейриты в толще моста совершают перекрест и в составе средних мозжечковых ножек вступают в кору мозжечка.
Нейриты клеток ядра шатра, шаровидного и зубчатого ядер образуют мозжечково-преддверный пучок, который проходит в составе нижней мозжечковой ножки и заканчивается в вестибулярных ядрах мозгового ствола. Нейриты клеток преддверных ядер образуют преддверно-спинномозговой путь, волокна которого заканчиваются на двигательных клетках передних рогов спинного мозга, а также на двигательных ядрах черепных нервов продолговатого мозга, моста и среднего мозга.
Основную массу волокон верхней мозжечковой ножки составляют нейриты клеток зубчатого ядра. Этот мощный пучок волокон в области покрышки ножек мозга совершает перекрест, после чего делится на восходящую и нисходящую ветви. Нисходящая ветвь заканчивается в ядрах ретикулярной формации покрышки, восходящая - вступает в красное ядро, где часть волокон заканчивается, а остальные проходят через него и вступают в латеральное ядро зрительного бугра, nucleus lateralis thalami.
Это ядро зрительного бугра связано с корой передней центральной извилины полушарий головного мозга.
Кроме волокон от мозжечка, красное ядро получает волокна от хвостатого ядра и скорлупы, которые образуют пучок, называемый лентикулярной петлей, ansa lenticularis. Волокна от клеток бледного шара идут в красное ядро, черное вещество, ядра зрительного бугра, ядра подбугорной области. Черное вещество соединено с красным ядром, стриарной системой и корой центральных извилин.
Типовые, возрастные и половые особенности строения центральной нервной системы
Центральная нервная система и особенно головной мозг у разных людей отличаются по своему строению. Эти отличия касаются формы мозга и его отдельных образований, величины, веса и других показателей. Установлено, что между формой головного мозга и формой черепа наблюдается определенное соответствие. Это обстоятельство имеет важное практическое значение в связи с возможностью проекции на наружные покровы головы различных ядер, борозд и извилин.
У одних людей лобный отдел полушарий развит больше, чем затылочный и теменной, а у других, наоборот, теменной и затылочный отделы преобладают над лобным. Индивидуально различны по размерам ядерные образования, борозды и извилины, площадь коры полушарий, а также цитоархитектонических полей.
Наблюдаются большие различия в весе мозга - от 1000 до 2285 г. Они не связаны с умственным развитием человека, так как у выдающихся людей вес мозга был большим или малым. Например, мозг И. С. Тургенева весил 2021 г, а мозг Анатоля Франса - 1017 г.
Вес мозга к 8-му месяцу жизни удваивается по сравнению с новорожденными, у которых мозг весит в среднем 370 г. К периоду полового созревания этот вес увеличивается в 4 раза. К 18-40 годам жизни у мужчин и к 16-18 годам у женщин вес мозга становится устойчивым, а к старости он постепенно снижается. Вес головного мозга у мужчин несколько больше, чем у женщин.
Оболочки головного мозга
Твердая оболочка головного мозга, dura mater encephali, прилегает к внутренней поверхности костей черепа и плотно сращена с ней в области основания и швов. Рыхлое соединение dura mater с костями свода черепа послужило основанием для выделения очень узкого эпидурального пространства, которое пронизано соединительнотканными волокнами, сосудами и нервами. К гладкой внутренней поверхности твердой оболочки прилегает паутинная оболочка головного мозга. Расположенную между ними узкую щель называют субдуралъным пространством.
В некоторых местах твердая оболочка образует отроги, которые проникают между различными отделами головного мозга. Так, в продольной щели полушарий расположен серповидной формы отросток - серп большого мозга, falx cerebri. Верхний утолщенный край серпа большого мозга, образованный двумя расходящимися пластинками твердой оболочки, примыкает к сагиттальной борозде черепа от петушьего гребня решетчатой кости до внутреннего затылочного выступа (рис. 213).
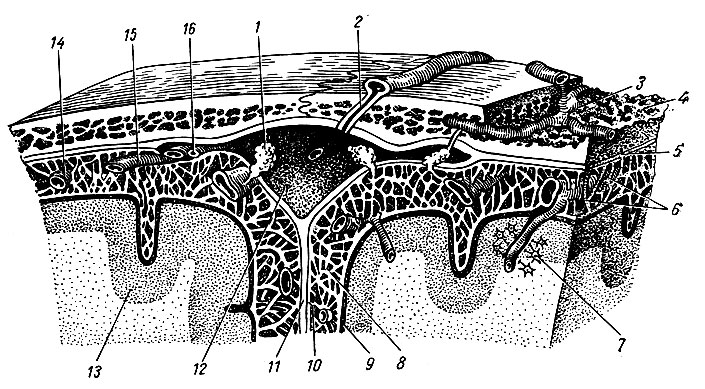
Рис. 213. Оболочки головного мозга. 1 - грануляции паутинной оболочки; 2 - вена-выпускник; 3 - вена губчатого вещества кости; 4 - губчатое вещество кости; 5 - твердая оболочка головного мозга; 6 - перекладины паутинной оболочки; 7 - околососудистое пространство; 8 - подпаутинное пространство; 9 - сосудистая оболочка; 10 - паутинная оболочка; 11 - серп большого мозга; 12 - верхний сагиттальный синус; 13 - кора мозга; 14 - ветвь артерии мозга; 15, 16 - вены мозга
Намет мозжечка, tentorium cerebelli, представляет собой широкую складку твердой оболочки, которая в виде двускатной крыши располагается в поперечной борозде мозга, отделяя содержимое задней черепной ямки от затылочных долей. Задне-латеральный утолщенный край мозжечкового намета сращен с бороздой поперечного синуса и верхней гранью пирамиды височной кости. Свободный край мозжечкового намета округлой формы. Он натянут между задним концом свободного края мозгового серпа и задним наклонным отростком клиновидной кости. Свободный край ограничивает вырезку намета мозжечка, incisura tentorii, которая сообщает заднюю и среднюю ямки черепа. В области этого отверстия расположены ножки мозга. В задней ямке черепа, между полушариями мозжечка, расположен серп мозжечка, falx cerebelli.
В области турецкого седла твердая оболочка образует диафрагму турецкого седла, diaphragma sellae, узкое отверстие которой пропускает воронку к гипофизу. На верхушке пирамиды височной кости отростки твердой оболочки образуют тройничную полость, cavum trigemina lis, где расположен тройничный узел тройничного нерва. В толще твердой оболочки находятся артерии, вены, нервные волокна и их окончания, а также особые каналы - венозные синусы, sinus durae matris.
Венозные синусы представляют собой пространства между расщепившимися листками твердой оболочки. Большая часть этих каналов расположена пристеночно. В венозные синусы поступает кровь из вен головного мозга, а затем выводится по ним в экстракраниальные венозные образования, главным образом во внутренние яремные вены.
Паутинная оболочка головного мозга, arachnoidea encephali, в виде очень тонкого, прозрачного бессосудистого листка покрывает головной мозг и книзу переходит в паутинную оболочку спинного мозга. Между твердой и паутинной оболочками имеется узкое субдуральное пространство, которое пронизано соединительнотканными волокнами, непрочно соединяющими эти листки. Прочные сращения между твердой и паутинной оболочками образуются посредством особых структур, которые называют грануляциями паутинной оболочки, а также на поверхности мозговых вен, при их вступлении в венозные синусы.
Паутинная и сосудистая оболочки ограничивают заполненное спинномозговой жидкостью подпаутинное пространство, cavum subarachnoidal. Оно пронизано многочисленными соединительнотканными волокнами, которые соединяют эти оболочки и осуществляют фиксацию расположенных здесь кровеносных сосудов. Иные соотношения этих оболочек имеются в области различных углублений мозга. Если сосудистая оболочка покрывает мозговую поверхность в глубине борозд, щелей и ямок, то паутинная - переходит с одной возвышенности на другую, образуя различной величины субарахноидальные вместилища. Самые крупные из этих вместилищ получили название цистерн. Наиболее значительными из этих цистерн являются следующие (рис. 214):
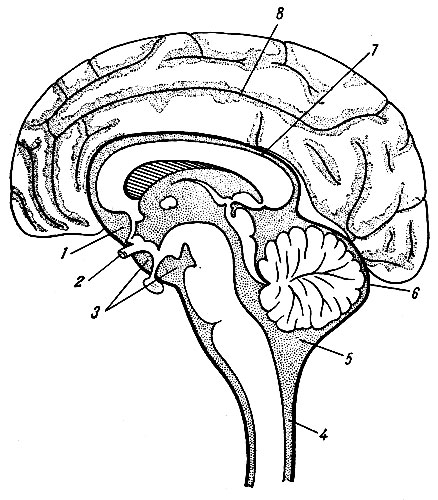
Рис. 214. Субарахноидальные цистерны. 1 - цистерна зрительного перекреста; 2 - зрительный перекрест; 3 - межножковая цистерна; 4 - подпаутинное пространство спинного мозга; 5 - мозжеч-ково-продолговатомозговая цистерна; 6 - паутинная оболочка; 7 - подпаутинное пространство над мозолистым телом; 8 - подпаутинное пространство в бороздах
1) cisterna cerebellomedullaris - находится между мозжечком и дорсальной поверхностью продолговатого мозга;
2) cisterna pontis lateralis - определяется в области мосто-мозжечкового угла;
3) cisterna interpeduncularis - расположена между ножками мозга;
4) cisterna chiasmatis - находится впереди зрительного перекрестка;
5) cisterna fossa lateralis cerebri - соответствует одноименной ямке и борозде.
Субарахноидальное пространство головного и спинного мозга является единым. Образующаяся в желудочках головного мозга спинномозговая жидкость поступает в это пространство через отверстия IV желудочка, а отводится в венозную систему посредством грануляций паутинной оболочки.
Сосудистая оболочка головного мозга, pia mater encephali, плотно прилегает к мозговому веществу, как на свободной поверхности мозга, так и в глубине борозд. Сосудистые сплетения желудочков осуществляют секрецию спинномозговой жидкости. В толще сосудистой оболочки расположены сети кровеносных сосудов.
При использовании материалов сайта активная ссылка обязательна:
http://anfiz.ru/ 'Анатомия и физиология человека'