Головной мозг (encephalon, s. cerebrum)
Во всей живой природе нет органа более сложного и совершенного, чем головной мозг человека. Именно головному мозгу человек обязан своим высоким положением, своей властью над живой и мертвой природой. Головной мозг регулирует все физиологические процессы организма и все реакции человека, вырабатывает все новые и более совершенные их формы. Человеческий мозг по своим функциям и структуре резко выделяется по сравнению с мозгом всех других животных и, обладая наиболее сложной организацией, является наиболее совершенно действующим аппаратом высшей нервной деятельности. Он представляет собой высокоорганизованное целое, построенное из миллиардов в строгом порядке размещенных и работающих нервных клеток и волокон.
Для суждения о степени развития мозга у человека могут служить следующие данные. Человеческий мозг весит у новорожденного около 350 г, составляя 1/8 веса тела, к концу первого месяца - около 450 г, а к концу года вес его почти удваивается, достигая 700-800 г. К 6 годам вес головного мозга примерно втрое больше веса мозга новорожденного, а к 10 годам уже достигает 1300 г; к 20 годам средний вес головного мозга достигает уже почти своего предела - в среднем 1375 г (740 общего веса). После 60 лет вес мозга постепенно и медленно начинает убывать.
По абсолютной величине мозг человека - не самый большой мозг в животном мире. Например, вес мозга у индийского слона равен 5200 г, у кита доходит до 7000 г, у дельфинов - 3000 г, но если принять во внимание огромные размеры этих животных, у которых вес тела измеряется не килограммами, а тоннами, то относительный вес мозга и у них будет меньше человеческого мозга. По отношению же к весу всего тела вес мозга у человека далеко превосходит мозг других животных. Так, например, отношение веса всего тела к весу головного мозга выражается следующим образом: у рыб вес тела в 5668 раз больше веса мозга, у пресмыкающихся - в 1321 раз, у птиц - в 212 раз, у кита - в 1200 раз, у человекообразной обезьяны - в 213 раз, у человека - только в 45 раз. Но следует отметить, что у некоторых обезьян и певчих птиц, а также у мышей относительный вес мозга больше, чем у человека.
На рис. 274 изображен головной мозг ряда позвоночных животных и человека. Нетрудно убедиться, что головной мозг по своему развитию, в особенности больших полушарий переднего мозга, занимает у человека первое место.
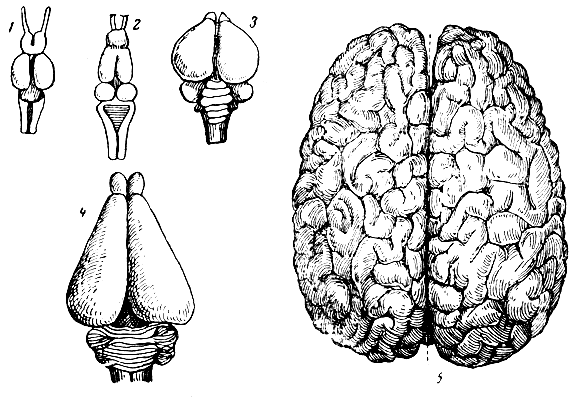
Рис. 274. Сравнительная форма головного мозга. 1 - карп; 2 - лягушка; 3 - голубь; 4 - кролик; 5 - человек
Поверхность полушарий головного мозга у человека значительно увеличивается многочисленными складками-извилинами, отделенными друг от друга глубокими бороздами. Складчатость увеличивает поверхность мозга у человека в 8-10 раз. Так, площадь коры человеческого мозга в среднем равна 2250 см2, а у лошади - только 350 см2. У низших же животных, например у рыб, лягушек и даже у низших млекопитающих (грызуны), борозды и извилины, увеличивающие поверхность мозга, совершенно отсутствуют.
Рельеф мозговой поверхности человека при ближайшем его изучении поражает своим разнообразием. Нет таких двух человек, чтобы у них полностью совпадала картина извилин и борозд, покрывающих полушария. Даже полушария одного и того же человека далеко не идентичны, т. е. борозды и извилины правого и левого полушарий мозга не вполне симметричны. Однако, несмотря на бесконечную изменчивость типов борозд и извилин в деталях, наукой установлена общая схема, характерная для мозга современного человека.
Сильное развитие головного мозга человека, безусловно, говорит о его высоком умственном развитии. Но по одной только величине и строению мозга еще нельзя судить об умственных способностях человека. Мнение, что мозг с сильным развитием борозд принадлежит людям, обладающим более высоким интеллектом, остается недоказанным. Точно так же это относится и к животным. Лошадь, собака обладают небольшой поверхностью мозговой коры и слабым развитием извилин, в то время как овца и осел имеют мозг с очень большим количеством борозд и извилин.
Таким образом, надо полагать, что ни по форме и ни по величине мозга нельзя еще судить об умственных способностях; не только абсолютный, но и относительный вес мозга также не служит надежным показателем степени умственного развития данного вида животного, а тем более человека. Из истории человечества нам известно немало выдающихся по своим способностям и талантам людей как с очень большим, так и с очень малым весом мозга. Мозг писателя И. С. Тургенева весил 2012 г, Байрона - 2238 г, Шиллера - 1871 г, но мозг такого же веса встречался и у людей, в умственном отношении ничем не выдающихся. С другой стороны, весьма одаренные в умственном отношении люди имели мозг совсем небольшого веса: химик Либих - 1325 г, поэт Уитмен - 1282 г, а писатель Анатоль Франс - даже только 1017 г. Из этих даже немногих примеров ясно видно, что умственные способности или одаренность человека не стоит в прямой зависимости от объема и веса мозга. Таким образом, надо признать, что изменения интеллектуальной способности, по-видимому, связаны не только с весом мозга, с характером строения его коры, обширностью ее поверхности, а самое главное - с функциональными свойствами нервных клеток мозгового вещества, с их физиологической активностью.
Тем более следует подчеркнуть тот факт, что степень развитая мозга не связана ни с расовыми особенностями, ни с социальным положением.
Головной мозг, как уже упоминалось ранее, принято делить на стволовую часть и плащ. Принимая во внимание развитие мозга, удобнее начать его изучение со стволовой части.
Мозговой ствол (рис. 275, 276, 277). Хотя мозговой ствол значительно сложнее спинного мозга, но в своем строении все же сохраняет с ним сходство. Спинной мозг, войдя в полость черепа через большое затылочное отверстие, непосредственно продолжается в ствол головного мозга. По внешнему виду мозговой ствол напоминает спинной мозг. Снаружи он также состоит из белого вещества, а серое находится внутри; из ствола выходят двигательные и в ствол из черепномозговых (головных) узлов входят чувствительные нервы. Отличие ствола заключается в более сложных взаимоотношениях серого и белого вещества и в отсутствии правильности входа и выхода черепномозговых нервов. Кроме того, со стволом теснейшим образом связан мозжечок, в котором, как и в больших полушариях мозга, главная масса серого вещества расположена снаружи.
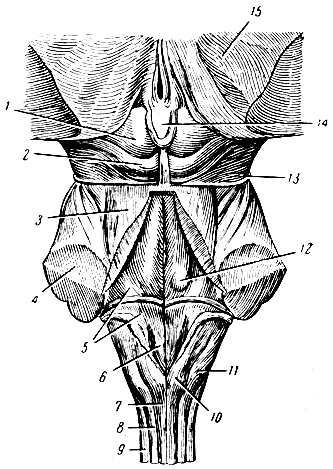
Рис. 275. Стволовая часть мозга сзади и сверху. 1 - верхний бугорок четверохолмия; 2 - нижний бугорок четверохолмия; 3 - ножка мозжечка к четверохолмию; 4 - ножка мозжечка к мосту; 5 - слуховое поле (место расположения ядер слухового нерва); 6 - треугольник подъязычного нерва (место расположения ядра подъязычного нерва); 7 - нежный пучок; 8 - клиновидный пучок; 9 - боковая часть продолговатого мозга (веревчатое тело); 10 - узел нежного пучка; 11 - узел клиновидного пучка; 12 - лицевой бугорок; 13 - блоковый нерв; 14 - шишковидная железа; 15 - зрительный бугор
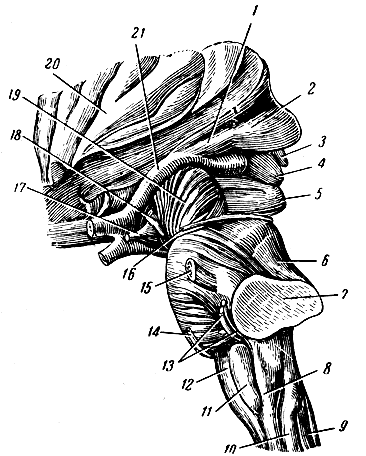
Рис. 276. Стволовая часть мозга сбоку. 1 - латеральное коленчатое тело; 2 - подушка (задняя часть зрительного бугра); 3 - шишковидная железа; 4-5 - четверохолмие; 6 - ножка мозжечка к четверохолмию; 7 - ножка мозжечка к мосту; 8 - боковой пучок продолговатого мозга (веревчатое тело); 9 - нежный пучок; 10 - клиновидный пучок; 11 - олива; 12 - пирамида; 13 - слуховой и лицевой нервы; 14 - отводящий нерв; 15 - тройничный нерв; 16 - блоковый нерв; 17 - воронка (дно III желудочка); 18 - глазодвигательный нерв; 19 - ножка мозга; 20 - островок Рейля; 21 - зрительный тракт
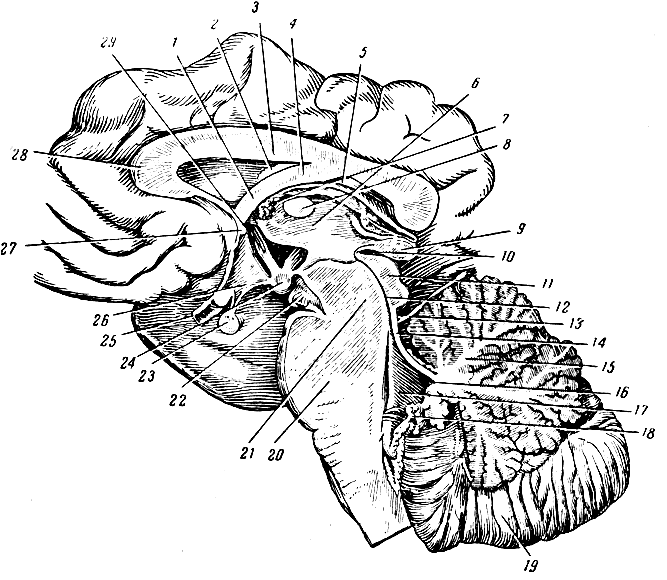
Рис. 277. Срединный сагиттальный разрез стволовой части мозга. 1 - передняя ножка свода; 2 - сосудистое сплетение бокового желудочка; 3 - мозолистое тело; 4 - тело свода; 5 - задняя ножка свода; 6 - правая боковая стенка III желудочка (медиальная поверхность зрительного бугра); 7 - монроево отверстие (из III желудочка в боковой); 8 - серая комиссура; 9 - шишковидная железа; 10 - задняя комиссура; 11 - чехтверохолмие; 12 - сильвиев водопровод; 13 - блоковый нерв; 14 - передний мозговой парус; 15 - червячок мозжечка в разрезе; 16 - верхушка IV желудочка; 17 - IV желудочек; 18 - сосудистое сплетение IV желудочка и задний мозговой парус; 19 - правое полушарие мозжечка; 20 - варолиев мост; 21 - ножка мозга; 22 - глазодвигательный нерв; 23 - гипофиз; 24 - сосковое тело; 25 - зрительный нерв; 26 - воронка; 27 - передняя белая комиссура; 28 - колено мозолистого тела; 29 - скрытая часть передней ножки свода
В состав ствола входят: продолговатый мозг, мозжечок, мозговой мост, четверохолмие, ножки мозга и зрительные бугры. Сверху ствол покрыт массой больших полушарий (плащом).
Продолговатый мозг (medulla oblongata)* представляет собой утолщенное и расширенное продолжение спинного мозга. Войдя в полость черепа, он поворачивает кпереди под тупым углом и ложится на скат затылочной кости. На передненижней поверхности продолговатого мозга проходит передняя продольная борозда, которая является продолжением одноименной борозды спинного мозга. По бокам от борозды лежат два продольных тяжа-пучка нервных волокон, это так называемые пирамиды, состоящие из скоплений двигательных (центробежных) нервных волокон, спускающихся от коры больших полушарий; по пути к ним присоединяются центробежные волокна, начинающиеся в сером веществе мозгового ствола. Вверх пирамидные пучки проходят через мозговой мост; внизу большая часть пучков на границе со спинным мозгом образует перекрест (decussatio pyramidum; волокна правой половины переходят на левую, а левой - на правую сторону. Перекрещенные волокна идут в спинном мозгу в боковых канатиках, образуя пирамидные лучки; неперекрещенные же образуют в передних канатиках спинного мозга прямые пирамидные лучки и переходят на другую сторону только в самом конце своего пути в спинном мозгу. Все волокна пирамидных пучков оканчиваются вокруг двигательных клеток передних рогов спинного мозга. Таким образом, каждое из полушарий головного мозга связано с периферическими двигательными нейронами противоположной стороны.
* (Продолговатый мозг старые анатомы называли луковицей спинного мозга - bulbus medulae spinalis; отсюда и произошло название "бульбарный", т. е. принадлежащий продолговатому мозгу.)
По бокам возле пирамид расположены белые овальные вытянутые тела - оливы (olivae), внутри которых имеются извилистые, окруженные толстым слоем белого вещества пластинки серого вещества, в которых лежат многими слоями нервные клетки. Их нейриты переходят во внутреннее белое вещество и большинство из них идет затем, перекрещиваясь, через так называемые веревчатые тела (corpora restiforrnia) вверх в мозжечок и вниз в спинной мозг. Веревчатые тела состоят из пучков, продолжающих центростремительные волокна боковых и задних канатиков спинного мозга. Часть этих волокон оканчивается в продолговатом мозгу, другая часть - в мозжечке, некоторые же волокна проходят и в вышележащие части мозга - четверохолмие и зрительные бугры. Все эти волокна также перекрещиваются или в начале своего пути в спинном мозгу, или же в продолговатом. Перекрест центростремительных путей в продолговатом мозгу носит название медиальной петли (lemniscus medialis). Таким образом, и периферические чувствительные нейроны связаны с полушариями противоположной стороны. Расходясь вверху под острым углом, веревчатые тела входят в мозжечок, образуя его нижние ножки.
На задней поверхности продолговатого мозга продолжается задняя продольная борозда. По обеим боковым сторонам от нее присоединяются к веревчатым телам уже известные нам чувствительные пучки - нежный (Голля) и клиновидный (Бурдаха), которые тянутся из спинного мозга.
Между нижними ножками мозжечка образуется треугольное пространство, обращенное вершиной вниз, а основанием кверху; выше него двумя расходящимися пучками волокон, выходящими из мозжечка (верхние ножки), также образуется треугольное пространство, но обращенное вершиной вверх, а основанием к основанию первого треугольника. Таким образом, на задневерхней поверхности продолговатого мозга образуется ромбовидное углубленное пространство - ромбовидная ямка (fossa rhomboidea), которая составляет дно IV желудочка мозга. Между верхними ножками мозжечка желудочек прикрыт тонкой пластинкой - передним мозговым парусом, а между веревчатыми телами - нижними ножками, над желудочком натянута другая тонкая пластинка - задний мозговой парус, в котором имеется срединное отверстие для оттока цереброспинальной жидкости из IV желудочка в подпаутинное пространство. Поверх парусов над желудочком лежит мозжечок. Передневерхний конец IV желудочка переходит в узкий канал - мозговой (сильвиев) водопровод (aquaeductus cerebri Sylvii), а задненижний - в центральный канал спинного мозга.
Вследствие перекреста и расхождения пучков белого вещества серое вещество продолговатого мозга расположено не равномерным слоем на дне ромбовидной ямки, а в виде отдельных скоплений (ядер) и внутри пучков белого вещества. Продолжение задних рогов спинного мозга образует чувствительные ядра, а продолжение передних - двигательные. В чувствительных ядрах оканчиваются периферические чувствительные волокна спинномозговых и черепномозговых нервов. В них же начинается второй нейрон, передающий раздражения к вышележащим частям мозгового ствола. В двигательных ядрах берут начало центробежные волокна черепномозговых нервов. На дне IV желудочка расположены ядра V, VI, VII, VIII, IX, X и XII пары черепномозговых нервов.
Составляя прямое продолжение спинного мозга, продолговатый мозг сходен с ним и по своим функциям. Прежде всего одной из его функций является передача нервных импульсов от периферии к центру и обратно, т. е. проводниковая функция. Он содержит ядра, дающие начало черепномозговым нервам; на дне ромбовидной ямки заложены центры для мышц лица. Но исключительно важное значение имеет продолговатый мозг вследствие наличия в нем центров, в которых замыкаются рефлекторные дуги громадного количества воспринимающих аппаратов и исполнительных органов тела.
Связи этих центров чрезвычайно многочисленны; так, например, ядра блуждающего нерва связаны со всеми внутренними органами. Здесь, в дне IV желудочка и отчасти внутри мозгового моста, располагаются весьма важные для жизни рефлекторные центры, регулирующие ряд основных функций тела. К числу их относятся центры дыхания, сердечной деятельности, глотания, рвоты, чихания, кашля, сосания, жевания, слюноотделения, моргания, потоотделения, сосудодвигательный и др. Кроме того, в продолговатом мозгу располагается также общий центр обмена веществ с центрами обмена воды и солей.
Таким образом, продолговатый мозг является не только местом передачи двигательных и чувствительных импульсов, но играет также огромную роль в регуляции рефлекторных актов, без которых жизнь организма невозможна. Вследствие этого его центры получили название жизненных центров. Расстройство их функций, или разрушение продолговатого мозга, ведет к неминуемой смерти вследствие прекращения дыхания и сердечной деятельности.
Мозжечок (cerebellum) и мозговой мост (pons cerebri Varolii) (рис. 278, 279, 280). Мозжечок располагается в задней черепной ямке позади продолговатого мозга. Над ним нависают затылочные доли больших полушарий мозга, от которых мозжечок отделен перепончатым отростком твердой мозговой оболочки - палаткой мозжечка (или, иначе, мозжечковым наметом). Мозжечок имеет очень сложное строение и состоит из трех частей. Средняя часть выступает в виде узкого валика с многочисленными поперечными перетяжками и называется червячком (vermis). Две боковые части носят название полушарий (hemisphaeria). Червячок является по своему происхождению более древним образованием, а полушария - более поздним. У различных животных мозжечок развит неодинаково. Например, у рыб и птиц, которые плавают и летают, т. е. производят сложные движения, мозжечок значительно более развит, чем у земноводных или пресмыкающихся животных.
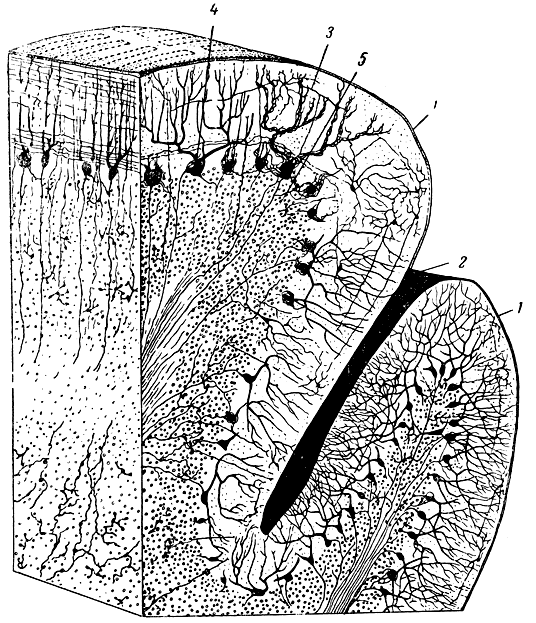
Рис. 278. Схема строения коры мозжечка. 1 - извилина мозжечка; 2 - борозда мозжечка; 3 - наружный зернистый слой; 4 - слой клеток Пуркинье; 5 - внутренний зернистый слой
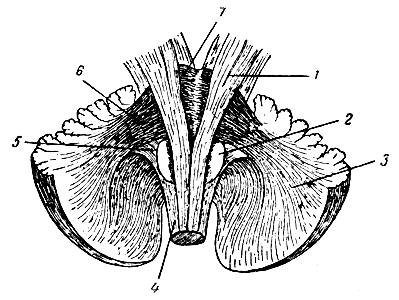
Рис. 279. Боковые ножки мозжечка от варолиева моста и нижние - от продолговатого мозга. 1 - пирамидный путь; 2 - олива; 3 - полушарие мозжечка; 4 - продолговатый мозг; 5 - ножки мозжечка к продолговатому мозгу; 6 - ножки мозжечка от варолиева моста; 7 - варолиев мост
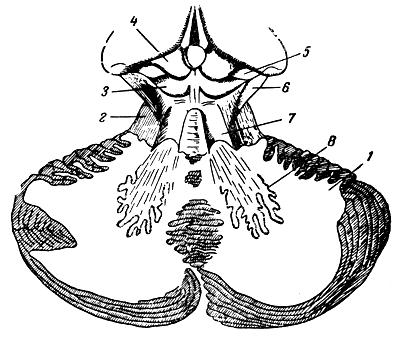
Рис. 280. Ножки мозжечка к четверохолмию (схема). Часть мозжечка удалена. 1 - мозжечок; 2 - ножка моста; 3 - задний бугорок четверохолмия; 4 - передний бугорок четверохолмия; 5 - внутреннее коленчатое тело; 6 - ножка мозга; 7 - ножки мозжечка к четверохолмию; 8 - зубчатое ядро
Поверхность полушарий мозжечка, как и поверхность больших полушарий, покрыта бороздками и извилинами (только более мелкими), а несколько более глубоких борозд делят каждое полушарие на доли. Серое вещество покрывает мозжечок снаружи, образуя кору мозжечка; белое вещество расположено внутри и на разрезе имеет ветвистые очертания. Если провести через мозжечок продольный разрез вдоль червячка, то сразу на разрезе бросится в глаза характерная фигура, напоминающая разветвленное дерево, поэтому старые анатомы и назвали эти разветвления белого вещества в мозжечке "древом жизни" (arbor vitae). Несмотря на свою небольшую величину, мозжечок обладает громадным количеством ганглиозных клеток, это объясняется значительно большей по сравнению с большим мозгом глубиной его борозд. На разрезах через мозжечок уже невооруженному глазу заметно относительно небольшое количество белого вещества по сравнению с серым веществом коры. Большинство ганглиозных клеток, находящихся в коре мозжечка, не выходит своими нейритами за ее пределы, что дает повод к предположению о существовании здесь собственного ассоциативного аппарата.
Из клеточных элементов мозжечковой коры (под микроскопом) обращают на себя внимание нервные клетки Пуркинье, которые имеют грушевидное, бедное пигментом тело с ясно выраженными глыбками Ниссля. От них отходят два мощных дендрита, которые распадаются на много ветвей и достигают свободной поверхности коры. Тела клеток Пуркинье окружены особыми фибриллярными корзиночками, фибриллы которых происходят от мультиполярных ганглиозных корзинчатых клеток. Нет сомнения, что фибриллярные корзинки являются очень сложными и очень важными структурными элементами коры мозжечка, передающими большинство раздражений, попадающих к клеткам Пуркинье.
Внутри полушарий в белом веществе мозжечка также заложены ядра серого вещества, в которых прерываются проводящие мозжечковые пути.
Прикрывая продолговатый мозг сверху и несколько сзади, мозжечок соединяется со всеми отделами мозга тремя парами ножек, представляющими собой совокупность нервных волокон. Нижние ножки образованы уже известными нам веревчатыми телами и соединяют полушария мозжечка с продолговатым и спинным мозгом, то есть нервные импульсы от рецепторов мышц, связок и суставов нашего тела направляются по нервным волокнам вверх через спинной и продолговатый мозг и по нижним ножкам в мозжечок. Другая часть нервных волокон нижних ножек несет нервные импульсы от полукружных каналов внутреннего уха, т. е. от органа, посредством которого человек получает представление о своем положении в пространстве. Верхние ножки состоят из волокон, соединяющих полушария мозжечка с задними буграми четверохолмия и вышерасположенными отделами мозга. Средние ножки являются самыми мощными. Они соединяют оба полушария мозжечка между собой и ядра серого вещества мозгового моста с полушариями мозжечка, т. е. через эти ножки волокна из коры мозжечка, направляются к ядрам моста, а оттуда в кору больших полушарий.
Таким образом, через средние ножки осуществляется связь между корой мозжечка и корой большого мозга.
Мозжечок занимает особое положение, будучи промежуточным звеном между стволовой частью и полушариями. Центры мозжечка чрезвычайно многочисленны; они заложены как внутри него, в серых ядрах, так и снаружи - в коре. Мозжечок связан со всеми отделами нервной системы: с корой больших полушарий, со спинным мозгом, продолговатым, средним и серыми ядрами внутри мозгового моста.
Основной функцией мозжечка является рефлекторная координация движений. В мозжечке сосредоточены центры бессознательной координации работы мышц и равновесия. У низших позвоночных, медленно и неуклюже ползающих, мозжечок развит слабо. Среди млекопитающих животных мозжечок достигает большого развития, особенно у хороших бегунов и летунов. Весьма подвижные приматы и вертикально ходящий человек обладают сильно развитым мозжечком.
В полушариях мозжечка находятся центры, регулирующие движения верхних и нижних конечностей, а в червячке - центры регуляции движений туловища и головы.
Вот почему удаление мозжечка сопровождается потерей координации движений, их силы и объема; при этом страдают такие двигательные акты, как письмо, ходьба, речевые движения, трудовые движения.
Мозжечок непосредственно связан с деятельностью больших полушарий мозга. Ученые считают, что импульсы мозжечка тормозят, задерживают рефлексы спинного, продолговатого и среднего мозга и тем самым обеспечивают возможность наиболее полного влияния импульсов больших полушарий. Так, мозжечок тормозит, например, многочисленные рефлексы, возникающие в нервных окончаниях мышц, сухожилий, связок при каждом движении. Деятельность мозжечка обусловливает собой точность, четкость движений тела, как бы производит "отделку" движений.
Мозговой (варолиев) мост представляет собой белое утолщение мозгового вещества, состоящее из широкого пучка поперечно идущих волокон, расположен на передней стороне мозгового ствола тотчас выше (кпереди) продолговатого мозга также на скате затылочной кости. Сзади от мозгового моста по обе стороны отходят вниз под мозжечок объемистые пучки нервных волокон - описанные выше средние ножки, соединяющие оба полушария мозжечка. Кпереди от мозгового моста отходят также мощные пучки нервных волокон - это ножки мозга (о них речь ниже).
Внутри моста проходят пирамидные пути, расположены ядра серого вещества, от которых часть волокон идет к коре мозжечка, а часть поднимается после перекреста выше. Вместе с волокнами от ядер в кору мозжечка направляются боковые ветви пирамидных волокон, связывающих мозжечок с корой больших полушарий.
В скоплениях серого вещества, находящихся в мозговом мосту, помещаются центры рефлекторных движений рук, ног, туловища и др., т. е. центры такого же рода, как и в спинном мозгу, но центры мозгового моста более высокого порядка, чем центры спинномозговые, они заведуют сочетанными сокращениями мышц конечностей и туловища, которые нужны для тех или иных сложных движений.
Четверохолмие (corpus quadrigeminum) и ножки мозга (pedunculi cerebri) (рис. 281). Часть мозгового ствола, лежащая впереди от мозгового мозга, является менее развитой с толстыми стенками и узкой полостью - мозговым (сильвиевым) водопроводом, соединяющим полости IV и III мозговых желудочков. Верхняя стенка представляет собой пластинку с четырьмя бугорками (холмиками) белого вещества - четверохолмие. В каждом из холмиков залегают скопления серого вещества - ядра, при этом ядра передних двух холмиков связаны со зрительными проводящими путями, а ядра задних двух - со слуховыми. Таким образом, четверохолмие служит как бы промежуточной станцией на пути зрительных нервов к полушариям. Однако разрушение четверохолмия у человека не ведет к слепоте, так как у человека лишь небольшая часть волокон зрительного нерва направляется к четверохолмию, большая же часть их оканчивается в ганглиозном узле латерального коленчатого тела (corpus geniculatum laterale).
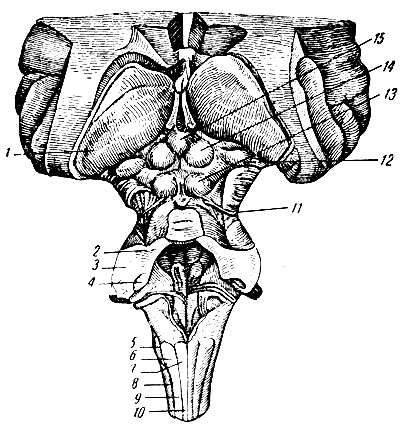
Рис. 281. Продолговатый и средний мозг сзади (мозжечок отрезан). 1 - подушка (задняя часть зрительного бугра); 2 - часть ножки мозжечка к четверохолмию; 3 - часть ножки мозжечка к мосту; 4 - часть ножки мозжечка к продолговатому мозгу; 5 - олива; 6 - ядро клиновидного пучка (ядро Бурдаха); 7 - ядре нежного пучка (ядро Голля); 8 - боковой пучок; 9 - клиновидный пучок; 10 - нежный пучок; 11 - блоковый нерв; 12 - медиальное коленчатое тело; 13-14 - четверохолмие; 15 - шишковидная железа
Верхняя пара холмиков (зрительные холмики) представляет собой древний зрительный аппарат мозга, к которому позднее присоединилась вторая нижняя пара (слуховых холмиков). Вокруг мозгового водопровода также расположен слой серого вещества, в котором заложены ядра двух черепномозговых нервов - глазодвигательного (III пара) и блоковидного (IV пара). У низших животных (например, у лягушек и рыб) вместо четверохолмия имеется зрительное двухолмие, разрушение которого ведет к полной слепоте. Из каждого бугорка четверохолмия в сторону отходят по одному белому тяжу нервных волокон - это так называемые ручки четверохолмия. В промежутке между передними бугорками располагается шишковидная железа - эпифиз.
Нижнюю стенку этой части ствола образуют два мощных пучка волокон белого вещества, расходящихся под углом кверху и уходящих в глубь полушарий к зрительным буграм и выше - это ножки мозга. В них проходят центробежные волокна от вышележащих частей мозга, через мост и пирамиды к спинному мозгу, а также и центростремительные волокна из продолговатого и спинного мозга, направляющиеся к вышележащим отделам мозга. Все волокна, проходящие в ножках мозга, в разных местах постепенно подвергаются перекресту, так что в конце концов правая ножка оказывается собранием двигательных и чувствительных путей левой половины тела, а левая ножка заключает в себе двигательные и чувствительные пути правой половины тела. В ножках мозга различают нижнюю часть (основание ножек), в которой проходят двигательные нервные волокна, и верхнюю часть, примыкающую непосредственно к четверохолмию (покрышку), в которой проходят чувствительные волокна.
Внутри ножек заложены ядра серого вещества в виде двух скоплений: между основанием и покрышкой располагается так называемое черное вещество (substantia nigra)*, клетки которого содержат большое количество черного пигмента - меланина, от него волокна отходят к мосту и мозжечку; на уровне передних бугорков четверохолмия вверху покрышки ножек располагается красное ядро (nucleus ruber), от которого отходят волокна к вышележащим частям мозга. В его клетках обнаруживается в небольшом количестве железо, которое, по-видимому, и придает ядру красноватую окраску. Сбоку от красного ядра в ножках мозга проходит общий пучок всех чувствительных проводящих путей - это уже упомянутая выше медиальная петля (lemniscus medialis); несколько сбоку от нее располагается пучок слуховых нервных волокон - латеральная петля (lemniscus lateralis). Чувствительные волокна медиальной петли направляются из ножек мозга дальше в зрительные бугры. В составе латеральной и медиальной петель проходят вверх к большому мозгу все чувствительные проводящие пути, за исключением зрительного и обонятельного. Каждая из мозговых ножек, войдя в полушарие мозга, поднимается вверх и образует участок белого вещества, лежащий кнаружи от зрительного бугра. Этот участок носит название внутренней, капсулы (capsula interna); она является весьма важным отделом белого вещества полушарий, через который проходят главные двигательные и чувствительные пути.
* (Черное вещество имеется только у человека и появляется на третьем году его жизни.)
В четверохолмии находятся нейроны рефлекторных дуг, связанные со световыми и звуковыми раздражителями. Так, здесь, например, происходит передача раздражений, вызывающих сочетанные движения глаз. В передних бугорках четверохолмия оканчивается часть центростремительных нервных волокон, которые являются нейритами вторых нейронов зрительного пути сетчатки. В задних бугорках четверохолмия оканчиваются нейриты вторых нейронов слухового пути. Таким образом, четверохолмия, не являясь центрами зрения и слуха, при действии на глаз и ухо световых и звуковых раздражителей осуществляют рефлексы движения (поворота) туловища, головы, глаз и ушей в сторону раздражителей.
Благодаря связи с воспринимающими аппаратами (глаз, ухо, мышцы) центры четверохолмия регулируют сложные двигательные акты. Средний мозг регулирует также распределение тонуса поперечнополосатой мускулатуры. Его центры принадлежат главным образом к моторной системе, имеющей большое значение для регуляции автоматических движений.
Зрительные бугры (thalami optici). В состав промежуточного мозга входят зрительные бугры, подбугровая область и III желудочек мозга, расположенный между правым и левым бугром. Кроме того, сюда же примыкают две эндокринные железы мозгового происхождения - гипофиз и эпифиз.
Зрительные бугры являются древнейшей частью головного мозга. Они расположены впереди от четверохолмия и представляют собой два довольно больших скопления серого вещества мозга. С боков они сливаются с массой полушарий, а медиальные их поверхности составляют стенки вертикально расположенной щели - полости III желудочка, который двумя боковыми межжелудочковыми отверстиями (монроевы отверстия) сообщается с боковыми желудочками мозга; сзади в него открывается мозговой водопровод. На границе верхней и внутренней поверхности зрительных бугров проходит мозговая полоска, кончающаяся сзади спайкой поводков, к последней прикреплена шишковидная железа (эпифиз). Нижняя поверхность выдается на основании мозга в виде серого бугра и называется подбугровой областью. Вершина бугра вытянута в воронку, на которой подвешен гипофиз (hypophysis), помещающийся в ямке турецкого седла. Верхняя стенка III желудочка очень тонка и прикрыта мягкой оболочкой мозга, образующей здесь сосудистое сплетение. Серое вещество зрительных бугров составляет продолжение серого вещества мозгового водопровода. Структура бугров очень сложная и отличается большим разнообразием ганглиозных клеток как по форме, так и по их распределению. В буграх заложены ядра, от которых отходят центростремительные волокна к кюре больших полушарий; центробежные волокна от ядер спускаются к нижележащим частям - ножкам мозга, четверохолмию, мозжечку. Почти все чувствительные проводящие пути собираются в зрительных буграх. Всякий центростремительный импульс, прежде чем достичь коры большого мозга, должен пройти через зрительные бугры, которые служат, таким образом, промежуточным этапом для всех чувствительных путей, направляющихся к коре мозга. Зрительные бугры связаны и с подкорковыми узлами полушарий. Некоторые ученые предполагают, что зрительные бугры активно участвуют в деятельности коры мозга и от них зависит возникновение чувства "приятного" и "неприятного" или, как говорят, та или иная "окраска ощущений".
Большие полушария мозга. Большие полушария мозга и прилежащие к ним части (рис. 282, 283, 284, 285, 286, 287, 288). Из первого мозгового пузыря, как мы уже знаем, развивается передний мозг. У человека в передний мозг входят большие полушария мозга с прилегающими к ним обонятельными луковицами, мозолистым телом, сводом и полосатым телом.
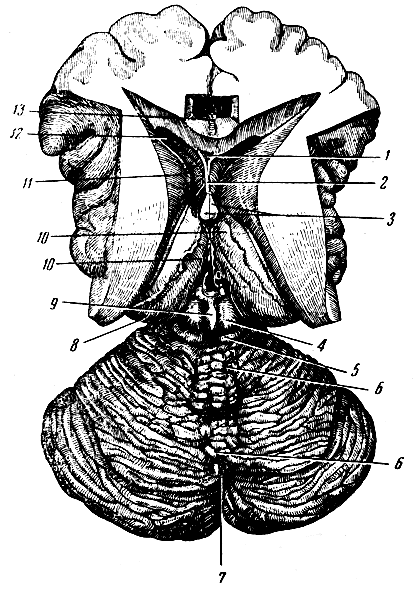
Рис. 282. Средний мозг и мозжечок. 1 - полость прозрачной перегородки; 2 - прозрачная перегородка; 3 - колонна свода в поперечном разрезе; 4-5 - четверохолмие; 6-7 - мозжечок; 8 - подушка - задняя часть зрительного бугра; 9 - шишковидная железа (эпифиз); 10 - зрительный бугор; 11 - головка хвостатого тела; 12 - передний рог бокового желудочка; 13 - колено мозолистого тела
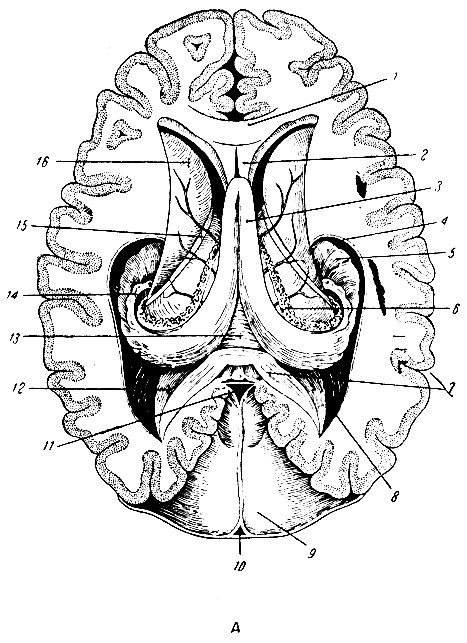
Рис. 282а. Вскрытые сверху желудочки мозга и мозговой свод (горизонтальный разрез мозга). 1 - мозолистое тело; 2 - прозрачная перегородка; 3 - свод мозга; 4 - нижний рог; 5 - аммонов рог; 6 - сосудистое сплетение; 7 - мозолистое тело; 8 - задний рог; 9 - мозжечковый намет; 10 - верхний сагиттальный синус; 11 - прямой синус; 12 - птичья шпора; 13 - спайка морского коня; 14 - бахромка морского коня; 15 - полоска зрительного бугра; 16 - головка хвостатого ядра
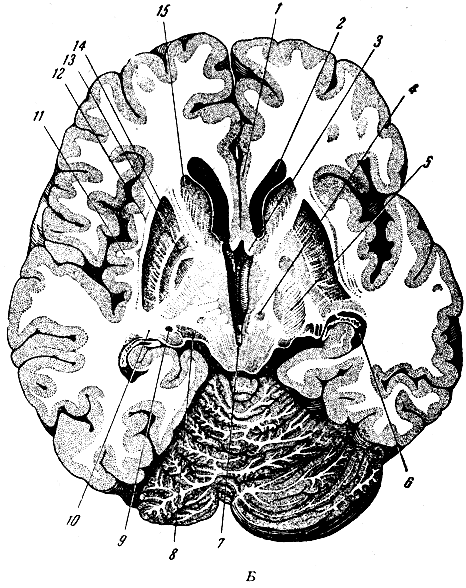
Рис. 282б. Полушария мозга и мозжечка на горизонтальном разрезе. 1 - подмозолистая извилина; 2 - передний рог бокового желудочка; 3 - свод мозга; 4 - красное ядро; 5 - ножки мозга у перехода во внутреннюю капсулу; 6 - боковой желудочек (нижний рог и сосудистое сплетение); 7 - водопровод мозга; 8 - медиальное коленчатое тело; 9 - латеральное коленчатое тело; 10 - свод мозга (задние ножки); 11 - островок; 12 - ограда; 13 - бледный шар; 14 - скорлупа; 15 - головка хвостатого ядра
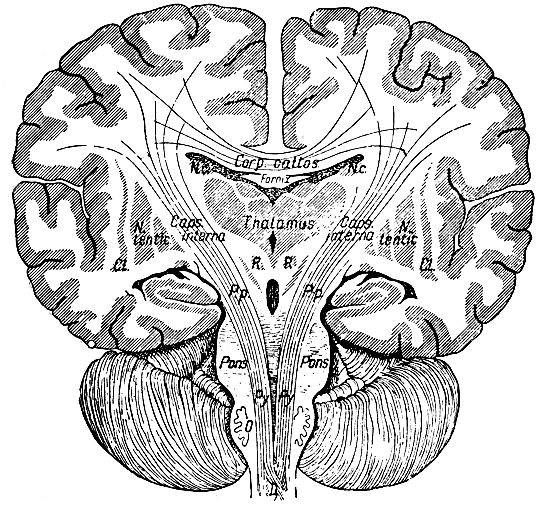
Рис. 283. Фронтальный разрез головного мозга. Corp. callos. - мозолистое тело; Thalamus - зрительный бугор; Caps. interna - внутренняя капсула; Р. р. - ножки мозга; Ру - пирамидный путь; Pons - варолиев мост; N. с. - хвостатое тело; N. lentic и Сl. - полосатое тело; R - красное ядро; Fornix - свод; D - перекрест пирамидных путей; О - олива
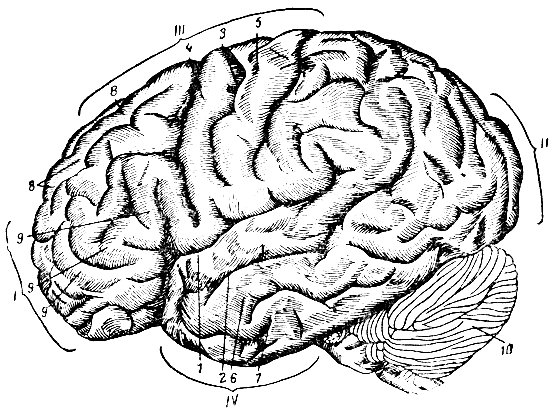
Рис. 284. Борозды и извилины на наружной поверхности полушарий головного мозга. I - лобная доля; II - затылочная; III - теменная; IV - височная; 1 - сильвиева борозда; 2 - верхняя височная борозда; 3 - роландова борозда; 4 - передняя центральная извилина; 5 - задняя центральная извилина; 6 - средняя височная извилина; 7 - нижняя височная извилина; 8 - верхняя лобная извилина; 9 - нижняя лобная извилина; 10 - мозжечок
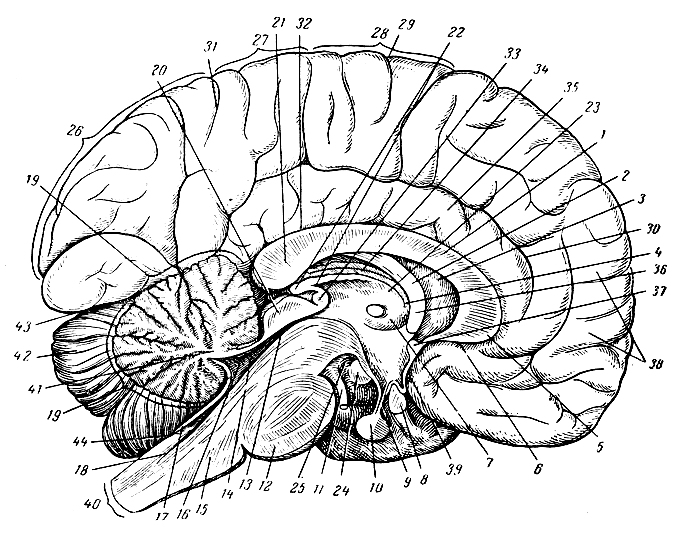
Рис. 285. Сагиттальный разрез головного мозга. 1 - мозолистое тело; 2 - свод; 3 - прозрачная перегородка; 4 - монроево отверстие; 5 - колено; 6 - клюв мозолистого тела; 7 - полость третьего желудочка; 8 - зрительный перекрест (в разрезе); 9 - воронка; 10 - придаток мозга; 11 - глазодвигательный нерв; 12 - варолиев мост; 13 - сильвиев водопровод; 14 - передний мозговой парус; 15 - продолговатый мозг; 16 - четвертый желудочек; 17 - задний мозговой парус; 18 - центральный канал спинного мозга; 19 - разрез червячка мозжечка; 20 - четверохолмие; 21 - утолщение мозолистого тела; 22 - шишковидное тело; 23 - боковая стенка третьего желудочка (зрительный бугор); 24 - титечное тело; 25 - заднее продырявленное пространство; 26 - часть затылочной доли (клин); 27-28 - теменная доля; 29 - верхний конец роландовой борозды; 30 - поясная борозда; 31 - теменно-затылочная борозда; 32 - борозда мозолистого тела; 33 - задняя спайка; 34 - сосудистая покрышка третьего желудочка; 35 - поясная извилина; 36 - передняя ножка свода; 37 - передняя спайка; 38 - лобная доля; 39 - передняя стенка третьего желудочка; 40 - место разреза продолговатого мозга у места перехода его в спинной мозг; 41 - белое вещество мозжечка; 42 - полушарие мозжечка; 43 - нижний край затылочной доли; 44 - отверстие Мажанди
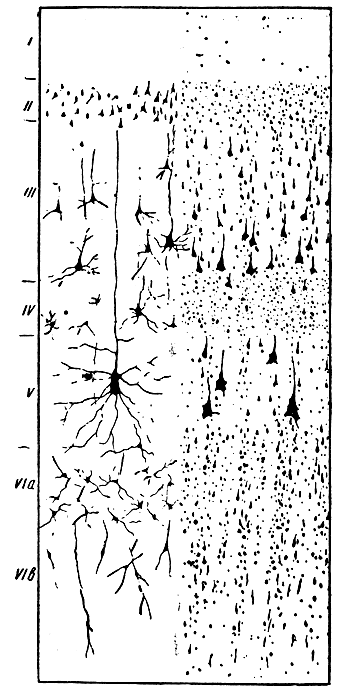
Рис. 286. Схема клеточного строения коры головного мозга. I - поверхностный опоясывающий слой молекулярных клеток; II - наружный слой зернистых клеток; III - слой пирамидных клеток средней и большой величины; IV - внутренний слой зернистых клеток; V - слой узловых гигантских пирамидных (Беца) клеток; VIa и VIв - слой многообразных (веретенообразных и полиморфных) клеток
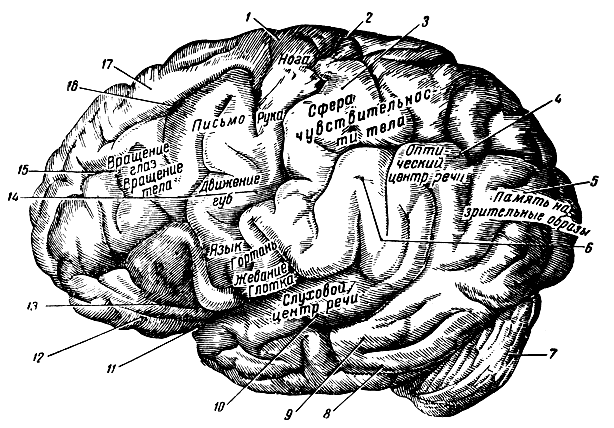
Рис. 287. Боковая поверхность левого полушария мозга с обозначением на ней областей коры мозга по функции. 1 - передняя центральная извилина; 2 - центральная борозда; 3 - задняя центральная извилина; 4 - угловая извилина; 5 - затылочная доля; 6 - надкраевая извилина; 7 - мозжечок; 8 - нижняя височная извилина; 9 - средняя височная извилина; 10 - верхняя височная извилина; 11 - боковая щель мозга (сильвиева); 12 - передняя горизонтальная ветвь; 13 - передняя восходящая ветвь сильвиевой борозды; 14 - передняя центральная извилина; 15 - средняя лобная извилина; 16 - верхняя лобная борозда; 17 - верхняя лобная извилина
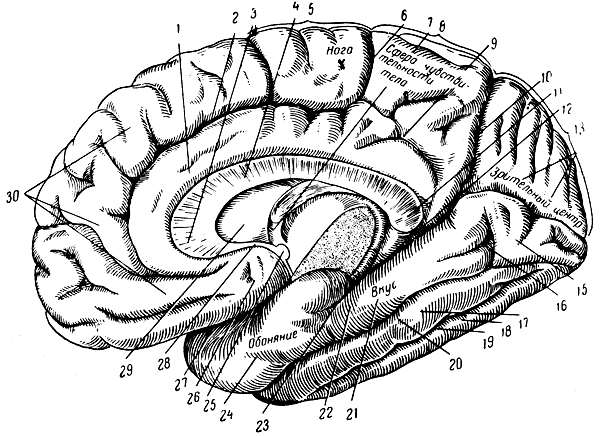
Рис. 288. А. Медиальная поверхность правого полушария мозга. 1 - поясная извилина; 2 - поясная борозда; 3 - колено мозолистого тела; 4 - ствол мозолистого тела; 5 - околоцентральная долька; 6 - поясная борозда (краевая часть); 7 - свод; 8 - предклинье; 9 - подтеменная борозда; 10 - теменно-затылочная щель; 11 - утолщение мозолистого тела; 12 и 23 - борозда морского коня; 13 - клин; 14 - шпорная борозда (под надписью 'зрительный центр'); 15, 16 - язычная извилина; 17 - веретенообразная извилина; 18 - нижняя височная извилина; 19 - нижняя височная борозда; 20 - обходная щель; 21 - корковый центр вкуса; 22 - извилина морского коня; 24 - корковый центр обоняния; 25 - передняя спайка; 26 - околообонятельный кружок (Брока); 27 - латеральная щель мозга (Сильвия); 28 - клюв мозолистого тела; 29 - прозрачная перегородка; 30 - верхняя лобная извилина
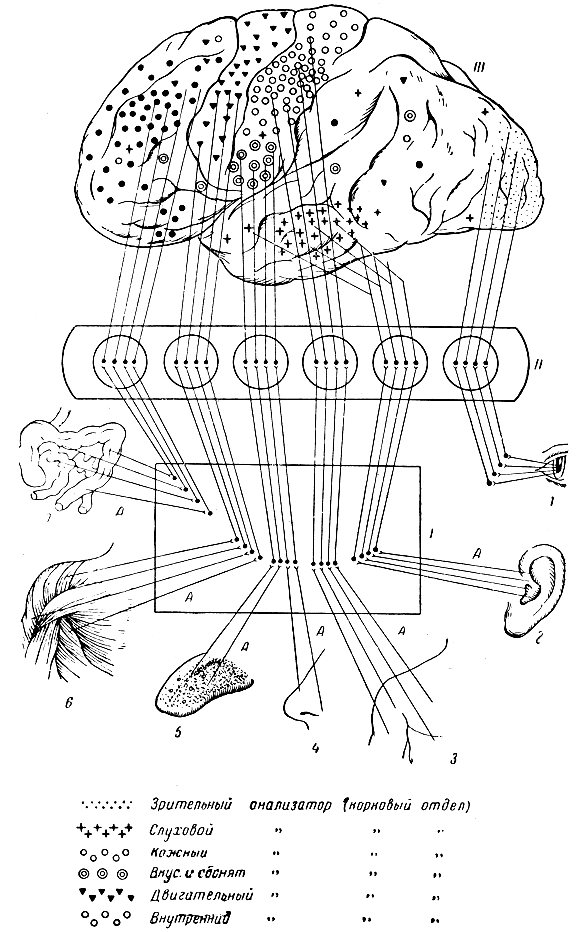
Рис. 288. Б. Схема, иллюстрирующая строение анализаторов. 1-7 - рецепторы (зрительный, слуховой, кожный, обонятельный, вкусовой, двигательный, внутренних органов): 1 - область спинного или продолговатого мозга, куда вступают афферентные волокна (А), импульсы с которых передаются на расположенные здесь невроны, образующие восходящие пути; аксоны последних идут до области зрительных бугров (II); аксоны нервных клеток зрительных бугров восходят в кору мозга (III). Вверху (III) намечено расположение ядерных частей корковых отделов различных анализаторов (для внутреннего, вкусового и обонятельного анализаторов это расположение еще не точно установлено): указаны также разбросанные по коре рассеянные клетки каждого анализатора (по К. М. Быкову, Учебник физиологии)
Наиболее старой по происхождению частью переднего мозга, которая непосредственно прилегает к промежуточному, является так называемое толстое тело (corpus striatum), расположенное латерально от внутренней капсулы. Оно представляет собой ряд скоплений серого вещества в глубине под большими полушариями. В нем различают три пары ядер: хвостатое тело, чечевицеобразное тело и ограду, которые являются важными подкорковыми центрами для регуляции автоматических движений и вегетативных функций. Эти ядра отделены друг от друга прослойками белого вещества, вследствие чего на поперечном разрезе мозга эта часть выглядит полосатой (отсюда и название "полосатое тело").
Часть чечевицеобразного тела носит название бледного шара (globus pallidus), которое является двигательным центром. Чувствительные импульсы с нейронов зрительного бугра переключаются здесь на нейрон бледного шара. Отсюда двигательные импульсы идут вниз последовательно в промежуточный, средний, продолговатый и спинной мозг. Бледное ядро, таким образом, контролирует и согласует деятельность всех нижележащих отделов стволовой части мозга и является по отношению к ним высшим и наиболее поздним по происхождению отделом. Оказывается, импульсы, идущие от бледного шара вниз, как правило, вызывают торможение нижележащих двигательных центров. У млекопитающих и человека бледный шар, как и зрительные бугры, подчиняется полосатому телу и коре больших полушарий. Все сложные врожденные рефлексы регулируются зрительными буграми и бледным шаром. Сюда относятся защитные и оборонительные движения и инстинкты (сложные врожденные двигательные рефлексы), которые совершаются помимо нашего сознания, например пищевой и половой инстинкты, закрывание глаз рукой при внезапном ярком свете и др.
Полосатое тело по отношению к бледному шару представляет собой высший двигательный центр. Импульсы полосатого тела регулируют деятельность бледного шара. Так, например, повреждение полосатого тела ведет к усилению рефлекторных защитных движений: при каком-либо малейшем раздражении (например, шорох) животное уже настораживается и принимает оборонительную позу. Импульсы от полосатого тела тормозят рефлексы бледного шара, а через него и других нижележащих отделов мозга. Благодаря деятельности полосатого тела все инстинктивные движения животных и человека становятся менее резкими, более законченными, соответствующими полученным раздражениям. Кроме того, считают, что в полосатом теле заложены также высшие центры вегетативной нервной системы, как, например, центры теплорегуляции и углеводного обмена.
Полосатое тело у низших позвоночных составляет главную массу полушарий. У человека оно получает волокна из зрительного бугра, из коры больших полушарий и от многих подкорковых нервных узлов.
Большие полушария (hemisphaerae cerebri) покрывают сверху всю стволовую часть мозга. Они составляют почти 6/7 всей массы головного мозга. Верхняя и боковая поверхности полушарий вместе представляют почти правильную сферическую форму. Медиальные поверхности полушарий, обращенные друг к другу, плоские, а нижние поверхности полушарий, лежащие на основании черепа, покрыты выступами и углублениями. Посредине нижней поверхности тянется мозговой ствол, который ближе кпереди уходит в глубь полушарий. Сверху посредине глубокая продольная борозда (fissura longitudinalis cerebri) разделяет одно полушарие от другого. В глубине этой борозды лежит толстая и плотная пластинка из белого вещества - большая спайка мозга, или мозолистое тело (corpus callosum). Волокна мозолистого тела соединяют оба полушария между собой. Под мозолистым телом располагается свод (fornix), состоящий из двух дугообразно идущих пучков нервных волокон, подпирающих мозолистое тело снизу.
На поверхности каждого полушария различают пять долей: лобную, теменную, затылочную, височную и рейлиев островок, скрывающийся в глубине сильвиевой борозды.
Борозды и извилины больших полушарий. На поверхности полушарий имеются глубокие щели и многочисленные борозды (fissurae et sulci), между которыми в виде изогнутых валиков выступают извилины (gyri) мозга. Все эти неровности значительно увеличивают общую поверхность мозговых полушарий. В животном мире извилины развиваются только у высших млекопитающих, особенно хорошо они развиты у человекообразных обезьян. Но у четырехмесячного плода, например, наружная поверхность полушарий почти еще гладкая. На пятом месяце внутриутробной жизни на гладкой поверхности полушарий возникают полоски быстро утолщающейся нервной ткани. Эти утолщающиеся участки принимают вид валиков. Активную роль в процессе образования борозд играют извилины, а борозды возникают вторично.
Самой ранней по времени развития является обширная боковая (сильвиева) борозда (sulcus cerebri lateralis Sylvii), которая является границей между височной долей с одной стороны, лобной и теменной - с другой. У зародыша края этой борозды сильно раздвинуты и между ними выступает центральная доля. К моменту рождения сильвиева борозда смыкается и закрывает центральную долю. Вторая большая борозда идет в поперечном направлении и разделяет лобную и теменную доли между собой - это центральная, или роландова, борозда (sulcus centralis Rolandi), которая появляется при дальнейшем росте полушарий на шестом месяце внутриутробной жизни; вместе с ней появляются зачатки и других главных борозд. Более мелкие борозды продолжают развиваться еще и после рождения. Кпереди от роландовой борозды, параллельно ей, идет передняя центральная извилина (gyrus centralis anterior). Позади роландовой борозды на теменной доле располагается задняя центральная извилина. Сзади между полушариями и мозжечком заметна глубокая поперечная щель мозга (fissura transversa cerebri). На внутренней поверхности полушарий расположены борозда мозолистого тела, затылочно-теменная, шпорная и др.
Другие менее важные борозды разделяют поверхность каждой доли на извилины и дольки. Так, в лобной и височной долях проходят по две горизонтальные борозды, разделяющие каждую из долей на три извилины (верхнюю, среднюю и нижнюю). В теменной и затылочной долях ход борозд и расположение извилин не так правильны. На медиальной поверхности полушарий, кроме ряда других, более крупной является сводная извилина (gyrus fornicatus) с крючком (uncus), в котором расположен центр обоняния. Эта извилина залегает над мозолистым телом, а под мозолистым телом проходит извилина аммонова рога (gyms hyppocampi) - гиппокампа (морского коня).
Кора больших полушарий. Снаружи полушария покрыты тонким слоем серого вещества, носящего название коры мозга, толщина которой в разных местах колеблется от 2 до 4 мм. Серое вещество составляет около 40% общего объема большого мозга. Количество нервных клеток, сосредоточенных здесь, больше, чем в каких-либо других частях нервной системы, и достигает приблизительно 14 млрд.
Микроскопически кора (рис. 286) состоит из шести слоев различных нервных клеток, расположенных этажами друг над другом*. Первый наружный слой содержит мало нервных клеток и много горизонтальных нервных волокон, он состоит главным образом из густого глиального синцития и концевых ветвей дендритов нервных клеток, лежащих в глубоких слоях коры; во втором и четвертом слое - масса мелких нервных клеток. Обращает на себя внимание громадное разнообразие нервных клеток как по их форме и величине, так и по количеству и расположению. Например, в коре встречаются малые, средние и большие пирамидальные клетки (третий и пятый слои); в коре передней центральной извилины содержатся так называемые бецовские гигантские пирамидные клетки (до 120 μ), являющиеся клетками моторных пирамидных путей. Они сходны с моторными клетками мозгового ствола и спинного мозга. В шестом слое имеются мелкие нервные клетки разнообразной формы.
* (Послойное распределение клеточных элементов коры мозга принадлежит замечательному русскому анатому В. А. Бецу (1874).)
Пирамидные клетки являются основой клеточного строения коры полушарий. Они обладают различной величиной, но сходны по строению. Их тело имеет форму конуса, обращенного вершиной к поверхности полушарий. Основание конуса слегка выпукло. От центра основания отходит нейрит, направляющийся в глубокие слои серого вещества и переходящий в белое вещество. От нейритов отходят в стороны перпендикулярные отростки, которые, по-видимому, обеспечивают связь с другими пирамидными клетками. Основной дендрит пирамидной клетки направляется к поверхности, где и разветвляется. Другие нервные клетки коры имеются в меньшем количестве и они иного строения: часть их составляет передаточный и связующий аппарат, другая часть посылает нейриты в пирамидные пути.
Соотношение толщины корковых слоев и характер самих клеток у различных людей индивидуально меняются. Различен корковый слой также в отдельных его участках. В мозговой коре, как и во всяком сером веществе, где бы оно ни находилось, кроме нервных клеток со всеми их отростками, всегда имеется и глиозная ткань (нейроглия).
Направление волокон в полушариях различно: одни идут от нервных клеток одного полушария к нервным клеткам того же полушария, другие связывают различные пункты обоих полушарий; третьи, наконец, вступают в мозговые ножки, при помощи которых, как мы уже знаем, полушария соединяются с продолговатым и спинным мозгом. В коре оканчиваются центростремительные волокна, поднимающиеся из лежащих ниже частей мозга, и начинаются центробежные волокна, идущие к лежащим ниже частям. Рефлекторные центры коры и их связи с лежащими ниже частями мозга неизмеримо сложнее по сравнению с подобными же центрами стволовой части.
Деятельность коры больших полушарий включает и объединение (интегрирование) функции подкорковых центров. Полосатое тело доминирует над промежуточным мозгом, который объединяет и соматические, и вегетативные функции, в свою очередь подчиняя себе деятельность среднего мозга. Средний же мозг доминирует над продолговатым мозгом.
Кора мозга обильно снабжается кровью и потребляет в 10-20 раз больше кислорода, чем другие ткани. Обмен веществ даже в покое значительно выше, чем обмен веществ в нерве. Кора чрезвычайно чувствительна к нарушениям кровоснабжения и к кислородному голоданию.
Например, нервные клетки коры больших полушарий погибают уже через 5-6 минут после прекращения кровоснабжения.
Нервные клетки коры также более чувствительны к действию некоторых ядов (наркотики, алкоголь и др.), чем клетки других тканей. При действии наркоза деятельность нервных центров временно парализуется.
Кора головного мозга - это орган высшей нервной деятельности, в основе которой лежит условный рефлекс. С понятием о ней необходимо в кратком виде быть знакомым и при изучении анатомии*.
* (Излагаемое ниже - краткое извлечение из учебника А. Г, Гинецинского и Л. В. Лебединского "Курс нормальной физиологии", М., 1956.)
Каждое явление в организме зависит от рефлекторной деятельности нервной системы.
Постоянные, неизменные, врожденные реакции организма осуществляются благодаря деятельности низших отделов центральной нервной системы - это так называемые безусловные рефлексы. Безусловные рефлексы - это рефлексы, вызываемые раздражением рецепторов, которые:
1. Всегда одинаково наступают у животных данного вида в ответ на раздражение определенного рецепторного поля данного рефлекса.
2. Могут осуществляться и в отсутствие высших отделов центральной нервной системы.
Безусловные рефлексы осуществляются подкорковыми образованиями центральной нервной системы, в которых сосредоточены аппараты, ведающие жизненно важными функциями, необходимыми для существования организма (например, кровообращение, обмен). Их ответные реакции одинаковы у всех особей данного вида. К безусловным рефлексам относятся: мигание при раздражении роговицы, кашель при попадании инородного тела в дыхательные пути, инстинкты (пищевой, половой, оборонительный). Характер безусловных рефлексов зависит от того, какой рецептор раздражен, и от силы его раздражения.
При безусловном рефлексе между внешним агентом (раздражителем) и ответной реакцией имеется постоянная связь. Безусловный рефлекс годен для уравновешивания деятельности организма лишь при абсолютном постоянстве внешней среды. Но организм может существовать в постоянно изменяющейся окружающей среде лишь в том случае, если он отвечает на эти изменения такими реакциями, которые обеспечивают его уравновешивание с этими условиями.
Уравновешивание в его высшей форме достигается благодаря деятельности наиболее высоко организованного и наиболее реактивного отдела центральной нервной системы у высших позвоночных животных - коры головного мозга.
Наиболее совершенной динамической формой взаимодействия организма с постоянно изменяющимися условиями окружающей среды являются условные рефлексы. Условные рефлексы формируются в течение жизни и осуществляются путем возникновения временных связей в коре мозга. Условные рефлексы оказываются выработанными тогда, когда в кюре мозга устанавливается замыкание, обеспечивающее передачу возбуждения с корковых клеток, воспринимающих действие условного раздражителя, на эфферентные невроны, а через них на эффекторы, выполняющие рефлекс.
Кора головного мозга - орган условно-рефлекторной деятельности, орган высшего анализа и синтеза.
Вся поверхность больших полушарий - кора - занята воспринимающими центрами, так называемыми мозговыми концами анализаторов.
Анализатором называется система нервных образований, посредством которой организм разлагает на отдельные элементы сложные комплексы раздражителей из внешней и внутренней среды и обеспечивает этим различное реагирование организма на разнообразные раздражения. Анализатор состоит из периферического аппарата - рецептора, воспринимающего раздражение, возникшее под влиянием действия агента или изменения внешней или внутренней среды, нервного проводника, передающего возникший в рецепторе под влиянием раздражения импульс в центральную нервную систему, и клеток центральной нервной системы, воспринимающих этот центростремительный импульс.
Корковый отдел или корковый конец каждого анализатора есть образование мозговой коры, в который адресуются импульсы от рецепторов, составляющих периферический отдел данного анализатора.
Кора состоит из корковых отделов различных анализаторов, имеющих на поверхности постоянную локализацию. Эти отделы нерезко отграничены, заходят, перекрывают друг друга краями. Каждый анализатор состоит из ядерной и рассеянной части. Ядерная часть представляет точную и подробную проекцию в коре всех элементов периферического рецептора и является необходимой для осуществления сложных форм условнорефлекторной деятельности, для выработки тонких дифференцировок. Разбросанные части могут встречаться далеко от расположения ядерной части. При удалении участка коры, заключающего ядро данного анализатора, исчезают условные рефлексы на раздражения, которые поступают (адресуются) в удаленный участок коры. Различают корковые отделы зрительного (светового), слухового (звукового) анализаторов, обонятельного, вкусового, кожного (тактильного, болевого, температурного), внутреннего (интерорецептивного) и мышечного (двигательного) анализаторов.
Наличие у каждой группы рецепторов особо повышенной восприимчивости лишь к определенным колебаниям (изменениям) среды, к действию определенных агентов, улавливаемых данным видом рецепторов, обеспечивает уже примитивный анализ. Дальнейший этап анализа осуществляется низшими отделами центральной нервной системы; благодаря их деятельности раздражения рецепторов вызывают различные безусловные рефлексы. Поэтому в каждом анализаторе в качестве его проводникового отдела включается (вслед за периферическим) низший отдел центральной нервной системы.
Высший анализ достигается только аналитической деятельностью коры - корковым отделом анализатора.
Безусловные рефлексы, осуществляющие врожденные реакции. В подкорковых отделах сосредоточены нервные аппараты, в которые непосредственно поступают нервные импульсы от внутренней среды, отражающие потребности организма. Деятельность подкорковых образований является физиологическим фундаментом для деятельности коры, направляя ее на внешнюю среду. Безусловные рефлексы после выработки на них условных рефлексов (раздражителей) обычно в чистом виде не осуществляются. В естественных условиях безусловный раздражитель действует всегда в комплексе с рядом агентов, становящихся его сигналами. Например, пища обладает различными случайными признаками, которые являются условными, или сигнальными, раздражителями, ее сигналами. Этими раздражителями могут быть разные признаки - вид, звук, которые и становятся возбудителями условного рефлекса, выражающегося в движении животного по направлению к пище.
Совпадение действия любого агента с действием безусловного раздражителя (пищи) делает этот агент сигналом (в данном случае пищи). Выработка условных рефлексов обеспечивает реагирование организма на огромную массу агентов, являющихся сигналами безусловного раздражителя: к выделению того, что нужно для существования организма, что надо взять из окружающей среды и чего надо беречься, бояться, от чего надо удаляться. Таким образом, безусловные рефлексы в норме спаяны воедино с условнорефлекторными, образуя сложнорефлекторные.
Аналитическая деятельность мозга неразрывно связана с синтетической деятельностью, благодаря которой выделенные анализом отдельные изменения среды объединяются в разнообразные комплексы, включающиеся во временные связи с той или иной деятельностью организма.
Нервный синтез - это замыкание между агентом, воздействующим на рецепторы, и ответной деятельностью. Синтез происходит и в низших подкорковых отделах центральной нервной системы, например при простых спинномозговых рефлексах, когда в ответ на раздражение рецептора происходит сокращение мышц (коленный рефлекс). Но здесь имеет место постоянная связь: раздражение данного рецептора вызывает одну и ту же реакцию. Другими словами, безусловный рефлекс характеризуется постоянным замыканием между агентом, воздействующим на рецептор, и ответной деятельностью. Он является проявлением постоянной связи раздражителя внешнего мира с функциями организма.
Высшей формой синтеза является корковый синтез, т. е. деятельность коры головного мозга. Она выражается в образовании временных связей или замыкательной деятельностью, которая и лежит в основе условного рефлекса. Условные рефлексы представляют лишь временные связи агента внешней среды с той или иной ответной деятельностью, так как имеют дело только с сигналами безусловных раздражителей, которые в зависимости от условий могут меняться. Деятельность коры мозга есть сигнальная с бесчисленным количеством сигналов и с переменной сигнализацией (запах, вид пищи).
Каждая рефлекторная деятельность происходит лишь в том случае, если к осуществляющим ее эффекторным органым передаются импульсы, возникающие в центральной нервной системе в ответ на раздражение рецепторов. Кора мозга не имеет непосредственной связи с периферическими органами (мышцами, железами). Эфферентные волокна, непосредственно иннервирующие различные органы тела, отходят от нервных клеток, расположенных в продолговатом и спинном мозгу, через которые происходят и безусловные рефлексы.
Значение различных областей коры мозга. Как указано выше, многочисленные корковые центры разбросаны по всей коре мозга и различные участки коры не одинаковы по своим функциям, а также по строению и расположению клеток, поэтому кору полушарий и разбивают обычно для удобства изучения на отдельные области, бродман разделил ее на 52 поля. В настоящее время существует деление коры более чем на 200 цитоархитектонических полей.
Двигательная (моторная) зона занимает переднюю центральную извилину обоих полушарий и соседние с ней участки лобной доли. Здесь известны более или менее обособленные центры для рук, ног, речевых мышц, мышц, участвующих в письме, и т. д. Отсюда центробежные волокна, получая перерыв в двигательных клетках стволовой части головного мозга и передних рогов спинного мозга, несут импульсы к поперечнополосатой мускулатуре тела.
Двигательная зона коры является местом восприятия, зоной корковой проекции рецепторов мышц, корковым концом двигательного анализатора. Так как раздражение этой зоны всегда происходит одновременно с движением тех мышц, откуда приходят импульсы, возникает временная связь (замыкание) чувствительного или афферентного импульса с двигательными невронами - пирамидными клетками Беца. Временная связь вследствие постоянства совпадения раздражения мышечных рецепторов с движением превращается в постоянную морфологическую. Пирамидные клетки являются путем от двигательного анализатора к двигательным клеткам спинного мозга.
Постоянное, в течение жизни тысяч поколений повторяющееся сочетание во время сокращения определенных мышц и действие импульсов от рецепторов этих мышц на область двигательного анализатора, привело к формированию эфферентных путей от корковой проекции мышечных рецепторов (двигательной зоны) к мотоневронам. Этим и объясняется, что корковый отдел двигательного анализатора является и эффекторной зоной.
Зона тактильной чувствительности кожи располагается позади роландовой борозды в задней центральной извилине. Сюда по чувствительным волокнам поступают возбуждения с периферии - от кожи. Здесь замыкаются рефлекторные дуги чувствительных раздражений со всей поверхности тела. Возбуждения чувствительных центров вызывают ощущения боли, холода, тепла и т. п.
Корковый отдел зрительного анализатора представляет собой проекцию сетчатки глаза в область шпорной борозды. Здесь заложены нервные центры, к которым поступают возбуждения от органов зрения, т. е. здесь замыкаются рефлекторные дуга, проводящие раздражения зрительных рецепторов сетчатки глаза. В этой области особо выделяется участок, известный под названием зрительного центрa pечи, или центра чтения (корковая зона речедвигательного анализатора).
Корковый отдел слухового анализатора занимает часть коры на наружной поверхности височной доли, главным образом в верхней височной извилине и в передней части поперечной извилины. Здесь замыкаются рефлекторные дуги раздражений, поступающих от органа слуха. При поражении некоторых участков этой области наступает музыкальная глухота - музыкальные звуки воспринимаются как беспорядочный шум. Здесь также часть слуховой области обосабливается как слуховой центр речи (Вернике), поражение ее вызывает словесную глухоту; при этом утрачивается способность распознавать слова и понимать речь, при сохранении восприятия слов как звуков наступает сенсорная афазия*.
* (От греческого А - не, phasis - речь.)
Корковые отделы анализаторов обоняния и вкуса располагаются на внутренней и нижней поверхности височной доли (извилина аммонова рога). Здесь замыкаются рефлекторные дуги раздражений, поступающих от вкусовых рецепторов языка и обонятельных рецепторов носа. Выключение обонятельной области вызывает расстройство обоняния или полную его потерю - аносмию. При этом необходимо отметить, что местонахождение "вкусового центра" в пределах коры больших полушарий полностью еще не уточнено.
Центр речи, или центр Брока, расположен в третьей лобной извилине левого полушария. Животные способны производить только нечленораздельные звуки (лай, ржание, рев и т. д.), членораздельная же речь свойственна только человеку. Центр речи в коре головного мозга животных отсутствует. У человека центр речи развивается сравнительно поздно, примерно на втором году жизни. Как известно, ребенок, подобно животному, способен только произносить звуки, кричать, но не говорить.
Необходимо еще заметить, что корковые зоны органов чувств (зрительная, слуховая, осязательная и другие отделы) занимают только небольшую часть коры полушарий мозга. Остальную же, самую большую, часть коры занимают центры более высокого порядка, которые выполняют наиболее сложную работу, - это так называемые объединяющие (ассоциативные) центры. В отличие от других центров, связанных центростремительными путями с органами чувств (ухом, глазом, кожей и др.) и через них с внешним миром, высшие центры не находятся в непосредственной связи с внешним миром, к ним притекают возбуждения из различных других центров коры, в них отдельные ощущения объединяются и перерабатываются в цельный образ, представление или идею. Например, мы держим в руке яркий по окраске и резкий по запаху цветок; благодаря осязательным, зрительным и обонятельным ощущениям мы получаем об этом цветке определенное представление. Отдельные ощущения о цветке, как мы уже знаем, возникают в различных областях мозговой коры, где они как бы откладываются, запечатлеваются и запоминаются. Но при мысли об этом цветке у нас возникают не разрозненные отдельные, ранее полученные ощущения, а полный образ, весь цветок в целом с его формой, окраской и даже запахом. Вот эта цельность образа и является продуктом ассоциативной деятельности корковых зон, объединяющих и связывающих в одном стройном сочетании работу многих других центров.
К таким сложным центрам в коре относится и центр речи. Когда ребенка учат говорить и произносят какие-нибудь слона, например "мяч", звуковые раздражения по слуховому нерву проводятся в корковый отдел слухового анализатора, где вырабатываются звуковые ощущения и где они запечатлеваются. Но слово - не сочетание пустых звуков, оно имеет содержание. Со словом "мяч" для осмысленного его усвоения у ребенка должны связываться известные ощущения; таким образом, должна быть установлена связь между звуковыми образами ирительными осязательными и другими ощущениями, доставляемыми мячом. Эта работа выполняется так называемым сенсорным центром речи в левой первой височной извилине. А для того чтобы произнести данное слово, должен прийти в движение ряд мышц, различным образом напрягающихся. Каждая мышца сокращается, как известно, под влиянием возбуждения, идущего от двигательных отделов коры. Согласованное же гармоническое участие всех мышц, сокращение которых необходимо для произнесения слова, возможно только тогда, когда все двигательные центры находятся под общим руководством. Эту роль высшего регулятора берет на себя участок коры в одной из лобных извилин левого полушария, носящий название двигательного центра речи, который обеспечивает согласованное сокращение мышц речи.
При поражении двигательного цетра речи, несмотря на целость мышц речи и нервных путей, ведущих к ним, произношение слов невозможно, но больной при этом понимает все, что говорят. Если же при целости двигательного центра поражен каким-либо болезненным процессом сенсорный центр речи, т. е. то место, где устанавливается и запечатлевается связь между словом и его содержанием, больной слышит речь, но не понимает ее, он в состоянии сам произносить слова, но они для него являются пустыми звуками, утратившими всякий смысл, скрытый в каждом слове. Речь есть одна из наиболее сложных функций человека. Функция речи является филогенетически наиболее новой, она развивается поздно и зависит от индивидуального развития (онтогенеза) человека, составляя специфическую особенность его корковой деятельности, отсутствующую у животных. Речь человека вырабатывается, как мы видели, в результате сочетания функций многих центров коры головного мозга и поэтому требует для своего осуществления деятельности почти всей коры в целом. Тем не менее на основании клинических данных имеется возможность все же выделить некоторые определенные участки коры больших полушарий, функционирование которых имеет прямое отношение к осуществлению речи.
Белое вещество мозга. На разрезах мозгового вещества полушарий обнаруживается лежащее внутри белое вещество с заключенными в нем серыми ядрами. Белое вещество полушарий состоит из огромного количества нервных волокон, соединяющих, как уже упоминалось, различные участки или области одного и того же полушария (ассоциативные волокна), или же через волокна мозолистого тела полушария между собою (комиссуральные). Кроме того, в состав белого вещества входят волокна, соединяющие кору с нижележащими частями мозга в центробежном и центростремительном направлениях (проекционные волокна); эти волокна и представляют собой продолжение ножек мозга, связывающих кору полушарий мозга с периферией.
Желудочки мозга (рис. 289). Белое вещество не занимает всего пространства внутри мозговых полушарий. Внутри белого вещества в каждом полушарии располагается по одному боковому желудочку мозга, которые представляют собой остатки полостей первого мозгового пузыря; каждый из этих желудочков соединяется межжелудочковым (монроевым) отверстием с полостью III желудочка, представляющую собой узкую вертикальную щель. Боковой желудочек имеет очень сложные очертания и состоит из трех рогов - переднего, направляющегося в лобную долю, нижнего - в височную, и заднего, уходящего в затылочную долю.
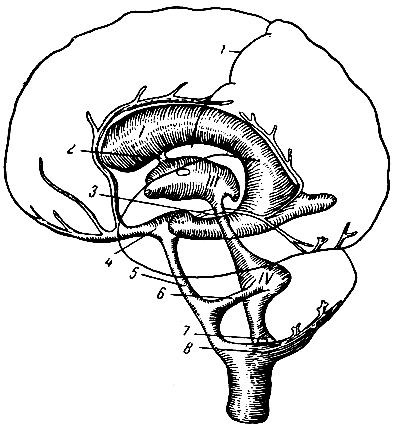
Рис. 289. Схематическое изображение мозговых желудочков и их связей с субарахноидальным пространством. 1 - боковой желудочек с передним, задним и нижним рогом; III - третий желудочек; IV - четвертый желудочек; 1 - роландова борозда; 2 - монроево отверстие; 3 - сильвиев водопровод; 4 - cisterna interpeduncularis; 5 - cisterna pontis; 6 - отверстие Люшка; 7 - отверстие Maжанди; 8 - cisterna magna
Под мозжечком находится IV желудочек мозга. Полость его имеет вид шатра, основанием (дном) которого служит ромбовидная ямка, а крышей - две тонкие пластинки (передний и задний парус). Задняя часть крыши истончена. Через среднее непарное отверстие (foramen Magendie) и два боковых (foramen Luschka) осуществляется связь подпаутинного пространства с IV желудочком. Внизу IV желудочек переходит в центральный канал спинного мозга, а впереди и вверху - в мозговой водопровод.
В полостях боковых желудочков, так же как и в полостях III и IV желудочков, вворачивается мягкая мозговая оболочка с большим количеством кровеносных сосудов, образуя в них сосудистые сплетения (plexus chorioideus), из которых в полость желудочков проникает цереброспинальная жидкость; эпендиме, выстилающей стенки мозговых желудочков, видимо, тоже присуща способность выделять цереброспинальную жидкость.
Все желудочки головного мозга и центральный канал спинного мозга составляют одну общую полость - остаточную полость бывшей мозговой трубки.
Проводящие пути (рис. 290 и 291). Рассмотрим те сановные нервные пути, которые связывают кору полушарий со всеми воспринимающими аппаратами и со всеми рабочими органами тела.
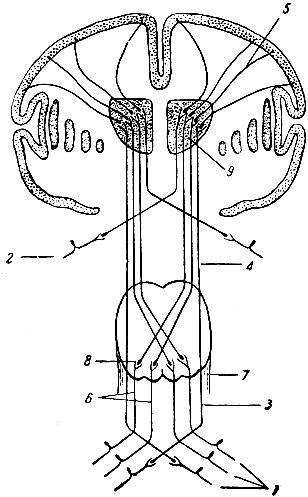
Рис. 290. Схема чувствительных путей. 1 - задние корешки спинномозговых нервов; 2 - черепномозговой нерв; 3, 4 и 5 - пути проведения осязательных, болевых и температурных ощущений; 6 - пути проприоцептивной (мышечно-суставной и кожной) чувствительности; 7 - продолговатый мозг; 8 - ядра нежного и клиновидного пучка; 9 - зрительный бугор
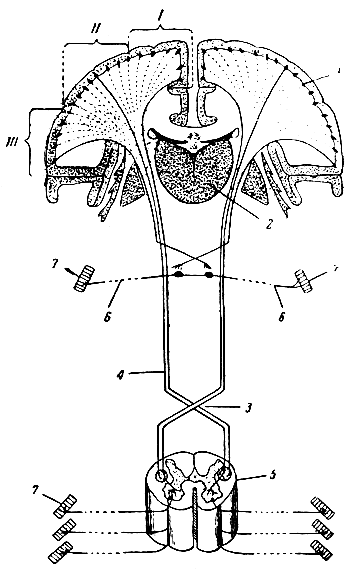
Рис. 291. Схема двигательных путей. 1 - мозговая кора; 2 - зрительный бугор; 3 - перекрест пирамид; 4 - пирамидный путь; 5 - отрезок спинного мозга с передними корешками нервов; 6 - двигательный головной нерв; 7 - мышцы; I, II, III - двигательная область мозговой коры (глубокие слои коры)
Периферические чувствительные нейроны с их нервными окончаниями, т. е. клетки спинномозговых узлов, являются началом чувствительного пути. Как мы уже знаем, из центральных нейритов этих клеток составляются задние спинномозговые корешки. Длинные веточки нервных волокон этих корешков, войдя в спинной мозг, поднимаются вверх в задних канатиках белого вещества, где образуют два пучка - нежный (Голля) и клиновидный (Бурдаха), которые, не перекрещиваясь, проходят до продолговатого мозга, где в них заложены чувствительные ядра, в которых оканчивается первый нейрон пути. Второй нейрон пути составляют клетки, залегающие в этих ядрах; их нейриты направляются вверх и образуют медиальную петлю, волокна которой после перекреста идут выше через ножки мозга и оканчиваются в ядрах зрительного бугра.
Клетки зрительного бугра составляют третий нейрон пути; их нейриты входят во внутреннюю капсулу и оканчиваются в теменной доле полушарий позади центральной борозды, где чувствительные (сенсорные) центры коры принимают возбуждение, идущее с периферии и воспринимаемое как то или иное ощущение.
Другая часть нервных волокон задних корешков входит в серое вещество задних рогов спинного мозга; здесь у клеток задних рогов оканчивается первый нейрон. Второй нейрон пути представляют клетки задних рогов, нейриты которых, перейдя на другую сторону, идут вверх в боковых канатиках белого вещества; в продолговатом мозгу эти волокна присоединяются к волокнам медиальной петли после ее перекреста и оканчиваются в зрительном бугре. Этот пучок называется спинально-бугровым. Третий нейрон пути составляют клетки зрительного бугра, нейриты которых идут в составе внутренней капсулы вместе с медиальной петлей и оканчиваются также в коре теменной доли полушарий.
Нежный и клиновидный пучки являются проводниками мышечного чувства (глубокой чувствительности) и отчасти осязания, спинально-бугровый пучок - проводником болевого и температурного чувства (поверхностной чувствительности), а также и осязательного. К волокнам этих пучков на протяжении ствола мозга присоединяются чувствительные волокна черепномозговых нервов, которые оканчиваются в различных участках коры.
Итак, чувствительный путь (путь ощущений) состоит из трех (и более) нейронов и служит проводником поверхностной и глубокой чувствительности (кожно-мышечного чувства), тактильной, болевой и температурной чувствительности. Но, кроме того, существуют и более сложные пути - с заходом в мозжечок и четверохолмие. Эти пути образуют в боковом канатике спинного мозга два мозжечковых пучка: прямой пучок Флексига и перекрещеный пучок Говерса.
Клетки мозговой коры являются конечными пунктами этого длинного центростремительного (чувствительного) пути, о котором сейчас шла речь. Другие клетки коры являются начальной станцией, исходным пунктом другого нервного пути - центробежного, т. е. пути произвольных движений.
Двигательный (центробежный) путь начинается от клеток мозговой коры в области передней центральной извилины (обоих полушарий) и ближайших участков лобной доли. Возбуждение, возникшее в двигательных центрах коры в результате импульсов от других нервных центров, распространяется по нейритам, направляющимся в составе внутренней капсулы IB мозговой ствол. Пройдя ножки мозга и мозговой мост, волокна этого пути образуют пирамидные пучки продолговатого мозга; большая часть волокон переходит здесь на другую сторону, образуя перекрест пирамид. Перекрещенные волокна идут дальше в боковых канатиках белого вещества спинного мозга, составляя боковой пирамидный путь, а неперекрещенные образуют прямой пирамидный пучок в передних канатиках спинного мозга и переходят на другую сторону только в конце своего пути. Волокна обоих пирамидных лучков оканчиваются в передних рогах спинного мозга. Здесь возбуждение переключается на двигательный нейрон передних рогов и дальше по его нейриту через передний корешок, а затем через смешанный спинномозговой нерв достигает мышцы.
В стволовой части мозга некоторая часть пирамидных волокон оканчивается в двигательных ядрах среднего и продолговатого мозга; клетки этих ядер составляют второй нейрон для черепномозговых нервов.
Кроме этого главного двигательного пути, имеются и более сложные пути через ядра среднего мозга и мозжечка.
На подопытных животных и путем клинических наблюдений над больными людьми, а также путем изучения изменений различных частей мозга и проводящих путей, вследствие болезней были установлены перекресты проводящих путей. Теперь мы уже точно знаем, что если парализована ("отнялась") правая рука или нога, то причину нужно искать в нарушении нормальной работы двигательных центров коры или целости центробежных двигательных волокон с левой стороны.
Всякое раздражение с периферического воспринимающего аппарата (в коже, слизистой оболочке, сухожилиях и др.) передается на рабочие органы, пробегая при этом по различным рефлекторным дугам. Когда раздражение передается с центростремительного волокна на центробежное через спинной мозг на уровне входа задних корешков, - это будет самая простая и короткая дуга, наиболее элементарная из всех мыслимых рефлекторных дуг высших животных. Только для очень немногих рефлексов допускается возможность ограничения в пределах одного сегмента спинного мозга - это сухожильные рефлексы, например коленный рефлекс. Вообще же подавляющее большинство рефлексов высших позвоночных животных не ограничивается одним сегментом; они не ограничиваются и одним спинным мозгом, так как в деятельность вовлекаются расположенные выше отделы центральной нервной системы. Таким образом, возбуждение, пришедшее в спинной мозг с любого рецептора по любому афферентному (центростремительному) волокну, имеет возможность по сложной системе коротких и длинных проводящих путей спинного мозга перейти на любой эфферентный (центробежный) нейрон и достигнуть любого эффекторного органа*.
* (Эффекторным органом называют образование, выполняющее внешнюю работу под влиянием пришедшего к нему возбуждения, например мышца, железа.)
Итак, по восходящей и нисходящей ветвям центростремительного волокна через промежуточный нейрон раздражение может попасть на двигательные нейроны различных этажей спинного мозга. По длинным восходящим ветвям нежного и клиновидного пучков оно может проникнуть в стволовую часть, например в продолговатый мозг, где через промежуточные нейроны передается на двигательный нейрон, - это будет более сложная рефлекторная дуга. Еще более сложной рефлекторная дуга будет в случае, когда раздражение передается по ветвям спинномозговых путей в мозжечок, а оттуда через ряд нейронов в ствол мозга и уже из ствола на двигательный нейрон спинного мозга.
И, наконец, примером наиболее сложного рефлекторного пути является случай, когда через зрительные бугры раздражение попадает в кору голодного мозга и через двигательные нейроны коры направляется в спинной мозг.
Значение больших полушарий головного мозга для жизни животных и человека. С деятельностью мозговых полушарий тесно связана вся умственная (интеллектуальная) жизнь человека. В коре полушарий создаются разнообразные и многочисленные нервные связи, позволяющие организму приспособлять свое поведение к различным и быстро меняющимся условиям жизни. С совершающимися в нервных клетках коры процессами связаны все наши сознательные ощущения и образы. Кора полушарий - это вместилище всей нашей интеллектуальной жизни, это мастерская наших желаний, мыслей и чувств. И. П. Павлов впервые доказал возможность изучать все сложные психические процессы так же научно, объективно, как и все другие физиологические функции организма. Он очень ярко и убедительно доказал, что все то, что раньше называли душой, есть продукт жизнедеятельности коры мозга, вполне доступный объективному анализу.
Кора больших полушарий мозга имеет связи и со всеми без исключения внутренними органами.
Корковые образования, воспринимающие импульсы от внутренних органов (сердца, сосудов), вероятно, также являются областями коры, от которых исходят эфферентные влияния на эти органы. Ряд областей коры при электрическом раздражении вызывают эффекты со стороны зрачков, кишечника, органов дыхания и т. д.
В коре головного мозга имеется, так сказать, представительство всех функций - высших и низших, анимальных и вегетативных. И. П. Павлов писал, что в коре головного мозга наряду "с грандиозным представительством внешнего мира имеется также и широкое представительство внутреннего мира организма, т. е. состояний, работы массы органов и тканей, массы внутренних органических процессов". "Наши исследования, основываясь на учении и методе условного рефлекса, с полной достоверностью показали, что внутренняя для организма среда непрерывно посылает сигналы в центральную нервную систему, в кору головного мозга, создавая специализированную информацию о событиях, совершающихся во внутренних органах. Доказано, что эти сигналы рефлекторно влияют на функциональное состояние головного мозга, отражая в его деятельности все многообразие работы внутренних органов. Важным обстоятельством при этом следует считать точно установленные факты, что раздражения внутренних органов способны превратиться в условный рефлекс, а по природе эти внутренние (интерорецептивные) условные рефлексы принципиально тождественны с теми, которые открыл Павлов в отношении внешних для организма раздражений" (К. М. Быков).
Влияние головного мозга на внутренние органы, на обмен веществ освещает весьма важный вопрос о связи болезней с функцией организма как целого.
При использовании материалов сайта активная ссылка обязательна:
http://anfiz.ru/ 'Анатомия и физиология человека'