Нервная ткань
Нервная ткань является основным компонентом нервной системы. Она состоит из нервных клеток и клеток нейроглии. Нервные клетки способны под действием раздражения приходить в состояние возбуждения, вырабатывать импульсы и передавать их. Эти свойства определяют специфическую функцию нервной системы. Нейроглия органически связана с нервными клетками и осуществляет трофическую, секреторную, защитную функции и функцию опоры.
Нервные клетки - нейроны, или нейроциты, представляют собой отростчатые клетки. Размеры тела нейрона колеблются в значительных пределах (от 3 - 4 до 130 мкм). По форме нервные клетки также очень разные (рис. 10). Отростки нервных клеток проводят нервный импульс из одной части тела человека в другую, длина отростков от нескольких микрон до 1,0 - 1,5 м.
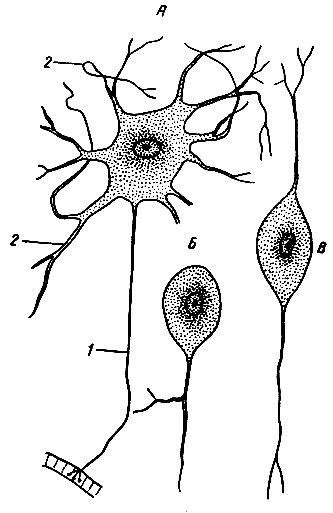
Рис. 10. Нейроны (нервные клетки). А - мультиполярный нейрон; Б - псевдоуниполярный нейрон; В - биполярный нейрон; 1 - аксон; 2 - дендрит
Различают два вида отростков нервной клетки. Отростки первого вида проводят импульсы от тела нервной клетки к другим клеткам или тканям рабочих органов, они называются нейритами, или аксонами. Нервная клетка имеет всегда только один аксон, который заканчивается концевым аппаратом на другом нейроне или в мышце, железе. Отростки второго вида называются дендритами, они древовидно ветвятся. Их количество у разных нейронов различно. Эти отростки проводят нервные импульсы к телу нервной клетки. Дендриты чувствительных нейронов имеют на периферическом конце специальные воспринимающие аппараты - чувствительные нервные окончания, или рецепторы.
По количеству отростков нейроны делятся на биполярные (двухполюсные) - с двумя отростками, мультиполярные (многополюсные) - с несколькими отростками. Особо выделяют псевдоуниполярные (ложные однополюсные) нейроны, нейрит и дендрит которых начинаются от общего выроста тела клетки с последующим Т-образным делением. Такая форма характерна для чувствительных нейроцитов.
Нервная клетка имеет одно ядро, содержащее 2 - 3 ядрышка. Цитоплазма нейронов, помимо органелл, характерных для любых клеток, содержит хроматофильное вещество (вещество Ниссля) и нейрофибриллярный аппарат. Хроматофильное вещество представляет собой зернистость, образующую в теле клетки и дендритах нерезко ограниченны глыбки, окрашивающиеся основными красителями. Оно меняется в зависимости от функционального состояния клетки. В условиях перенапряжения, травмы (перерезка отростков, отравление, кислородное голодание и др.) глыбки распадаются и исчезают. Этот процесс получил название хроматолиза, т. е. растворения.
Другим характерным компонентом цитоплазмы нервных клеток являются тонкие нити - нейрофибриллы. В отростках они лежат вдоль волокон параллельно друг другу, в теле клетки образуют сеть.
Нейроглия представлена клетками различной формы и величины, которые делятся на две группы: макроглию (глиоциты) и микроглию (глиальные макрофаги) (рис. 11). Среди глиоцитов различают эпендимоциты, астроциты и олигодендроциты. Эпендимоциты выстилают спинномозговой канал и желудочки головного мозга. Астроциты образуют опорный аппарат центральной нервной системы. Олигодендроциты окружают тела нейронов в центральной и периферической нервной системе, образуют оболочки нервных волокон и входят в состав нервных окончаний. Клетки микроглии подвижны и способны фагоцитировать.
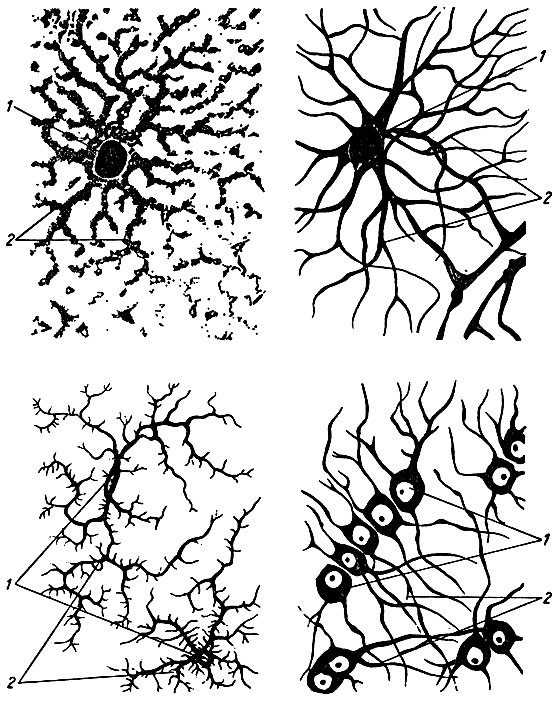
Рис. 11. Клетки нейроглии. 1 - тело; 2 - отростки
Нервными волокнами называются отростки нервных клеток (осевые цилиндры), покрытые оболочками. Оболочка нервных волокон (нейролемма) образована клетками, которые называются нейролеммоцитами (шванновские клетки). В зависимости от строения оболочки различают безмиелиновые (безмякотные) и миелиновые (мякотные) нервные волокна. Безмиелиновые нервные волокна характеризуются тем, что леммоциты в них лежат плотно друг к другу и образуют тяжи протоплазмы. В такой оболочке располагаются один или несколько осевых цилиндров. Миелиновые нервные волокна имеют более толстую оболочку, внутренняя часть которой содержит миелин. При обработке осмиевой кислотой гистологических препаратов миелиновая оболочка окрашивается в темно-коричневый цвет. На определенном расстоянии в миелиновом волокне расположены косые белые линии - насечки миелина и сужения - узлы нервного волокна (перехваты Ранвье). Они соответствуют границам леммоцитов. Миелиновые волокна толще безмиелиновых, их диаметр 1 - 20 мкм.
Пучки миелиновых и безмиелиновых нервных волокон, покрытые соединительнотканной оболочкой, образуют нервные стволы, или нервы. Соединительнотканная оболочка нерва называется эпиневрием. Она проникает в толщу нерва и покрывает пучки нервных волокон (периневрий) и отдельные волокна (эндоневрий). В эпиневрии располагаются кровеносные и лимфатические сосуды, которые проходят в периневрий и эндоневрий.
Перерезка нервных волокон вызывает дегенерацию периферического отростка нервного волокна, при которой он распадается на участку различной величины. На месте перерезки возникает воспалительная реакция и образуется рубец, через который в дальнейшем возможно прорастание центральных отрезков нервных волокон при регенерации (восстановлении) нерва. Регенерация нервного волокна начинается с интенсивного размножения леммоцитов и образования из них своеобразных лент, проникающих в рубцовую ткань. Осевые цилиндры центральных отростков образуют на концах утолщения - колбы роста и врастают в рубцовую ткань и ленты леммоцитов. Периферический нерв растет со скоростью 1 - 4 мм/су т.
Нервные волокна заканчиваются концевыми аппаратами - нервными окончаниями (рис. 12). По функции различают три группы нервных окончаний: чувствительные, или рецепторы, двигательные и секреторные, или эффекторы, и окончания на других нейронах - межнейрональные синапсы.
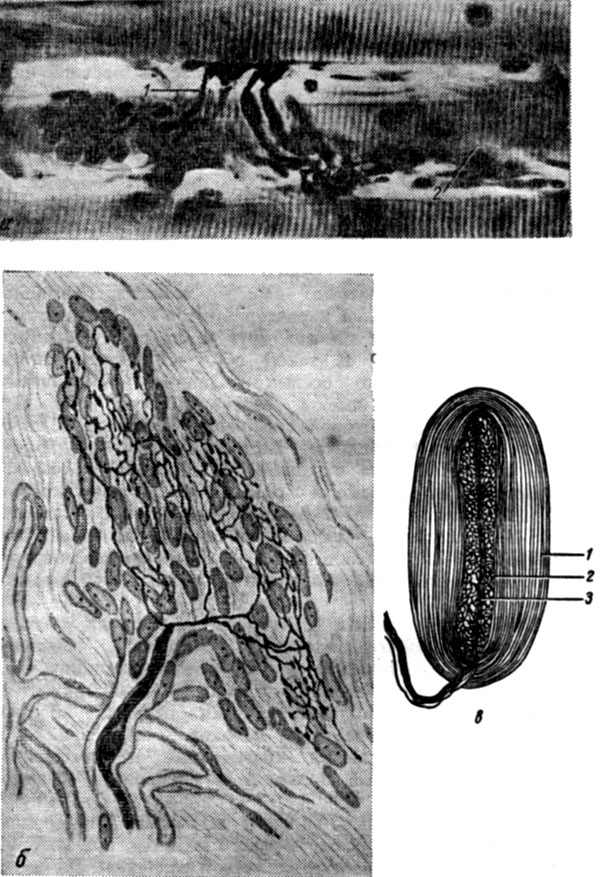
Рис. 12. Нервные окончания. а - нервно-мышечное окончание: 1 - нервное волокно; 2 - мышечное волокно; б - свободное нервное окончание в соединительной ткани; в - пластинчатое тельце (тельце Фатера - Пачини): 1 - наружная колба (луковица); 2 - внутренняя колба (луковица); 3 - концевой отдел нервного волокна
Чувствительные нервные окончания (рецепторы) образованы концевыми разветвлениями дендритов чувствительных нейронов. Они воспринимают раздражения из внешней среды (экстерорецепторы) и от внутренних органов (интерорецепторы). Различают свободные нервные окончания, состоящие только из концевого ветвления отростка нервной клетки, и несвободные, если в образовании нервного окончания принимают участие элементы нейроглии. Несвободные нервные окончания могут быть покрыты соединительнотканной капсулой. Такие окончания называются капсулированными: например, пластинчатого тельца (тельца Фатера - Пачини). Рецепторы скелетных мышц называются нервно-мышечными веретенами. Они состоят из нервных волокон, ветвящихся на поверхности мышечного волокна в виде спирали.
Эффекторы бывают двух типов - двигательные и секреторные. Двигательные (моторные) нервные окончания являются концевыми разветвлениями нейритов двигательных клеток в мышечной ткани и называются нервно-мышечными окончаниями. Секреторные окончания в железах образуют нервно-железистые окончания. Названные виды нервных окончаний представляют собой нервно-тканевой синапс.
Связь между нервными клетками осуществляется при помощи синапсов. Они образованы концевыми ветвлениями нейрита одной клетки на теле, дендритах или аксонах другой. В синапсе нервный импульс проходит только в одном направлении (с нейрита на тело или дендриты другой клетки). В различных отделах нервной системы они устроены по-разному.
Общая физиология возбудимых тканей
Все живые организмы и любая их клетка обладают раздражимостью, т. е. способностью отвечать на внешнее раздражение изменением обмена веществ.
Наряду с раздражимостью три вида ткани: нервная, мышечная и железистая - обладают возбудимостью. В ответ на раздражение в возбудимых тканях возникает процесс возбуждения.
Возбуждение представляет собой сложную биологическую реакцию. Обязательными признаками возбуждения являются изменение мембранного потенциала, усиление обмена веществ (повышение потребления О2, выделение СО2 и тепла) и возникновение деятельности, присущей данной ткани: мышца сокращается, железа выделяет секрет, нервная клетка генерирует электрические импульсы. В момент возбуждения ткань из состояния физиологического покоя переходит к присущей ей деятельности.
Следовательно, возбудимостью называют способность ткани отвечать на раздражение возбуждением. Возбудимость - это свойство ткани, тогда как возбуждение - это процесс, ответная реакция на раздражение.
Важнейшим признаком распространяющегося возбуждения является возникновение нервного импульса, или потенциала действия, благодаря которому возбуждение не остается на месте, а проводится по возбудимым тканям. Раздражителем, вызывающим возбуждение, может быть любой агент внешней или внутренней среды (электрический, химический, механический, термический и др.) при условии, что он является достаточно сильным, действует достаточно долго и нарастание его силы происходит достаточно быстро.
Биоэлектрические явления
Биоэлектрические явления - "животное электричество" было открыто в 1791 г. итальянским ученым Гальвани. Данные современной мембранной теории происхождения биоэлектрических явлений получены Ходжкиным, Кацом и Хаксли в исследованиях, проведенных с гигантским нервным волокном кальмара (диаметром 1 мм) в 1952 г.
Плазматическая мембрана клетки (плазмолемма), ограничивающая снаружи цитоплазму клетки, имеет
толщину около 10 нм и состоит из двойного слоя липидов, в который погружены глобулы белков (молекулы, свернутые в клубки или спирали). Белки выполняют функции ферментов, рецепторов, транспортных систем, ионных каналов. Они либо частично, либо целиком погружены в липидный слой мембраны (рис. 13). В состав мембраны входит также небольшое количество углеводов.
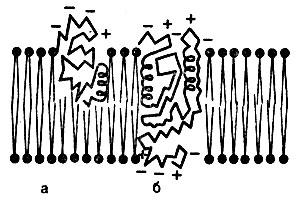
Рис. 13. Модель клеточной мембраны как жидкой мозаики из липидов и белков - поперечный разрез (Стерки П., 1984). а - липиды; в - белки
Сквозь мембрану движутся различные вещества в клетку и из клетки. Регуляция этого процесса - одна из основных функций мембраны. Основными ее свойствами являются избирательная и изменчивая проницаемость. Для одних веществ она служит барьером, для других - входными воротами. Вещества могут проходить через мембрану по закону концентрационного градиента (диффузия от большей концентрации к меньшей), по электрохимическому градиенту (разная концентрация заряженных ионов), путем активного транспорта - работа натрий-калиевых насосов.
Мембранный потенциал, или потенциал покоя. Между наружной поверхностью клетки и ее цитоплазмой существует разность потенциалов порядка 60 - 90 мВ (милливольт) , называемая мембранным потенциалом, или потенциалом покоя. Его можно обнаружить при помощи микроэлектродной методики. Микроэлектрод представляет собой тончайший стеклянный капилляр с диаметром кончика 0,2 - 0,5 мкм. Его заполняют раствором электролита (КС1). Второй электрод обычных размеров погружают в раствор Рингера, в котором находится исследуемый объект. Через усилитель биопотенциалов электроды подводят к осциллографу. Если под микроскопом с помощью микроманипулятора микроэлектрод ввести внутрь нервной клетки, нервного или мышечного волокна, то в момент прокола осциллограф покажет разность потенциалов - потенциал покоя (рис. 14). Микроэлектрод настолько тонок, что он практически не повреждает мембраны.
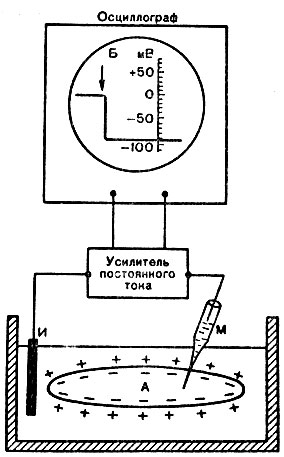
Рис. 14. Измерение потенциала покоя мышечного волокна (А) при помощи внутриклеточного микроэлектрода (схема). М - микроэлектрод; И - индифферентный электрод. Луч на экране осциллографа показан стрелкой
Мембрайно-ионная теория объясняет происхождение потенциала покоя неодинаковой концентрацией несущих электрические заряды К+, Na+ и Сl- внутри и вне клетки и различной проницаемостью для них мембраны.
В клетке в 30 - 50 раз больше К+ и в 8 - 10 раз меньше Na+, чем в тканевой жидкости. Следовательно, внутри клетки преобладают К+, снаружи - Na+. Основным анионом тканевой жидкости является Сl-. В клетке преобладают крупные органические анионы, которые не могут диффундировать сквозь мембрану. (Как известно, катионы имеют положительный заряд, а анионы - отрицательный.) Состояние неодинаковой ионной концентрации по обе стороны плазматической мембраны называют ионной асимметрией. Она поддерживается работой натрий-калиевых насосов, которые непрерывно перекачивают Na+ из клетки и К+ в клетку. Работа эта осуществляется с затратой энергии, освобождающейся при расщеплении аденозинтрифосфорной кислоты. Ионная асимметрия - физиологическое явление, сохраняющееся пока клетка жива.
В покое проницаемость мембраны значительно выше для К+, чем для Na+. В силу высокой концентрации ионы К+ стремятся выйти из клетки наружу. Сквозь мембрану они проникают на наружную поверхность клетки, но дальше уйти не могут. Крупные анионы клетки, для которых мембрана непроницаема, не могут последовать за калием, и скапливаются на внутренней поверхности мембраны, создавая здесь отрицательный заряд, который удерживает электростатической связью проскочившие через мембрану положительно заряженные ионы калия. Таким образом возникает поляризация мембраны, потенциал покоя; по обе ее стороны образуется двойной электрический слой: снаружи из положительно заряженных ионов К+ , а внутри из отрицательно заряженных различных крупных анионов.
Потенциал действия. Потенциал покоя сохраняется до тех пор, пока не возникло возбуждение. Под действием раздражителя проницаемость мембраны для Na+ повышается. Концентрация Na+ снаружи клетки в 10 раз больше, чем внутри нее. Поэтому Na+ сначала медленно, а затем лавинообразно устремляются внутрь. Ионы натрия заряжены положительно, поэтому происходит перезарядка мембраны и ее внутренняя поверхность приобретает положительный заряд, а наружная - отрицательный. Таким образом происходит реверсия потенциала, изменение его на обратный знак. Он становится отрицательным снаружи и положительным внутри клетки. Этим объясняется давно известный факт, что возбужденный участок становится электроотрицательным по отношению к находящемуся в покое. Однако повышение проницаемости мембраны для Na+ длится недолго; она быстро снижается и повышается для К+. Это вызывает усиление потока положительно заряженных ионов из клетки во внешний раствор. В итоге происходит реполяризация мембраны, ее наружная поверхность приобретает снова положительный заряд, а внутренняя - отрицательный.
Электрические изменения мембраны в процессе возбуждения получили название потенциала действия. Длительность его измеряется тысячными долями секунды (миллисекундами), амплитуда равна 90 - 120 мВ.
Во время возбуждения Na+ входят в клетку, а К+ выходят наружу. Казалось бы, что концентрация ионов в клетке должна меняться. Как показали опыты, даже многочасовое раздражение нерва и возникновение в нем десятков тысяч импульсов не изменяют содержания в нем Na+ и К+. Это объясняется работой натрий-калиевого насоса, который после каждого цикла возбуждения разводит ионы по местам: накачивает К+ обратно в клетку и выводит из нее Na+. Насос работает на энергии внутриклеточного обмена веществ. Это доказывается тем, что яды, прекращающие обмен веществ, прекращают работу насоса.
Потенциал действия, возникая в возбужденном участке, становится раздражителем для соседнего невозбужденного участка мышечного или нервного волокна и обеспечивает проведение возбуждения вдоль мышцы или нерва.
Возбудимость различных тканей неодинакова. Наиболее высокой возбудимостью отличаются рецепторы, специализированные структуры, приспособленные к улавливанию изменений во внешней среде и внутренней среде организма. Затем следует нервная, мышечная и железистая ткани.
Мерой возбудимости является порог раздражения, т. е. та наименьшая сила раздражителя, которая способна вызвать возбуждение. Порог раздражения иначе называют реобазой. Чем выше возбудимость ткани, тем меньшей силы раздражитель способен вызвать возбуждение.
Кроме того, возбудимость можно характеризовать тем временем, в течение которого должен действовать раздражитель, чтобы вызвать возбуждение, иначе говоря, порогом времени. Наименьшее время, в течение которого должен действовать электрический ток пороговой силы, чтобы вызвать возбуждение, называется полезным временем. Полезное время характеризует скорость течения процесса возбуждения.
Возбудимость тканей увеличивается в процессе умеренной деятельности и снижается при утомлении. Возбудимость претерпевает фазовые изменения во время возбуждения. Как только в возбудимой ткани возникает процесс возбуждения, она утрачивает способность отвечать на новое, даже сильное раздражение. Это состояние называется абсолютной невозбудимостью, или абсолютной рефрактерной фазой. Через некоторое время возбудимость начинает восстанавливаться. На пороговое раздражение ткань еще не отвечает, но на сильное раздражение отвечает возбуждением, хотя амплитуда возникающего потенциала действия в это время значительно снижена, т. е. процесс возбуждения слаб. Это фаза относительной рефрактерности. После нее возникает фаза повышенной возбудимости или супернормальности. В это время можно вызвать возбуждение очень слабым раздражителем, ниже пороговой силы. Только после этого возбудимость приходит в норму.
Для исследования состояния возбудимости мышечной или нервной ткани наносят два раздражения друг за другом через определенные интервалы. Первое вызывает возбуждение, а второе - тестирующее - испытывает возбудимость. Если на второе раздражение реакции нет, значит, ткань невозбудима; реакция слабая - возбудимость понижена; реакция усилена - возбудимость повышена. Так, если на сердце наносить раздражение во время систолы, то возбуждения не последует, к концу диастолы раздражение вызывает внеочередное сокращение - экстрасистолу, что свидетельствует о восстановлении возбудимости.
На рис. 15 сопоставлены во времени процесс возбуждения, выражением которого служит потенциал действия, и фазовые изменения возбудимости. Видно, что абсолютная рефрактерная фаза соответствует восходящей части пика - деполяризации, фаза относительной рефрактерности - нисходящей части пика - реполяризации мембраны и фаза повышенной возбудимости - отрицательному следовому потенциалу.
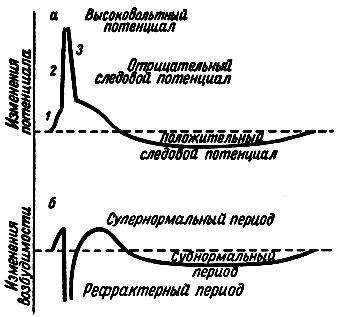
Рис. 15. Схемы изменений потенциала действия (а) и возбудимости нервного волокна (б) в различные фазы потенциала действия. 1 - местный процесс; 2 - фаза деполяризации; 3 - фаза реполяризации. Пунктиром на рисунке обозначены потенциал покоя и исходный уровень возбудимости
Проведение возбуждения по нерву
Нерву присущи два физиологических свойства - возбудимость и проводимость, т. е. способность на раздражение отвечать возбуждением и проводить его. Проведение возбуждения является единственной функцией нервов. От рецепторов они проводят возбуждение к центральной нервной системе, а от нее - к рабочим органам.
С физической точки зрения нерв очень плохой проводник. Его сопротивление в 100 млн. раз больше, чем у медной проволоки того же диаметра, однако нерв отлично выполняет свою функцию, проводя импульсы без затухания на большое расстояние.
Как осуществляется проведение нервного импульса?
Согласно мембранной теории, каждый возбужденный участок приобретает отрицательный заряд, а так как соседний невозбужденный участок имеет положительный заряд, то два участка оказываются противоположно заряженными. При создавшихся условиях между ними потечет электрический ток. Этот местный ток является раздражителем для покоящегося участка, он вызывает его возбуждение и изменяет заряд на отрицательный. Как только это произойдет, между вновь возбужденным и соседним покоящимся участками потечет электрический ток и все повторится.
Так распространяется возбуждение в тонких, безмиелиновых нервных волокнах. Там, где есть миелиновая оболочка, возбуждение может возникать только в узлах нервного волокна (перехватах Ранвье), т. е. в точках, где волокно оголено. Поэтому в миелиновых волокнах возбуждение распространяется скачками от одного перехвата к другому и движется гораздо быстрее, чем в тонких безмиелиновых волокнах (рис. 16).

Рис. 16. Проведение возбуждения в миелиновом нервном волокне. Стрелками показано направление тока, возникающего между возбужденным (А) и соседним покоящимся (Б) перехватами
Следовательно, в каждом участке волокна возбуждение генерируется заново и распространяется не электрический ток, а возбуждение. Этим объясняется способность нерва проводить импульс без затухания (без декремента). Нервный импульс остается постоянным по величине в начале и в конце своего пути и распространяется с постоянной скоростью. Кроме того, все импульсы, которые проходят по нерву, совершенно одинаковы по величине и не отражают качества раздражения. Меняться может только их частота, которая зависит от силы раздражителя.
Величина и длительность импульса возбуждения определяются свойствами нервного волокна, по которому оно распространяется.
Скорость проведения импульса зависит от диаметра волокна: чем оно толще, тем быстрее распространяется возбуждение. Наибольшей скоростью проведения (до 120 м/с) отличаются миелиновые двигательные и чувствительные волокна, управляющие функцией скелетных мышц, поддерживающих равновесие тела и выполняющие быстрые рефлекторные движения. Наиболее медленно (0,5 - 15 м/с) проводят импульсы безмиелиновые волокна, иннервирующие внутренние органы, и некоторые тонкие чувствительные волокна.
Законы проведения возбуждения по нерву
Доказательством того, что проведение по нерву - процесс физиологический, а не физический, служит опыт с перевязкой нерва. Если нерв туго перетянуть лигатурой, то проведение возбуждения прекращается - закон физиологической целостности.
Нервное волокно может проводить импульс в обе стороны. Если нерв раздражать посредине, то на двух его концах осциллограф зарегистрирует потенциалы действия - закон двустороннего проведения возбуждения.
В нервах импульсы проводятся по отдельным нервным волокнам изолированно. Поэтому один и тот же нерв в одном направлении проводит чувствительные (афферентные), а в другом - двигательные (эфферентные) импульсы - закон изолированного проведения.
При использовании материалов сайта активная ссылка обязательна:
http://anfiz.ru/ 'Анатомия и физиология человека'